Nitrate Supplementation Combined with a Running Training Program Improved Time-Trial Performance in Recreationally Trained Runners

 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Material and Methods
2.1. Experimental Approach to the Problem
2.2. Participants
2.3. Procedure
2.3.1. Experimental Protocol
2.3.2. Wingate Test and Blood Lactate Concentration
2.3.3. Running Time Trial and Blood Lactate Concentration
2.3.4. Training Protocol
3. Statistical Analyses
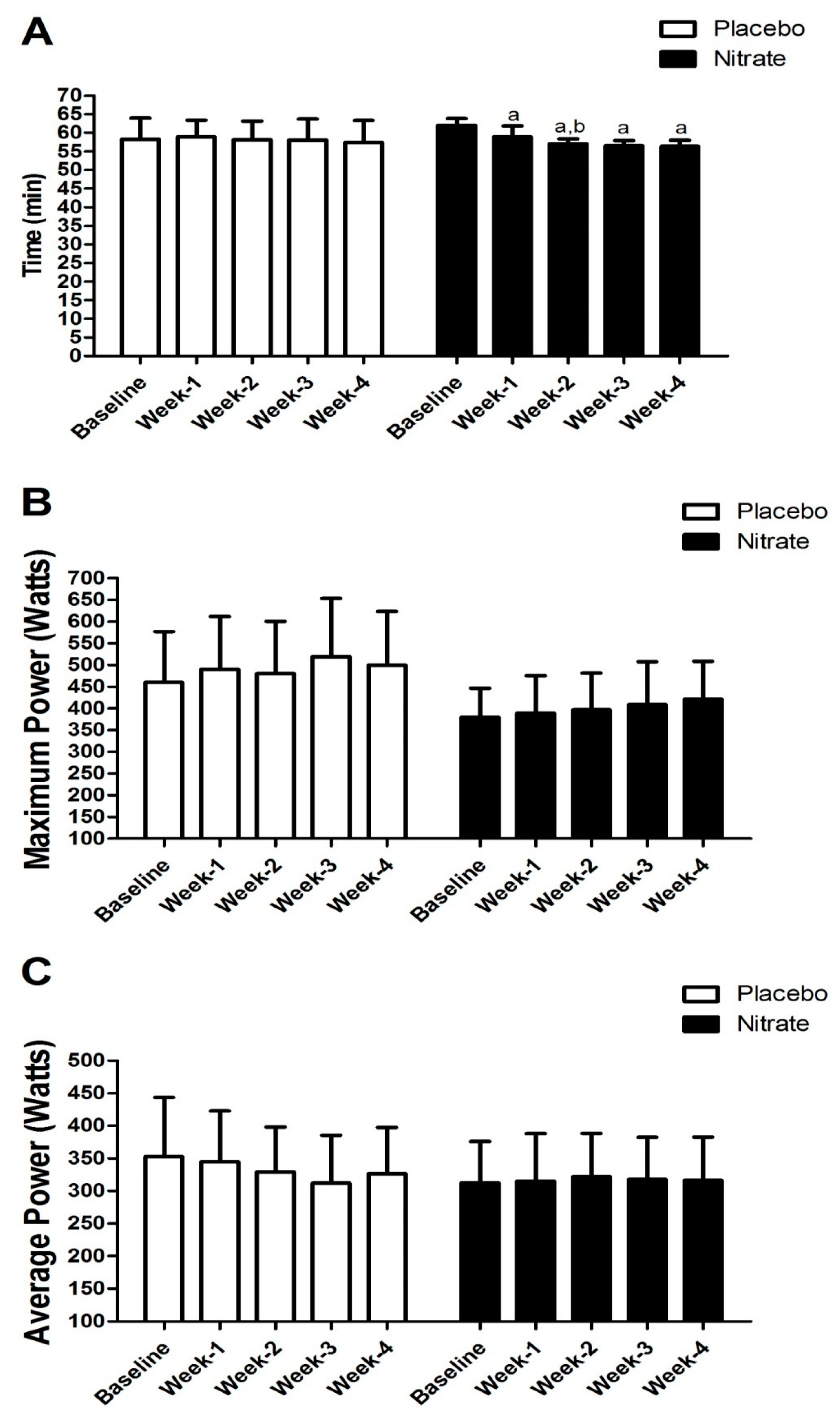
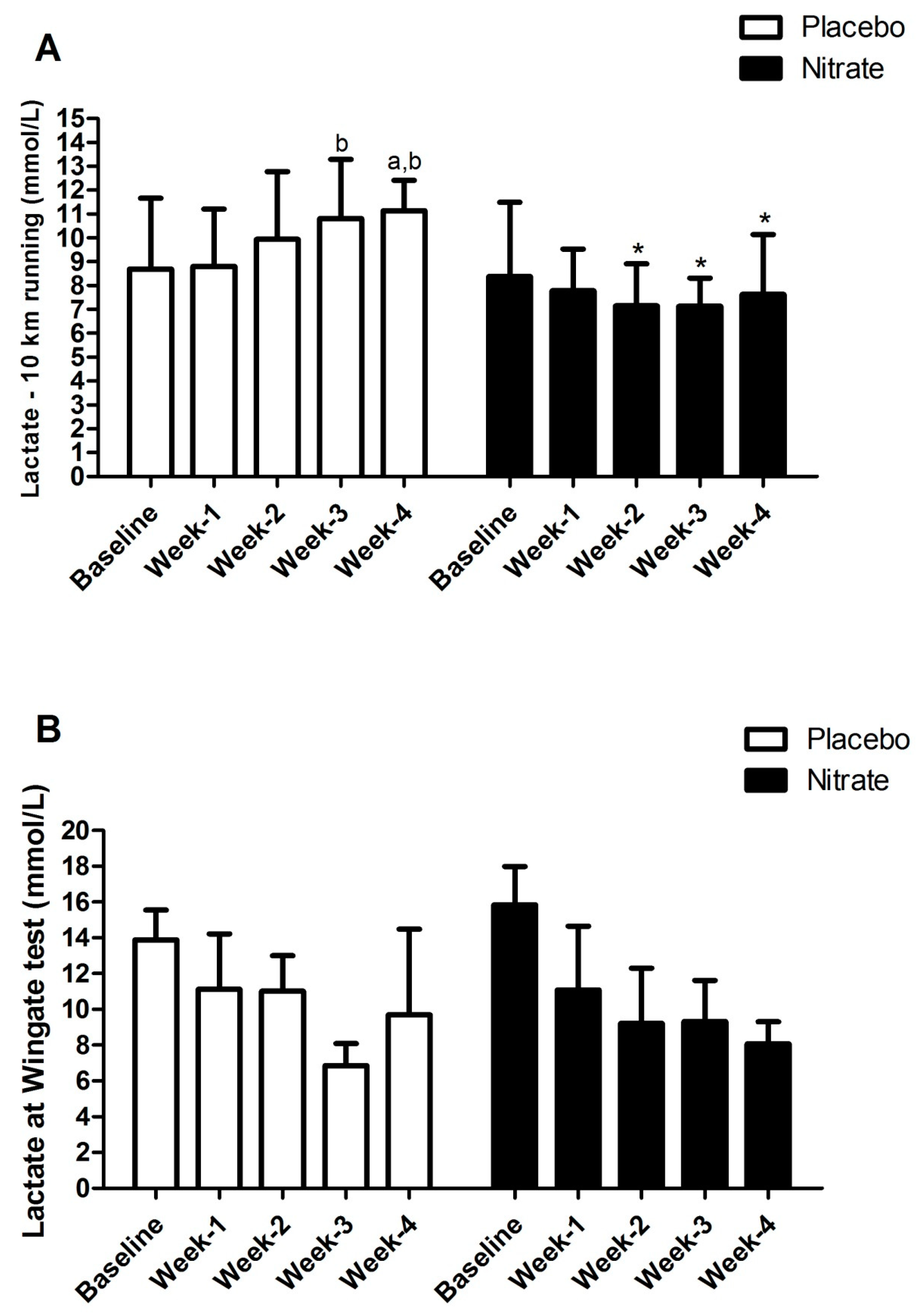
4. Results
5. Discussion
Limitations of Study
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jones, A.M. Dietary nitrate supplementation and exercise performance. Sports Med. 2014, 44, 35–45. [Google Scholar] [CrossRef]
- Tamme, T.; Reinik, M.; Roasto, M.; Juhkam, K.; Tenno, T.; Kiis, A. Nitrates and nitrites in vegetables and vegetable–based products and their intakes by the Estonian population. Food Addit. Contam. 2006, 23, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Clements, W.; Lee, S.R.; Bloomer, R. Nitrate Ingestion: A Review of the Health and Physical Performance Effects. Nutrients 2014, 6, 5224–5264. [Google Scholar] [CrossRef]
- Govoni, M.; Jansson, E.A.; Weitzberg, E.; Lundberg, J.O. The increase in plasma nitrite after a dietary nitrate load is markedly attenuated by an antibacterial mouthwash. Nitric Oxide 2008, 19, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, J.O.; Govoni, M. Inorganic nitrate is a possible source for systemic generation of nitric oxide. Free Radic. Biol. Med. 2004, 37, 395–400. [Google Scholar] [CrossRef]
- Wylie, L.J.; Kelly, J.; Bailey, S.J.; Blackwell, J.R.; Skiba, P.F.; Winyard, P.G.; Jeukendrup, A.E.; Vanhatalo, A.; Jones, A.M. Beetroot juice and exercise: Pharmacodynamic and dose-response relationships. J. Appl. Physiol. 2013, 115, 325–336. [Google Scholar] [CrossRef]
- Wylie, L.J.; Mohr, M.; Krustrup, P.; Jackman, S.R.; Ermdis, G.; Kelly, J.; Black, M.I.; Bailey, S.J.; Vanhatalo, A.; Jones, A.M. Dietary nitrate supplementation improves team sport-specific intense intermittent exercise performance. Eur. J. Appl. Physiol. 2013, 113, 1673–1684. [Google Scholar] [CrossRef]
- Vanhatalo, A.; Bailey, S.J.; Blackwell, J.R.; DiMenna, F.J.; Pavey, T.G.; Wilkerson, D.P.; Benjamin, N.; Winyard, P.G.; Jones, A.M. Acute and chronic effects of dietary nitrate supplementation on blood pressure and the physiological responses to moderate-intensity and incremental exercise. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- Muggeridge, D.J.; Howe, C.C.F.; Spendiff, O.; Pedlar, C.; James, P.E.; Easton, C. A single dose of beetroot juice enhances cycling performance in simulated altitude. Med. Sci. Sports Exerc. 2014, 46, 143–150. [Google Scholar] [CrossRef]
- Thompson, K.G.; Turner, L.; Prichard, J.; Dodd, F.; Kennedy, D.O.; Haskell, C.; Blackwell, J.R.; Jones, A.M. Influence of dietary nitrate supplementation on physiological and cognitive responses to incremental cycle exercise. Respir. Physiol. Neurobiol. 2014, 193, 11–20. [Google Scholar] [CrossRef]
- Bailey, S.J.; Fulford, J.; Vanhatalo, A.; Winyard, P.G.; Blackwell, J.R.; DiMenna, F.J.; Wilkerson, D.P.; Benjamin, N.; Jones, A.M. Dietary nitrate supplementation enhances muscle contractile efficiency during knee-extensor exercise in humans. J. Appl. Physiol. 2010, 109, 135–148. [Google Scholar] [CrossRef]
- Pinna, M.; Roberto, S.; Milia, R.; Marongiu, E.; Olla, S.; Loi, A.; Migliaccio, G.M.; Padulo, J.; Orlandi, C.; Tocco, F.; et al. Effect of beetroot juice supplementation on aerobic response during swimming. Nutrients 2014, 6, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Stamler, J.S.; Meissner, G. Physiology of Nitric Oxide in Skeletal Muscle. Physiol. Rev. 2001, 81, 209–237. [Google Scholar] [CrossRef] [PubMed]
- Larsen, F.J.; Schiffer, T.A.; Borniquel, S.; Sahlin, K.; Ekblom, B.; Lundberg, J.O.; Weitzberg, E. Dietary inorganic nitrate improves mitochondrial efficiency in humans. Cell Metab. 2011, 13, 149–159. [Google Scholar] [CrossRef]
- Lansley, K.E.; Winyard, P.G.; Fulford, J.; Vanhatalo, A.; Bailey, S.J.; Blackwell, J.R.; DiMenna, F.J.; Gilchrist, M.; Benjamin, N.; Jones, A.M. Dietary nitrate supplementation reduces the O2 cost of walking and running: A placebo-controlled study. J. Appl. Physiol. 2011, 110, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Bertuzzi, R.; Lima-Silva, A.E. Pacing Strategy Determinants During a 10 km Running Time Trial: Contributions of Perceived Effort, Physiological, and Muscular Parameters. J. Strength Cond. Res. 2014, 28, 1688–1696. [Google Scholar] [CrossRef] [PubMed]
- Vanhatalo, A.; Fulford, J.; Bailey, S.J.; Blackwell, J.R.; Winyard, P.G.; Jones, A.M. Dietary nitrate reduces muscle metabolic perturbation and improves exercise tolerance in hypoxia. J. Physiol. 2011, 589, 5517–5528. [Google Scholar] [CrossRef]
- Hoon, M.W.; Johnson, N.A.; Chapman, P.G.; Burke, L.M. The Effect of Nitrate Supplementation on Exercise Performance in Healthy Individuals: A Systematic Review and Meta-Analysis. Int. J. Sport Nutr. Exerc. Metab. 2013, 23, 522–532. [Google Scholar] [CrossRef]
- De Castro, T.F.; Manoel, F.D.A.; Figueiredo, D.H.; Figueiredo, D.H.; Machado, F.A. Effect of beetroot juice supplementation on 10 km performance in recreational runners. Appl. Physiol. Nutr. Metab. 2018, 44, 90–94. [Google Scholar] [CrossRef]
- Porcelli, S.; Ramaglia, M.; Bellistri, G.; Pavei, G.; Pugliese, L.; Montorsi, M.; Rasica, L.; Marzorati, M. Aerobic fitness affects the exercise performance responses to nitrate supplementation. Med. Sci. Sports Exerc. 2015, 47, 1643–1651. [Google Scholar] [CrossRef]
- Santana, J.O.; de Freitas, M.C.; dos Santos, D.M.; Rossi, F.E.; Lira, F.S.; Rosa-Neto, J.C.; Caperuto, E.C. Beta-Alanine Supplementation Improved 10-km Running Time Trial in Physically Active Adults. Front. Physiol. 2018, 9, 1–6. [Google Scholar] [CrossRef]
- Hoffman, J.R.; Ratamess, N.A.; Faigenbaum, A.D.; Ross, R.; Kang, J.; Stout, J.R.; Wise, J.A. Short-duration β-alanine supplementation increases training volume and reduces subjective feelings of fatigue in college football players. Nutr. Res. 2008, 28, 31–35. [Google Scholar] [CrossRef]
- Shannon, O.M.; Barlow, M.J.; Duckworth, L.; Williams, E.; Wort, G.; Woods, D.; Siervo, M.; O’Hara, J.P. Dietary nitrate supplementation enhances short but not longer duration running time-trial performance. Eur. J. Appl. Physiol. 2017, 117, 775–785. [Google Scholar] [CrossRef]
- Domínguez, R.; Cuenca, E.; Maté-Muñoz, J.L.; García-Fernández, P.; Serra-Paya, N.; Estevan, M.C.L.; Herreros, P.V.; Garnacho-Castaño, M.V. Effects of beetroot juice supplementation on cardiorespiratory endurance in athletes. A systematic review. Nutrients 2017, 9, 43. [Google Scholar] [CrossRef]
- McMahon, N.F.; Michael Leveritt, B.D.; Toby Pavey, B.G. The Effect of Dietary Nitrate Supplementation on Endurance Exercise Performance in Healthy Adults: A Systematic Review and Meta-Analysis. Sports Med. 2017, 47, 735–756. [Google Scholar] [CrossRef]
- Vassalle, C.; Lubrano, V.; Domenici, C.; L’Abbate, A. Influence of chronic aerobic exercise on microcirculatory flow and nitric oxide in humans. Influence de l’ activite physique reguliere sur la micro-circulation et l’ oxyde nitrique chez des humains. Int. J. Sports Med. 2003, 24, 30–35. [Google Scholar] [CrossRef]
- Naderi, A.; Oliveira, E.P.; Ziegenfuss, T.N.; Willems, M.T. Timing, optimal dose and intake duration of dietary supplements with evidence–based use in sports nutrition. J. Exerc. Nutr. Biochem. 2016, 20, 1–12. [Google Scholar] [CrossRef]
- Wylie, L.J.; Zevallos, J.O.; Isidore, T.; Nyman, L.; Vanhatalo, A.; Bailey, S.J.; Jones, A.M. Dose-dependent effects of dietary nitrate on the oxygen cost of moderate-intensity exercise: Acute vs. chronic supplementation. Nitric Oxide 2016, 57, 30–39. [Google Scholar] [CrossRef]
- Saunders, P.U.; Pyne, D.B.; Telford, R.D.; Hawley, J.A. Factors Affecting Running Economy in Trained Distance Runners. Sports Med. 2004, 34, 465–485. [Google Scholar] [CrossRef]
- Tanaka, K. Lactate-related factors as a critical determinant of endurance. Ann. Physiol. Anthr. 1990, 9, 191–202. [Google Scholar] [CrossRef][Green Version]
- Föhrenbach, R.; Mader, A.; Hollmann, W. Determination of Endurance Capacity and Prediction of Exercise Intensities for Training and Competition in Marathon Runners. Int. J. Sports Med. 1987, 8, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Sjödin, B.; Jacobs, I. Onset of Blood Lactate Accumulation and Marathon Running Performance. Int. J. Sports Med. 1981, 2, 23–26. [Google Scholar] [CrossRef]
- Denadai, B.S.; Ortiz, M.J.; Greco, C.C.; Mello, M.T. Interval training at 95% and 100% of the velocity at VO2 max: Effects on aerobic physiological indexes and running performance. Appl. Physiol. Nutr. Metab. 2006, 31, 737–743. [Google Scholar] [CrossRef]
- Billat, V.L.; Flechet, B.; Petit, B.; Muriaux, G.; Koralsztein, J. Interval training at VO2max: Effects on aerobic performance and overtraining markers. Med. Sci. Sports Exerc. 1999, 31, 156–163. [Google Scholar] [CrossRef]
- Domínguez, R.; Garnacho-Castaño, M.V.; Cuenca, E.; García-Fernández, P.; Muñoz-González, A.; Jesús, F.; Lozano-Estevan, M.C.; Silva, S.F.; Veiga-Herreros, P.; Maté-Munoz, J.L. Effects of Beetroot Juice Supplementation on a 30 s High-Intensity Inertial Cycle Ergometer Test. Nutrients 2017, 9, 1–14. [Google Scholar] [CrossRef]
- Jonvik, K.L.; Nyakayiru, J.; Van Dijk, J.W.; Maase, K.; Ballak, S.B.; Senden, J.M.G.; Van Loon, L.J.C.; Verdijk, L.B. Repeated-sprint performance and plasma responses following beetroot juice supplementation do not differ between recreational, competitive and elite sprint athletes. Eur. J. Sport Sci. 2018, 18, 524–533. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Characteristics | Placebo (n = 8) | Nitrate (n = 8) | p |
|---|---|---|---|
| Age (years) | 30.3 ± 4.5 | 30.0 ± 6.8 | 0.999 |
| Height (m) | 1.73 ± 0.1 | 1.66 ± 0.1 | 0.563 |
| Weight (kg) | 79.5 ± 11.2 | 68.8 ± 10.6 | 0.257 |
| BMI (kg/m2) | 25.65 ± 1.38 | 25.85 ± 1.84 | 0.793 |
| Fat free body mass (kg) | 63.7 ± 11.3 | 52.9 ± 8.5 | 0.071 |
| % Fat | 20.4 ± 4.5 | 23.3 ± 2.4 | 0.176 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santana, J.; Madureira, D.; de França, E.; Rossi, F.; Rodrigues, B.; Fukushima, A.; Billaut, F.; Lira, F.; Caperuto, E. Nitrate Supplementation Combined with a Running Training Program Improved Time-Trial Performance in Recreationally Trained Runners. Sports 2019, 7, 120. https://doi.org/10.3390/sports7050120
Santana J, Madureira D, de França E, Rossi F, Rodrigues B, Fukushima A, Billaut F, Lira F, Caperuto E. Nitrate Supplementation Combined with a Running Training Program Improved Time-Trial Performance in Recreationally Trained Runners. Sports. 2019; 7(5):120. https://doi.org/10.3390/sports7050120
Chicago/Turabian StyleSantana, Jeferson, Diana Madureira, Elias de França, Fabricio Rossi, Bruno Rodrigues, André Fukushima, François Billaut, Fabio Lira, and Erico Caperuto. 2019. "Nitrate Supplementation Combined with a Running Training Program Improved Time-Trial Performance in Recreationally Trained Runners" Sports 7, no. 5: 120. https://doi.org/10.3390/sports7050120
APA StyleSantana, J., Madureira, D., de França, E., Rossi, F., Rodrigues, B., Fukushima, A., Billaut, F., Lira, F., & Caperuto, E. (2019). Nitrate Supplementation Combined with a Running Training Program Improved Time-Trial Performance in Recreationally Trained Runners. Sports, 7(5), 120. https://doi.org/10.3390/sports7050120