Effect of ACTN3 Genotype on Sports Performance, Exercise-Induced Muscle Damage, and Injury Epidemiology

 ,
,  ,
,  , ,
, , 
 ,
,  and
and 
{kind=link}
Abstract
1. Introduction
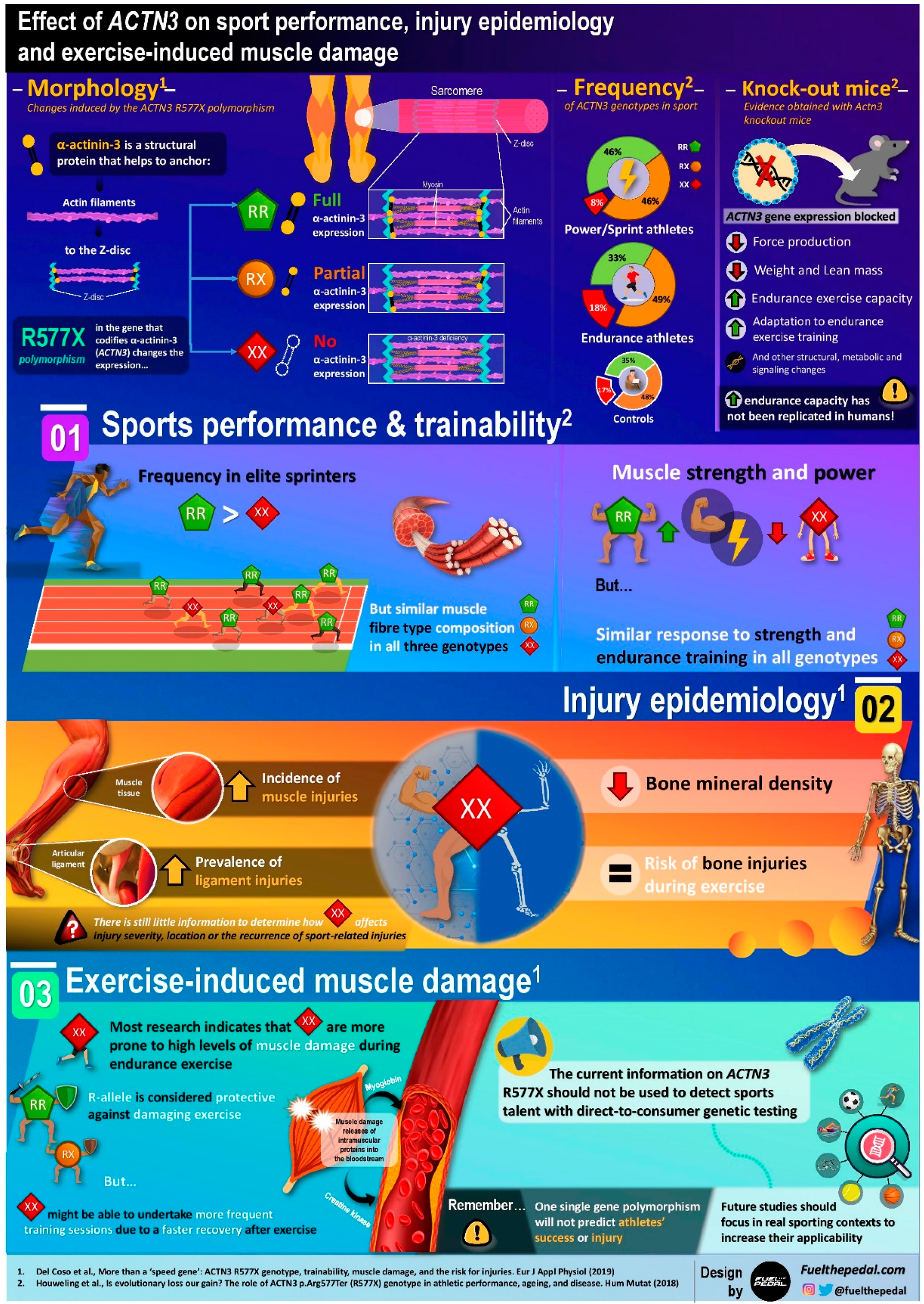
2. Effect of α-Actinin-3 Deficiency in Sports Performance
3. Mice with An Artificially-Induced α-Actinin-3 Deficiency
4. Effect of α-Actinin-3 Deficiency on Injury Epidemiology
5. Effect of α-Actinin-3 Deficiency on Exercise-Induced Muscle Damage
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Antero, J.; Saulière, G.; Marck, A.; Toussaint, J.F. A medal in the olympics runs in the family: A cohort study of performance heritability in the games history. Front. Physiol. 2018, 9, 1313. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, C.; Lesage, R.; Lortie, G.; Simoneau, J.A.; Hamel, P.; Boulay, M.R.; Pérusse, L.; Thériault, G.; Leblanc, C. Aerobic performance in brothers, dizygotic and monozygotic twins. Med. Sci. Sports Exerc. 1986, 18, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, C.; Warwick Daw, E.; Rice, T.; Pérusse, L.; Gagnon, J.; Province, M.A.; Leon, A.S.; Rao, D.C.; Skinner, J.S.; Wilmore, J.H. Familial resemblance for VO(2max) in the sedentary state: The HERITAHE family study. Med. Sci. Sports Exerc. 1998, 30, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Georgiades, E.; Klissouras, V.; Baulch, J.; Wang, G.; Pitsiladis, Y. Why nature prevails over nurture in the making of the elite athlete. BMC Genom. 2017, 18, 835. [Google Scholar] [CrossRef] [PubMed]
- Ahmetov, I.I.; Egorova, E.S.; Gabdrakhmanova, L.J.; Fedotovskaya, O.N. Genes and athletic performance: An update. Med. Sport Sci. 2016, 61, 41–54. [Google Scholar]
- Sarzynski, M.A.; Loos, R.J.F.; Lucia, A.; Pérusse, L.; Roth, S.M.; Wolfarth, B.; Rankinen, T.; Bouchard, C. Advances in exercise, fitness, and performance genomics in 2015. Med. Sci. Sports Exerc. 2016, 48, 1906–1916. [Google Scholar] [CrossRef]
- Jacob, Y.; Spiteri, T.; Hart, N.; Anderton, R. The potential role of genetic markers in talent identification and athlete assessment in elite sport. Sports 2018, 6, 88. [Google Scholar] [CrossRef]
- MacArthur, D.G.; North, K.N. A gene for speed? The evolution and function of α-actinin-3. BioEssays 2004, 26, 786–795. [Google Scholar] [CrossRef]
- Houweling, P.J.; Papadimitriou, I.D.; Seto, J.T.; Pérez, L.M.; Coso, J.D.; North, K.N.; Lucia, A.; Eynon, N. Is evolutionary loss our gain? The role of ACTN3 p.Arg577Ter (R577X) genotype in athletic performance, ageing, and disease. Hum. Mutat. 2018, 39, 1774–1787. [Google Scholar] [CrossRef]
- Del Coso, J.; Hiam, D.; Houweling, P.J.; Pérez, L.M.; Eynon, N.; Lucía, A. More than a ‘speed gene’: ACTN3 R577X genotype, trainability, muscle damage, and the risk for injuries. Eur. J. Appl. Physiol. 2019, 119, 49–60. [Google Scholar] [CrossRef]
- Lee, F.X.Z.; Houweling, P.J.; North, K.N.; Quinlan, K.G.R. How does α-actinin-3 deficiency alter muscle function? Mechanistic insights into ACTN3, the ‘gene for speed’. Biochim. Biophys. Acta Mol. Cell Res. 2016, 1863, 686–693. [Google Scholar] [CrossRef] [PubMed]
- North, K.N.; Beggs, A.H. Deficiency of a skeletal muscle isoform of α-actinin (α-actinin-3) in merosin-positive congenital muscular dystrophy. Neuromuscul. Dis. 1996, 6, 229–235. [Google Scholar] [CrossRef]
- North, K.N.; Yang, N.; Wattanasirichaigoon, D.; Mills, M.; Easteal, S.; Beggs, A.H. A common nonsense mutation results in α-actinin-3 deficiency in the general population. Nat. Genet. 1999, 21, 353–354. [Google Scholar] [CrossRef] [PubMed]
- Hogarth, M.W.; Garton, F.C.; Houweling, P.J.; Tukiainen, T.; Lek, M.; Macarthur, D.G.; Seto, J.T.; Quinlan, K.G.R.; Yang, N.; Head, S.I.; et al. Analysis of the ACTN3 heterozygous genotype suggests that α-actinin-3 controls sarcomeric composition and muscle function in a dose-dependent fashion. Hum. Mol. Genet. 2016, 25, 866–877. [Google Scholar] [CrossRef] [PubMed]
- Moreno, V.; Areces, F.; Ruiz-Vicente, D.; Ordovás, J.M.; Del Coso, J. Influence of the ACTN3 R577X genotype on the injury epidemiology of marathon runners. PLoS ONE 2020, 15, e0227548. [Google Scholar] [CrossRef]
- Scott, R.A.; Irving, R.; Irwin, L.; Morrison, E.; Charlton, V.; Austin, K.; Tladi, D.; Deason, M.; Headley, S.A.; Kolkhorst, F.W.; et al. ACTN3 and ACE genotypes in elite Jamaican and US sprinters. Med. Sci. Sports Exerc. 2010, 42, 107–112. [Google Scholar] [CrossRef]
- Yang, N.; Macarthur, D.G.; Wolde, B.; Onywera, V.O.; Boit, M.K.; Lau, S.Y.M.A.; Wilson, R.H.; Scott, R.A.; Pitsiladis, Y.P.; North, K. The ACTN3 R577X polymorphism in east and west African athletes. Med. Sci. Sports Exerc. 2007, 39, 1985–1988. [Google Scholar] [CrossRef]
- Pickering, C.; Kiely, J. ACTN3: More than Just a Gene for Speed. Front. Physiol. 2017, 8, 1080. [Google Scholar] [CrossRef]
- Deschamps, C.L.; Connors, K.E.; Klein, M.S.; Johnsen, V.L.; Shearer, J.; Vogel, H.J.; Devaney, J.M.; Gordish-Dressman, H.; Many, G.M.; Barfield, W.; et al. The ACTN3 R577X polymorphism Is associated with cardiometabolic fitness in healthy young adults. PLoS ONE 2015, 10, e0130644. [Google Scholar] [CrossRef]
- Fiuza-Luces, C.; Ruiz, J.R.; Rodríguez-Romo, G.; Santiago, C.; Gómez-Gallego, F.; Yvert, T.; Cano-Nieto, A.; Garatachea, N.; Morán, M.; Lucia, A. Are “endurance” alleles “survival” alleles? Insights from the ACTN3 R577X polymorphism. PLoS ONE 2011, 6, e17558. [Google Scholar] [CrossRef]
- Yang, N.; MacArthur, D.G.; Gulbin, J.P.; Hahn, A.G.; Beggs, A.H.; Easteal, S.; North, K. ACTN3 genotype is associated with human elite athletic performance. Am. J. Hum. Genet. 2003, 73, 627–631. [Google Scholar] [CrossRef]
- Eynon, N.; Duarte, J.A.; Oliveira, J.; Sagiv, M.; Yamin, C.; Meckel, Y.; Sagiv, M.; Goldhammer, E. ACTN3 R577X polymorphism and israeli top-level athletes. Int. J. Sports Med. 2009, 30, 695–698. [Google Scholar] [CrossRef]
- Eynon, N.; Hanson, E.D.; Lucia, A.; Houweling, P.J.; Garton, F.; North, K.N.; Bishop, D.J. Genes for elite power and sprint performance: ACTN3 leads the way. Sport. Med. 2013, 43, 803–817. [Google Scholar] [CrossRef]
- Alfred, T.; Ben-Shlomo, Y.; Cooper, R.; Hardy, R.; Cooper, C.; Deary, I.J.; Gunnell, D.; Harris, S.E.; Kumari, M.; Martin, R.M.; et al. ACTN3 genotype, athletic status, and life course physical capability: Meta-analysis of the published literature and findings from nine studies. Hum. Mutat. 2011, 32, 1008–1018. [Google Scholar] [CrossRef]
- Roth, S.M.; Walsh, S.; Liu, D.; Metter, E.J.; Ferrucci, L.; Hurley, B.F. The ACTN3 R577X nonsense allele is under-represented in elite-level strength athletes. Eur. J. Hum. Genet. 2008, 16, 391–394. [Google Scholar] [CrossRef]
- Weyerstraß, J.; Stewart, K.; Wesselius, A.; Zeegers, M. Nine genetic polymorphisms associated with power athlete status—A Meta-Analysis. J. Sci. Med. Sport 2018, 21, 213–220. [Google Scholar] [CrossRef]
- Santiago, C.; González-Freire, M.; Serratosa, L.; Morate, F.J.; Meyer, T.; Gómez-Gallego, F.; Lucia, A. ACTN3 genotype in professional soccer players. Br. J. Sports Med. 2008, 42, 71–73. [Google Scholar] [CrossRef]
- Galeandro, V.; Notarnicola, A.; Bianco, A.; Tafuri, S.; Russo, L.; Pesce, V.; Moretti, B.; Petruzzella, V. ACTN3/ACE genotypes and mitochondrial genome in professional soccer players’ performance. J. Biol. Regul. Homeost. Agents 2017, 31, 207–213. [Google Scholar] [PubMed]
- Moreno-Pérez, V.; Machar, R.; Sanz-Rivas, D.; Del Coso, J. ACTN3 R577X genotype in professional and amateur tennis players. J. Strength Cond. Res. 2020, 34, 952–956. [Google Scholar] [CrossRef]
- Ruiz, J.R.; Fernández del Valle, M.; Verde, Z.; Díez-Vega, I.; Santiago, C.; Yvert, T.; Rodríguez-Romo, G.; Gómez-Gallego, F.; Molina, J.J.; Lucia, A. ACTN3 R577X polymorphism does not influence explosive leg muscle power in elite volleyball players. Scand. J. Med. Sci. Sports 2011, 21, e34–e41. [Google Scholar] [CrossRef]
- Papadimitriou, I.D.; Lucia, A.; Pitsiladis, Y.P.; Pushkarev, V.P.; Dyatlov, D.A.; Orekhov, E.F.; Artioli, G.G.; Guilherme, J.P.L.F.; Lancha, A.H.; Ginevičienė, V.; et al. ACTN3 R577X and ACE I/D gene variants influence performance in elite sprinters: A multi-cohort study. BMC Genom. 2016, 17, 285. [Google Scholar] [CrossRef] [PubMed]
- Broos, S.; Van Leemputte, M.; Deldicque, L.; Thomis, M.A. History-dependent force, angular velocity and muscular endurance in ACTN3 genotypes. Eur. J. Appl. Physiol. 2015, 115, 1637–1643. [Google Scholar] [CrossRef] [PubMed]
- Del Coso, J.; Moreno, V.; Gutiérrez-Hellín, J.; Baltazar-Martins, G.; Ruíz-Moreno, C.; Aguilar-Navarro, M.; Lara, B.; Lucía, A. ACTN3 R577X genotype and exercise phenotypes in recreational marathon runners. Genes 2019, 10, 413. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.; Liu, D.; Metter, E.J.; Ferrucci, L.; Roth, S.M. ACTN3 genotype is associated with muscle phenotypes in women across the adult age span. J. Appl. Physiol. 2008, 105, 1486–1491. [Google Scholar] [CrossRef]
- Gentil, P.; Pereira, R.W.; Leite, T.K.M.; Bottaro, M. ACTN3 R577X polymorphism and neuromuscular response to resistance training. J. Sport. Sci. Med. 2011, 10, 393–399. [Google Scholar]
- Norman, B.; Esbjörnsson, M.; Rundqvist, H.; Österlund, T.; Glenmark, B.; Jansson, E. ACTN3 genotype and modulation of skeletal muscle response to exercise in human subjects. J. Appl. Physiol. 2014, 116, 1197–1203. [Google Scholar] [CrossRef]
- Broos, S.; Malisoux, L.; Theisen, D.; van Thienen, R.; Ramaekers, M.; Jamart, C.; Deldicque, L.; Thomis, M.A.; Francaux, M. Evidence for ACTN3 as a speed gene in isolated human muscle fibers. PLoS ONE 2016, 11, e0150594. [Google Scholar] [CrossRef]
- Delmonico, M.J.; Kostek, M.C.; Doldo, N.A.; Hand, B.D.; Walsh, S.; Conway, J.M.; Carignan, C.R.; Roth, S.M.; Hurley, B.F. Alpha-actinin-3 (ACTN3) R577X polymorphism influences knee extensor peak power response to strength training in older men and women. J. Gerontol. A Biol. Sci. Med. Sci. 2007, 62, 206–212. [Google Scholar] [CrossRef]
- Jones, N.; Kiely, J.; Suraci, B.; Collins, D.J.; Lorenzo, D.D.; Pickering, C.; Grimaldi, K.A. A genetic-based algorithm for personalized resistance training. Biol. Sport 2016, 33, 117–126. [Google Scholar] [CrossRef]
- Montgomery, H.E.; Marshall, R.; Hemingway, H.; Myerson, S.; Clarkson, P.; Dollery, C.; Hayward, M.; Holliman, D.E.; Jubb, M.; World, M.; et al. Human gene for physical performance. Nature 1998, 393, 221–222. [Google Scholar] [CrossRef]
- Head, S.I.; Chan, S.; Houweling, P.J.; Quinlan, K.G.R.; Murphy, R.; Wagner, S.; Friedrich, O.; North, K.N. Altered Ca2+ kinetics associated with α-actinin-3 deficiency may explain positive selection for ACTN3 null allele in human evolution. PLoS Genet. 2015, 11, e1004862. [Google Scholar] [CrossRef] [PubMed]
- Friedlander, S.M.; Herrmann, A.L.; Lowry, D.P.; Mepham, E.R.; Lek, M.; North, K.N.; Organ, C.L. ACTN3 allele frequency in humans covaries with global latitudinal gradient. PLoS ONE 2013, 8, e52282. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, I.D.; Papadopoulos, C.; Kouvatsi, A.; Triantaphyllidis, C. The ACTN3 gene in elite greek track and field athletes. Int. J. Sports Med. 2008, 29, 352–355. [Google Scholar] [CrossRef]
- Saunders, C.J.; September, A.V.; Xenophontos, S.L.; Cariolou, M.A.; Anastassiades, L.C.; Noakes, T.D.; Collins, M. No association of the ACTN3 gene R577X polymorphism with endurance performance in Ironman Triathlons. Ann. Hum. Genet. 2007, 71, 777–781. [Google Scholar] [CrossRef]
- Papadimitriou, I.D.; Lockey, S.J.; Voisin, S.; Herbert, A.J.; Garton, F.; Houweling, P.J.; Cieszczyk, P.; Maciejewska-Skrendo, A.; Sawczuk, M.; Massidda, M.; et al. No association between ACTN3 R577X and ACE I/D polymorphisms and endurance running times in 698 Caucasian athletes. BMC Genom. 2018, 19, 13. [Google Scholar] [CrossRef] [PubMed]
- Sessa, F.; Chetta, M.; Petito, A.; Franzetti, M.; Bafunno, V.; Pisanelli, D.; Sarno, M.; Iuso, S.; Margaglione, M. Gene polymorphisms and sport attitude in Italian athletes. Genet. Test. Mol. Biomark. 2011, 15, 285–290. [Google Scholar] [CrossRef]
- Wang, G.; Mikami, E.; Chiu, L.-L.; De Perini, A.; Deason, M.; Fuku, N.; Miyachi, M.; Kaneoka, K.; Murakami, H.; Tanaka, M.; et al. Association analysis of ACE and ACTN3 in elite Caucasian and East Asian swimmers. Med. Sci. Sports Exerc. 2013, 45, 892–900. [Google Scholar] [CrossRef]
- Pickering, C.; Kiely, J.; Grgic, J.; Lucia, A.; Del Coso, J. Can genetic testing identify talent for sport? Genes 2019, 10, 972. [Google Scholar] [CrossRef]
- MacArthur, D.G.; Seto, J.T.; Raftery, J.M.; Quinlan, K.G.; Huttley, G.A.; Hook, J.W.; Lemckert, F.A.; Kee, A.J.; Edwards, M.R.; Berman, Y.; et al. Loss of ACTN3 gene function alters mouse muscle metabolism and shows evidence of positive selection in humans. Nat. Genet. 2007, 39, 1261–1265. [Google Scholar] [CrossRef]
- Seto, J.T.; Quinlan, K.G.R.; Lek, M.; Zheng, X.F.; Garton, F.; MacArthur, D.G.; Hogarth, M.W.; Houweling, P.J.; Gregorevic, P.; Turner, N.; et al. ACTN3 genotype influences muscle performance through the regulation of calcineurin signaling. J. Clin. Invest. 2013, 123, 4255–4263. [Google Scholar] [CrossRef]
- Quinlan, K.G.R.; Seto, J.T.; Turner, N.; Vandebrouck, A.; Floetenmeyer, M.; Macarthur, D.G.; Raftery, J.M.; Lek, M.; Yang, N.; Parton, R.G.; et al. Alpha-actinin-3 deficiency results in reduced glycogen phosphorylase activity and altered calcium handling in skeletal muscle. Hum. Mol. Genet. 2010, 19, 1335–1346. [Google Scholar] [CrossRef]
- MacArthur, D.G.; Seto, J.T.; Chan, S.; Quinlan, K.G.R.; Raftery, J.M.; Turner, N.; Nicholson, M.D.; Kee, A.J.; Hardeman, E.C.; Gunning, P.W.; et al. An Actn3 knockout mouse provides mechanistic insights into the association between alpha-actinin-3 deficiency and human athletic performance. Hum. Mol. Genet. 2008, 17, 1076–1086. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, I.D.; Eynon, N.; Yan, X.; Munson, F.; Jacques, M.; Kuang, J.; Voisin, S.; North, K.N.; Bishop, D.J. A “human knockout” model to investigate the influence of the α-actinin-3 protein on exercise-induced mitochondrial adaptations. Sci. Rep. 2019, 9, 12688. [Google Scholar] [CrossRef]
- Sakuma, K.; Yamaguchi, A. The functional role of calcineurin in hypertrophy, regeneration, and disorders of skeletal muscle. J. Biomed. Biotechnol. 2010, 2010. [Google Scholar] [CrossRef]
- Garcia-Roves, P.M.; Huss, J.; Holloszy, J.O. Role of calcineurin in exercise-induced mitochondrial biogenesis. Am. J. Physiol. Metab. 2006, 290, E1172–E1179. [Google Scholar] [CrossRef] [PubMed]
- Dunn, S.E.; Burns, J.L.; Michel, R.N. Calcineurin is required for skeletal muscle hypertrophy. J. Biol. Chem. 1999, 274, 21908–21912. [Google Scholar] [CrossRef] [PubMed]
- Garton, F.C.; Seto, J.T.; Quinlan, K.G.R.; Yang, N.; Houweling, P.J.; North, K.N. α-Actinin-3 deficiency alters muscle adaptation in response to denervation and immobilization. Hum. Mol. Genet. 2014, 23, 1879–1893. [Google Scholar] [CrossRef]
- Koizumi, K.I. The ACTN3 Gene is a Potential Biomarker for the Risk of Non-Contact Sports Injury in Female Athletes. J. Mol. Biomark. Diagn. 2015, 6. [Google Scholar] [CrossRef]
- Massidda, M.; Voisin, S.; Culigioni, C.; Piras, F.; Cugia, P.; Yan, X.; Eynon, N.; Calò, C.M. ACTN3 R577X polymorphism is associated with the incidence and severity of injuries in professional football players. Clin. J. Sport Med. 2019, 29, 57–61. [Google Scholar] [CrossRef]
- Clos, E.; Pruna, R.; Lundblad, M.; Artells, R.; Esquirol Caussa, J. ACTN3 single nucleotide polymorphism is associated with non-contact musculoskeletal soft-tissue injury incidence in elite professional football players. Knee Surg. Sport. Traumatol. Arthrosc. 2019, 27, 4055–4061. [Google Scholar] [CrossRef]
- Kim, J.H.; Jung, E.S.; Kim, C.-H.; Youn, H.; Kim, H.R. Genetic associations of body composition, flexibility and injury risk with ACE, ACTN3 and COL5A1 polymorphisms in Korean ballerinas. J. Exerc. Nutr. Biochem. 2014, 18, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Qi, B.; Liu, J.Q.; Liu, G.L. Genetic association between ACTN3 polymorphism and risk of non-acute ankle sprain. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Li, Z.; Cao, X.; Xie, C.; Gu, M.; Chen, P.; Yang, X.; Cai, J. The association between the ACTN3 R577X polymorphism and noncontact acute ankle sprains. J. Sports Sci. 2015, 33, 1775–1779. [Google Scholar] [CrossRef] [PubMed]
- Min, S.K.; Lim, S.T.; Kim, C.S. Association of ACTN3 polymorphisms with BMD, and physical fitness of elderly women. J. Phys. Ther. Sci. 2016, 28, 2731–2736. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Schindeler, A.; McDonald, M.M.; Seto, J.T.; Houweling, P.J.; Lek, M.; Hogarth, M.; Morse, A.R.; Raftery, J.M.; Balasuriya, D.; et al. α-Actinin-3 deficiency is associated with reduced bone mass in human and mouse. Bone 2011, 49, 790–798. [Google Scholar] [CrossRef]
- Yan, X.; Eynon, N.; Papadimitriou, I.D.; Kuang, J.; Munson, F.; Tirosh, O.; O’Keefe, L.; Griffiths, L.R.; Ashton, K.J.; Byrne, N.; et al. The gene SMART study: Method, study design, and preliminary findings. BMC Genom. 2017, 18, 821. [Google Scholar] [CrossRef]
- Levinger, I.; Yan, X.; Bishop, D.; Houweling, P.J.; Papadimitriou, I.; Munson, F.; Byrnes, E.; Vicari, D.; Brennan-Speranza, T.C.; Eynon, N. The influence of α-actinin-3 deficiency on bone remodelling markers in young men. Bone 2017, 98, 26–30. [Google Scholar] [CrossRef]
- Clarkson, P.M.; Hubal, M.J. Exercise-induced muscle damage in humans. Am. J. Phys. Med. Rehabil. 2002, 81, S52–S69. [Google Scholar] [CrossRef]
- Furman, J. When exercise causes exertional rhabdomyolysis. J. Am. Acad. Phys. Assist. 2015, 28, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Del Coso, J.; Fernández, D.; Abián-Vicen, J.; Salinero, J.J.; González-Millán, C.; Areces, F.; Ruiz, D.; Gallo, C.; Calleja-González, J.; Pérez-González, B.; et al. Running pace decrease during a marathon is positively related to blood markers of muscle damage. PLoS ONE 2013, 8, e57602. [Google Scholar] [CrossRef]
- Vincent, B.; Windelinckx, A.; Nielens, H.; Ramaekers, M.; Van Leemputte, M.; Hespel, P.; Thomis, M.A. Protective role of α-actinin-3 in the response to an acute eccentric exercise bout. J. Appl. Physiol. 2010, 109, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Pimenta, E.M.; Coelho, D.B.; Cruz, I.R.; Morandi, R.F.; Veneroso, C.E.; De Azambuja Pussieldi, G.; Carvalho, M.R.S.; Silami-Garcia, E.; De Paz Fernández, J.A. The ACTN3 genotype in soccer players in response to acute eccentric training. Eur. J. Appl. Physiol. 2012, 112, 1495–1503. [Google Scholar] [CrossRef] [PubMed]
- Del Coso, J.; Valero, M.; Salinero, J.J.; Lara, B.; Díaz, G.; Gallo-Salazar, C.; Ruiz-Vicente, D.; Areces, F.; Puente, C.; Carril, J.C.; et al. ACTN3 genotype influences exercise-induced muscle damage during a marathon competition. Eur. J. Appl. Physiol. 2017, 117, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Del Coso, J.; Salinero, J.J.; Lara, B.; Gallo-Salazar, C.; Areces, F.; Puente, C.; Herrero, D. ACTN3 X-allele carriers had greater levels of muscle damage during a half-ironman. Eur. J. Appl. Physiol. 2017, 117, 151–158. [Google Scholar] [CrossRef]
- Belli, T.; Crisp, A.H.; Verlengia, R. Greater muscle damage in athletes with ACTN3 R577X (RS1815739) gene polymorphism after an ultra-endurance race: A pilot study. Biol. Sport 2017, 34, 105–110. [Google Scholar] [CrossRef]
- Del Coso, J.; Valero, M.; Salinero, J.J.; Lara, B.; Gallo-Salazar, C.; Areces, F. Optimum polygenic profile to resist exertional rhabdomyolysis during a marathon. PLoS ONE 2017, 12, e0172965. [Google Scholar] [CrossRef]
- Del Coso, J.; Salinero, J.J.; Lara, B.; Gallo-Salazar, C.; Areces, F.; Herrero, D.; Puente, C. Polygenic profile and exercise-induced muscle damage by a competitive half-ironman. J. Strength Cond. Res. 2017, 34, 1400–1408. [Google Scholar] [CrossRef]
- Clarkson, P.M.; Hoffman, E.P.; Zambraski, E.; Gordish-Dressman, H.; Kearns, A.; Hubal, M.; Harmon, B.; Devaney, J.M. ACTN3 and MLCK genotype associations with exertional muscle damage. J. Appl. Physiol. 2005, 99, 564–569. [Google Scholar] [CrossRef]
- Broos, S.; Malisoux, L.; Theisen, D.; Van Thienen, R.; Francaux, M.; Thomis, M.A.; Deldicque, L. The stiffness response of type IIa fibres after eccentric exercise-induced muscle damage is dependent on ACTN3 r577X polymorphism. Eur. J. Sport Sci. 2019, 19, 480–489. [Google Scholar] [CrossRef]
- Venckunas, T.; Skurvydas, A.; Brazaitis, M.; Kamandulis, S.; Snieckus, A.; Moran, C.N. Human alpha-actinin-3 genotype association with exercise-induced muscle damage and the repeated-bout effect. Appl. Physiol. Nutr. Metab. 2012, 37, 1038–1046. [Google Scholar] [CrossRef]
- Tanisawa, K.; Wang, G.; Seto, J.; Verdouka, I.; Twycross-Lewis, R.; Karanikolou, A.; Tanaka, M.; Borjesson, M.; Di Luigi, L.; Dohi, M.; et al. Sport and exercise genomics: The FIMS 2019 consensus statement update. Br. J. Sports Med. 2020. [Google Scholar] [CrossRef] [PubMed]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baltazar-Martins, G.; Gutiérrez-Hellín, J.; Aguilar-Navarro, M.; Ruiz-Moreno, C.; Moreno-Pérez, V.; López-Samanes, Á.; Domínguez, R.; Del Coso, J. Effect of ACTN3 Genotype on Sports Performance, Exercise-Induced Muscle Damage, and Injury Epidemiology. Sports 2020, 8, 99. https://doi.org/10.3390/sports8070099
Baltazar-Martins G, Gutiérrez-Hellín J, Aguilar-Navarro M, Ruiz-Moreno C, Moreno-Pérez V, López-Samanes Á, Domínguez R, Del Coso J. Effect of ACTN3 Genotype on Sports Performance, Exercise-Induced Muscle Damage, and Injury Epidemiology. Sports. 2020; 8(7):99. https://doi.org/10.3390/sports8070099
Chicago/Turabian StyleBaltazar-Martins, Gabriel, Jorge Gutiérrez-Hellín, Millán Aguilar-Navarro, Carlos Ruiz-Moreno, Victor Moreno-Pérez, Álvaro López-Samanes, Raúl Domínguez, and Juan Del Coso. 2020. "Effect of ACTN3 Genotype on Sports Performance, Exercise-Induced Muscle Damage, and Injury Epidemiology" Sports 8, no. 7: 99. https://doi.org/10.3390/sports8070099
APA StyleBaltazar-Martins, G., Gutiérrez-Hellín, J., Aguilar-Navarro, M., Ruiz-Moreno, C., Moreno-Pérez, V., López-Samanes, Á., Domínguez, R., & Del Coso, J. (2020). Effect of ACTN3 Genotype on Sports Performance, Exercise-Induced Muscle Damage, and Injury Epidemiology. Sports, 8(7), 99. https://doi.org/10.3390/sports8070099