
Cardiac Autonomic and Salivary Responses to a Repeated Training Bout in Elite Swimmers



Abstract
:1. Introduction
2. Materials and Methods
2.1. Participant Demographics
2.2. Training Monitoring
2.2.1. Monday a.m. and Tuesday a.m.
2.2.2. Monday p.m.
2.2.3. Training Overview
2.3. Procedures
2.4. Data and Statistical Analysis
3. Results
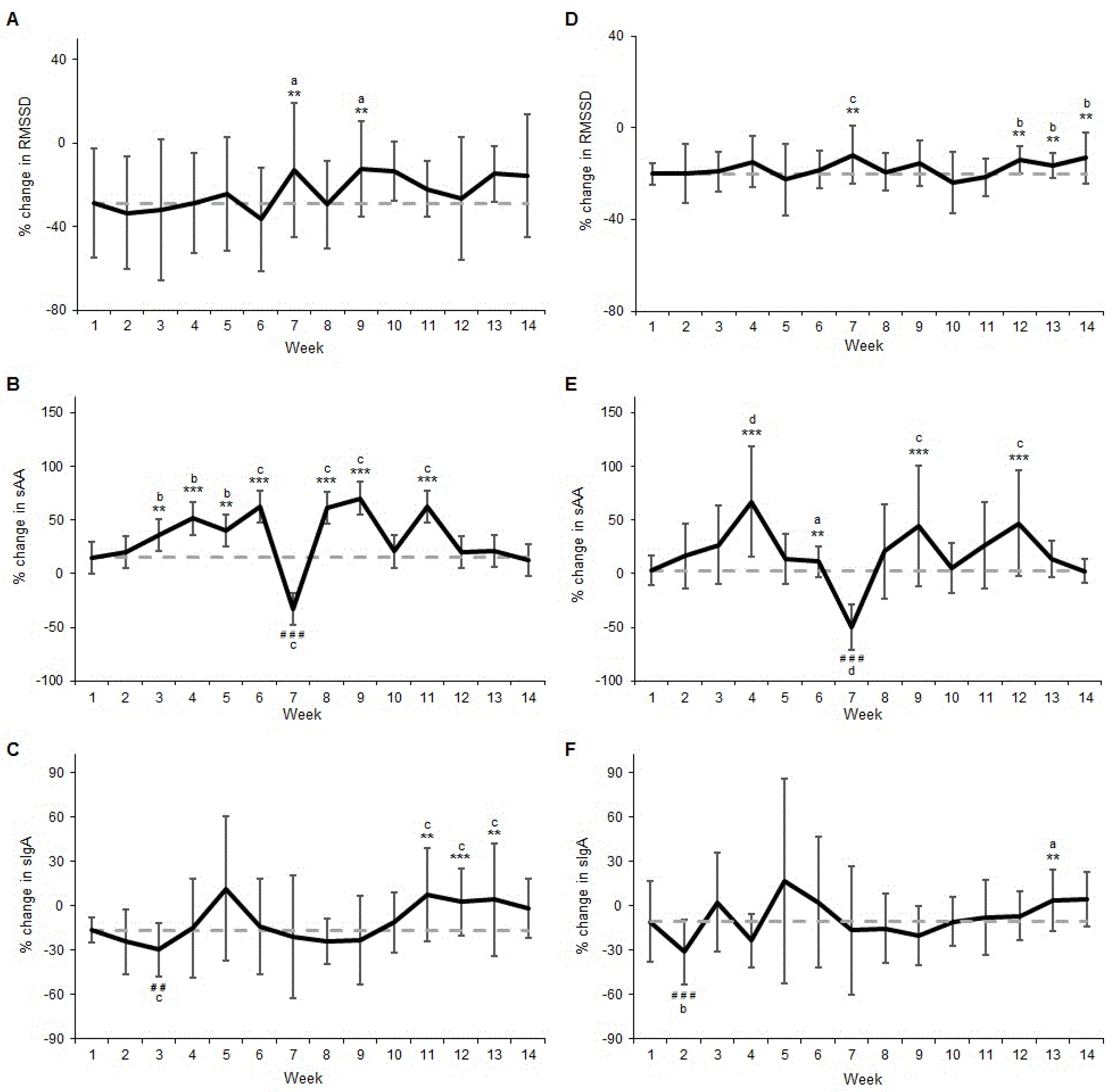
3.1. Change from Monday a.m. to Monday p.m.
3.2. Change from Monday a.m. to Tuesday a.m.
4. Discussion
4.1. Change from Monday a.m. to Monday p.m.
4.2. Change from Monday a.m. to Tuesday a.m.
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Heisterberg, M.F.; Fahrenkrug, J.; Krustrup, P.; Storskov, A.; Kjaer, M.; Andersen, J.L. Extensive monitoring through multiple blood samples in professional soccer players. J. Strength Cond. Res. 2013, 27, 1260–1271. [Google Scholar] [CrossRef] [PubMed]
- Borresen, J.; Lambert, M.I. The quantification of training load, the training response and the effect on performance. Sports Med. 2009, 39, 779–795. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.-L.; Chapman, D.W.; Cronin, J.B.; Newton, M.J.; Gill, N. Fatigue monitoring in high performance sport: A survey of current trends. J. Aust. Strength Cond. 2012, 20, 12–23. [Google Scholar]
- Boullosa, D.A.; Abreu, L.; Nakamura, F.Y.; Munoz, V.E.; Dominguez, E.; Leicht, A.S. Cardiac autonomic adaptations in elite spanish soccer players during preseason. Int. J. Sports Physiol. Perform. 2013, 8, 400–409. [Google Scholar] [PubMed]
- Edmonds, R.; Leicht, A.; McKean, M.; Burkett, B. Daily heart rate variability of paralympic gold medallist swimmers: A 17-week investigation. J. Sport Health Sci. 2014, in press. [Google Scholar] [CrossRef]
- Atlaoui, D.; Pichot, V.; Lacoste, L.; Barale, F.; Lacour, J.R.; Chatard, J.C. Heart rate variability, training variation and performance in elite swimmers. Int. J. Sports Med. 2007, 28, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Stanley, J.; Peake, J.M.; Buchheit, M. Cardiac parasympathetic reactivation following exercise: Implications for training prescription. Sports Med. 2013, 43, 1259–1277. [Google Scholar] [CrossRef] [PubMed]
- Plews, D.J.; Laursen, P.B.; Stanley, J.; Kilding, A.E.; Buchheit, M. Training adaptation and heart rate variability in elite endurance athletes: Opening the door to effective monitoring. Sports Med. 2013, 43, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Moreira, A.; de Moura, N.R.; Coutts, A.; Costa, E.C.; Kempton, T.; Aoki, M.S. Monitoring internal training load and mucosal immune responses in futsal athletes. J. Strength Cond. Res. 2013, 27, 1253–1259. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Kappel, M.; Klokker, M.; Nielsen, H.B.; Secher, N.H. The immune system during exposure to extreme physiologic conditions. Int. J. Sports Med. 1994, 15 (Suppl. 3), S116–S121. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C.; Pedersen, B.K. Exercise and immune function. Recent developments. Sports Med. 1999, 27, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Mackinnon, L.T.; Ginn, E.; Seymour, G.J. Decreased salivary immunoglobulin a secretion rate after intense interval exercise in elite kayakers. Eur. J. Appl. Physiol. Occup. Physiol. 1993, 67, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C. Exercise, infection, and immunity. Int. J. Sports Med. 1994, 15 (Suppl. 3), S131–S141. [Google Scholar] [CrossRef] [PubMed]
- Nater, U.M.; Rohleder, N. Salivary alpha-amylase as a non-invasive biomarker for the sympathetic nervous system: Current state of research. Psychoneuroendocrinology 2009, 34, 486–496. [Google Scholar] [CrossRef] [PubMed]
- Bocanegra, O.L.; Diaz, M.M.; Teixeira, R.R.; Soares, S.S.; Espindola, F.S. Determination of the lactate threshold by means of salivary biomarkers: Chromogranin a as novel marker of exercise intensity. Eur. J. Appl. Physiol. 2012, 112, 3195–3203. [Google Scholar] [CrossRef] [PubMed]
- Steerenberg, P.A.; van Asperen, I.A.; van Nieuw Amerongen, A.; Biewenga, A.; Mol, D.; Medema, G.J. Salivary levels of immunoglobulin a in triathletes. Eur. J. Oral. Sci. 1997, 105, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Walsh, N.P.; Blannin, A.K.; Clark, A.M.; Cook, L.; Robson, P.J.; Gleeson, M. The effects of high-intensity intermittent exercise on saliva iga, total protein and alpha-amylase. J. Sports Sci. 1999, 17, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Papacosta, E.; Nassis, G.P. Saliva as a tool for monitoring steroid, peptide and immune markers in sport and exercise science. J. Sci. Med. Sport 2011, 14, 424–434. [Google Scholar] [CrossRef] [PubMed]
- Hedelin, R.; Wiklund, U.; Bjerle, P.; Henriksson-Larsen, K. Cardiac autonomic imbalance in an overtrained athlete. Med. Sci. Sports Exerc. 2000, 32, 1531–1533. [Google Scholar] [CrossRef] [PubMed]
- Mourot, L.; Bouhaddi, M.; Perrey, S.; Cappelle, S.; Henriet, M.T.; Wolf, J.P.; Rouillon, J.D.; Regnard, J. Decrease in heart rate variability with overtraining: Assessment by the poincare plot analysis. Clin. Physiol. Funct. Imaging 2004, 24, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Edmonds, R.; Burkett, B.; Leicht, A.; McKean, M. Effect of chronic training on heart rate variability, salivary iga and salivary alpha-amylase in elite swimmers with a disability. PLoS One 2015, 10, e0127749. [Google Scholar] [CrossRef] [PubMed]
- Edmonds, R.C.; Sinclair, W.H.; Leicht, A.S. Effect of a training week on heart rate variability in elite youth rugby league players. Int. J. Sports Med. 2013, 34, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, J.; Armitage, M.; Jehanli, A.; Browne, A. Mucosal immunity and self-reported upper respiratory symptoms in a cohort of premier league academy soccer players. Int. J. Exerc. Sci. Conf. Proc. 2013, 10, Article 45. [Google Scholar]
- Dunbar, J.; Jehanli, A.; Skelhorn, S. Investigating the use of a point of care siga test in the sporting environment. In Proceedings of the 10th International Society of Exercise Immunology, Oxford, UK, 11–13 July 2011; p. 85.
- Buchheit, M.; Laursen, P.B.; Al Haddad, H.; Ahmaidi, S. Exercise-induced plasma volume expansion and post-exercise parasympathetic reactivation. Eur. J. Appl. Physiol. 2009, 105, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Stanley, J.; Halliday, A.; D’Auria, S.; Buchheit, M.; Leicht, A.S. Effect of sauna-based heat acclimation on plasma volume and heart rate variability. Eur. J. Appl. Physiol. 2015, 115, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.G.; Marshall, S.W.; Batterham, A.M.; Hanin, J. Progressive statistics for studies in sports medicine and exercise science. Med. Sci. Sports Exerc. 2009, 41, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Massin, M.M.; Maeyns, K.; Withofs, N.; Ravet, F.; Gerard, P. Circadian rhythm of heart rate and heart rate variability. Arch. Dis. Child. 2000, 83, 179–182. [Google Scholar] [CrossRef] [PubMed]
- Hayano, J.; Sakakibara, Y.; Yamada, M.; Kamiya, T.; Fujinami, T.; Yokoyama, K.; Watanabe, Y.; Takata, K. Diurnal variations in vagal and sympathetic cardiac control. Am. J. Physiol. 1990, 258, H642–H646. [Google Scholar] [PubMed]
- Chatterton, R.T., Jr.; Vogelsong, K.M.; Lu, Y.C.; Ellman, A.B.; Hudgens, G.A. Salivary alpha-amylase as a measure of endogenous adrenergic activity. Clin. Physiol. 1996, 16, 433–448. [Google Scholar] [CrossRef] [PubMed]
- Ljungberg, G.; Ericson, T.; Ekblom, B.; Birkhed, D. Saliva and marathon running. Scand. J. Med. Sci. Sports 1997, 7, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Nater, U.M.; Rohleder, N.; Schlotz, W.; Ehlert, U.; Kirschbaum, C. Determinants of the diurnal course of salivary alpha-amylase. Psychoneuroendocrinology 2007, 32, 392–401. [Google Scholar] [CrossRef] [PubMed]
- McLean, B.D.; Coutts, A.J.; Kelly, V.; McGuigan, M.R.; Cormack, S.J. Neuromuscular, endocrine, and perceptual fatigue responses during different length between-match microcycles in professional rugby league players. Int. J. Sports Physiol. Perform. 2010, 5, 367–383. [Google Scholar] [PubMed]
- Carvalho Leme, L.; Milanez, V.; Oliveira, R.; de Paulo Ramos, S.; Leicht, A.; Nakamura, F. The influence of a weekend with passive rest on the psychological and autonomic recovery in professional handball players. Kinesiology 2015, 47, 44–52. [Google Scholar]
- Hautala, A.; Tulppo, M.P.; Makikallio, T.H.; Laukkanen, R.; Nissila, S.; Huikuri, H.V. Changes in cardiac autonomic regulation after prolonged maximal exercise. Clin. Physiol. 2001, 21, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Garet, M.; Tournaire, N.; Roche, F.; Laurent, R.; Lacour, J.R.; Barthelemy, J.C.; Pichot, V. Individual interdependence between nocturnal ans activity and performance in swimmers. Med. Sci. Sports Exerc. 2004, 36, 2112–2118. [Google Scholar] [CrossRef] [PubMed]
- Pichot, V.; Roche, F.; Gaspoz, J.M.; Enjolras, F.; Antoniadis, A.; Minini, P.; Costes, F.; Busso, T.; Lacour, J.R.; Barthelemy, J.C. Relation between heart rate variability and training load in middle-distance runners. Med. Sci. Sports Exerc. 2000, 32, 1729–1736. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C. Immune response to heavy exertion. J. Appl. Physiol. 1997, 82, 1385–1394. [Google Scholar] [PubMed]
- Hoffman-Goetz, L.; Pedersen, B.K. Exercise and the immune system: A model of the stress response? Immunol. Today 1994, 15, 382–387. [Google Scholar] [CrossRef]
- Nieman, D.C. Exercise immunology: Practical applications. Int. J. Sports Med. 1997, 18 (Suppl. 1), S91–S100. [Google Scholar] [CrossRef] [PubMed]
- Blannin, A.K.; Robson, P.J.; Walsh, N.P.; Clark, A.M.; Glennon, L.; Gleeson, M. The effect of exercising to exhaustion at different intensities on saliva immunoglobulin a, protein and electrolyte secretion. Int. J. Sports Med. 1998, 19, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Sari-Sarraf, V.; Reilly, T.; Doran, D.A.; Atkinson, G. The effects of single and repeated bouts of soccer-specific exercise on salivary iga. Arch. Oral. Biol. 2007, 52, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Nehlsen-Cannarella, S.L.; Nieman, D.C.; Fagoaga, O.R.; Kelln, W.J.; Henson, D.A.; Shannon, M.; Davis, J.M. Saliva immunoglobulins in elite women rowers. Eur. J. Appl. Physiol. 2000, 81, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Allgrove, J.E.; Gomes, E.; Hough, J.; Gleeson, M. Effects of exercise intensity on salivary antimicrobial proteins and markers of stress in active men. J. Sports Sci. 2008, 26, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Sari-Sarraf, V.; Reilly, T.; Doran, D.A. Salivary iga response to intermittent and continuous exercise. Int. J. Sports Med. 2006, 27, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Edmonds, R.C.; Leicht, A.S.; McKean, M.; Burkett, B. Daily heart rate variability during an 18-day staging camp in paralympic medallist swimmers. J. Exerc. Physiol. 2014, 17, 84–92. [Google Scholar]
- Plews, D.J.; Laursen, P.B.; Kilding, A.E.; Buchheit, M. Heart rate variability in elite triathletes, is variation in variability the key to effective training? A case comparison. Eur. J. Appl. Physiol. 2012, 112, 3729–3741. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Athlete | Sex | Age (years) | Impairment | Athlete (Classification) a | National Experience (years) b |
|---|---|---|---|---|---|
| Athlete 1 | F | 20 | Intellectual | S14 | 2 |
| Athlete 2 | M | 29 | Amputee | SM10 | 11 |
| Athlete 3 | M | 17 | Vision | S13 | 1 |
| Athlete 4 | F | 15 | Cerebral Palsy | S8 | 1 |
| Athlete 5 | M | 24 | Amputee | S8 | 6 |
| Athlete 6 | M | 27 | Neuromuscular | S10 | 11 |
| Athlete 7 | M | 16 | Amputee | S9 | 1 |
| Athlete 8 | M | 19 | Vision | S13 | 2 |
| Training Phase | Week | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | |
| Aerobic Maintenance | 28 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Aerobic Capacity | - | 34 | 43 | 47 | 48 | - | - | - | - | - | - | - | - | - | - |
| Anaerobic Capacity | - | - | - | - | - | 45 | 48 | 48 | 45 | 47 | - | - | - | - | - |
| Power and Speed | - | - | - | - | - | - | - | - | - | - | 46 | 35 | - | - | - |
| Taper | - | - | - | - | - | - | - | - | - | - | - | - | 28 | 29 | - |
| Competition | X | X | X | X | |||||||||||
| Swimming Test | X | X | |||||||||||||
| Change from Monday a.m. to Monday p.m. | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Week | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 |
| HR | 26.88 | 30.71 | 32.09 | 28.12 | 27.21 | 35.85 | 20.74 | 23.02 | 23.33 | 21.90 | 31.55 | 30.47 | 19.80 | 17.54 |
| (±SD) | (±23.5) | (±29.8) | (±31.5) | (±20.0) | (±20.9) | (±26.4) | (±14.6) | (±13.8) | (±18.0) | (±17.4) | (±19.8) | (±21.0) | (±14.2) | (±11.4) |
| ES | 0.12 | 0.14 | 0.07 | 0.02 | 0.33 | −0.21 | −0.11 | −0.12 | −0.17 | 0.20 | 0.15 | −0.25 | −0.33 | |
| QI | Unclear | Unclear | Unclear | Unclear | Possibly +ve | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | |
| 37/56/7 | 45/38/17 | 32/51/16 | 25/56/19 | 72/24/4 | 12/34/54 | 16/43/41 | 21/36/43 | 7/44/49 | 54/39/7 | 46/45/9 | 12/30/58 | 6/24/70 | ||
| HF(nu) | −21.61 | −13.03 | −26.67 | −22.27 | −22.75 | −24.32 | −2.84 | −7.31 | −22.62 | −10.00 | −18.58 | −14.11 | 2.43 | −5.68 |
| (±SD) | (±57.2) | (±53.7) | (±46.1) | (±44.0) | (±35.4) | (±70.2) | (±53.1) | (±86.7) | (±31.9) | (±38.7) | (±33.2) | (±63.3) | (±59.9) | (±74.3) |
| ES | 0.08 | −0.03 | 0.05 | 0.11 | −0.32 | 0.33 | −0.07 | 0.15 | 0.34 | 0.18 | 0.11 | 0.39 | 0.25 | |
| QI | Unclear | Unclear | Trivial | Unclear | Possibly −ve | Possibly +ve | Unclear | Unclear | Likely +ve | Unclear | Unclear | Likely +ve | Unclear | |
| 33/56/12 | 14/65/21 | 11/87/2 | 41/45/15 | 4/25/71 | 71/24/5 | 16/52/32 | 46/35/18 | 76/22/3 | 51/44/6 | 37/54/8 | 83/16/1 | 60/31/9 | ||
| SD1 | −26.71 | −32.74 | −32.05 | −28.88 | −22.10 | −36.53 | −13.35 | −33.77 | −10.05 | −12.43 | −19.65 | −22.92 | −19.53 | −13.65 |
| (±SD) | (±26.6) | (±26.7) | (±33.7) | (±24.1) | (±25.9) | (±24.8) | (±20.2) | (±25.8) | (±21.5) | (±14.2) | (±14.9) | (±27.2) | (±15.9) | (±28.0) |
| ES | −0.17 | −0.23 | −0.01 | 0.16 | −0.23 | 0.38 | −0.13 | 0.44 | 0.42 | 0.28 | 0.16 | 0.28 | 0.35 | |
| QI | Trivial | Possibly −ve | Unclear | Unclear | Possibly −ve | Likely +ve | Unclear | Likely +ve | Unclear | Unclear | Unclear | Possibly +ve | Unclear | |
| 1/51/47 | 5/35/59 | 12/73/14 | 48/39/13 | 1/33/66 | 77/19/4 | 7/54/40 | 79/17/4 | 74/19/7 | 61/28/11 | 48/33/18 | 66/29/5 | 68/23/9 | ||
| α1 | 44.64 | 38.18 | 50.24 | 58.37 | 19.85 | 65.13 | 33.21 | 57.79 | 35.43 | 29.47 | 49.97 | 71.56 | 33.13 | 33.07 |
| (±SD) | (±55.4) | (±53.6) | (±47.8) | (±60.7) | (±34.6) | (±85.3) | (±84.0) | (±78.8 ) | (±57.2) | (±55.3) | (±61.9) | (±91.4) | (±87.5) | (±53.6) |
| ES | −0.08 | 0.13 | 0.21 | −0.35 | 0.21 | −0.32 | 0.10 | −0.13 | −0.27 | 0.08 | 0.28 | −0.31 | −0.17 | |
| QI | Unclear | Unclear | Unclear | Unclear | Possibly +ve | Possibly −ve | Unclear | Unclear | Unclear | Unclear | Possibly +ve | Unclear | Unclear | |
| 7/64/28 | 43/46/11 | 55/39/6 | 8/24/69 | 57/39/5 | 5/26/69 | 38/48/15 | 17/39/44 | 11/28/61 | 36/47/17 | 68/29/3 | 8/26/66 | 9/41/49 | ||
| Change from Monday a.m. to Tuesday a.m. | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Week | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 |
| HR | 5.45 | 6.86 | 6.16 | 7.73 | 3.98 | 9.44 | 7.75 | 7.10 | 6.29 | 9.26 | 8.34 | 6.43 | 3.56 | 5.28 |
| (±SD) | (±12.3) | (±8.5) | (±9.9) | (±7.6) | (±13.9) | (±10.7) | (±16.1) | (±9.1) | (±12.9) | (±7.6) | (±8.5) | (±10.0) | (±6.6) | (±5.8) |
| ES | 0.12 | 0.07 | 0.18 | −0.12 | 0.28 | 0.14 | 0.13 | 0.06 | 0.29 | 0.22 | 0.08 | −0.10 | 0.03 | |
| QI | Trivial | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | |
| 34/63/3 | 39/33/28 | 51/40/9 | 20/37/43 | 60/26/14 | 47/29/25 | 46/30/24 | 40/29/31 | 62/27/11 | 54/28/18 | 30/62/8 | 25/33/42 | 31/43/26 | ||
| HF(nu) | −2.10 | −8.38 | −11.94 | −5.20 | −7.40 | 11.80 | 18.43 | −13.91 | −6.62 | −19.42 | −18.04 | −15.17 | 7.55 | 11.17 |
| (±SD) | (±40.3) | (±35.4) | (±34.5) | (±39.8) | (±35.6) | (±40.4) | (±55.4) | (±22.4) | (±39.5) | (±25.9) | (±20.3) | (±29.3) | (±46.4) | (±79.8) |
| ES | −0.03 | −0.07 | 0.02 | −0.04 | 0.26 | 0.33 | −0.06 | −0.02 | −0.21 | −0.14 | −0.14 | 0.21 | 0.10 | |
| QI | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | Trivial | Unclear | Unclear | |
| 25/44/31 | 18/47/34 | 28/49/23 | 34/26/39 | 60/30/10 | 69/24/6 | 20/47/33 | 34/28/38 | 9/37/55 | 11/46/43 | 3/59/38 | 53/27/20 | 41/37/22 | ||
| SD1 | −18.67 | −21.80 | −19.03 | −14.80 | −17.52 | −18.08 | −2.83 | −19.83 | −15.03 | −23.81 | −20.00 | −13.11 | −16.04 | −11.61 |
| (±SD) | (±6.4) | (±11.9) | (±8.7) | (±11.1) | (±25.5) | (±8.1) | (±26.6) | (±8.7) | (±9.6) | (±13.3) | (±8.8) | (±6.5) | (±5.5) | (±10.8) |
| ES | −0.53 | −0.08 | 0.47 | −0.25 | 0.06 | 1.70 | −0.19 | 0.46 | −0.86 | −0.22 | 0.74 | 0.36 | 0.89 | |
| QI | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | Likely −ve | Unclear | Likely +ve | Unclear | Likely +ve | |
| 10/15/74 | 29/30/41 | 67/16/17 | 33/14/53 | 39/32/29 | 90/4/6 | 23/26/51 | 70/17/13 | 5/8/88 | 16/30/54 | 88/8/4 | 67/22/11 | 92/6/3 | ||
| α1 | 12.34 | 4.18 | 19.60 | 18.61 | 9.45 | 22.15 | 4.08 | 46.60 | 4.67 | 17.58 | 29.94 | 22.26 | 11.40 | 20.91 |
| (±SD) | (±41.5) | (±33.7) | (±29.1) | (±25.2) | (±39.3) | (±58.2) | (±49.0) | (±43.3) | (±24.3) | (±28.3) | (±29.8) | (±40.3) | (±23.7) | (±56.1) |
| ES | −0.17 | 0.23 | 0.23 | −0.09 | 0.14 | −0.28 | 0.75 | −0.12 | 0.20 | 0.47 | 0.25 | 0.07 | 0.14 | |
| QI | Unclear | Unclear | Unclear | Unclear | Unclear | Unclear | Likely +ve | Unclear | Unclear | Likely +ve | Possibly +ve | Unclear | Unclear | |
| 7/45/48 | 56/31/13 | 56/29/15 | 31/25/44 | 47/31/22 | 10/28/62 | 94/5/1 | 24/31/45 | 54/37/10 | 80/16/4 | 65/33/2 | 38/38/25 | 46/34/20 | ||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Edmonds, R.; Leicht, A.; Burkett, B.; McKean, M. Cardiac Autonomic and Salivary Responses to a Repeated Training Bout in Elite Swimmers. Sports 2016, 4, 13. https://doi.org/10.3390/sports4010013
Edmonds R, Leicht A, Burkett B, McKean M. Cardiac Autonomic and Salivary Responses to a Repeated Training Bout in Elite Swimmers. Sports. 2016; 4(1):13. https://doi.org/10.3390/sports4010013
Chicago/Turabian StyleEdmonds, Rohan, Anthony Leicht, Brendan Burkett, and Mark McKean. 2016. "Cardiac Autonomic and Salivary Responses to a Repeated Training Bout in Elite Swimmers" Sports 4, no. 1: 13. https://doi.org/10.3390/sports4010013
APA StyleEdmonds, R., Leicht, A., Burkett, B., & McKean, M. (2016). Cardiac Autonomic and Salivary Responses to a Repeated Training Bout in Elite Swimmers. Sports, 4(1), 13. https://doi.org/10.3390/sports4010013