
Combined Aerobic Exercise with Intermittent Fasting Is Effective for Reducing mTOR and Bcl-2 Levels in Obese Females

 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Intermittent Fasting Protocol
2.3. Aerobic Exercise Protocol
2.4. Body Composition Assessment
2.5. Blood Sampling and Biochemical Analysis
2.6. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). Obesity and Overweight; WHO Press: Geneva, Switzerland, 2020. [Google Scholar]
- Basic Health Research (Riskesdas). National Report on Basic Health Research; Ministry of Health Indonesia: Jakarta, Indonesia, 2018.
- Ayuningtyas, D.; Kusuma, D.; Amir, V.; Tjandrarini, D.H.; Andarwati, P. Disparities in Obesity Rates among Adults: Analysis of 514 Districts in Indonesia. Nutrients 2022, 14, 3332. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.L.; Sinha, S. Obesity and aging: Molecular mechanisms and therapeutic approaches. Ageing Res. Rev. 2021, 67, 101268. [Google Scholar] [CrossRef] [PubMed]
- Menon, K.; de Courten, B.; Ademi, Z.; Owen, A.J.; Liew, D.; Zomer, E. Estimating the benefits of obesity prevention on productivity: An Australian perspective. Int. J. Obes. 2022, 46, 1463–1469. [Google Scholar] [CrossRef] [PubMed]
- Romieu, I.; Dossus, L.; Barquera, S.; Blottière, H.M.; Franks, P.W.; Gunter, M.; Hwalla, N.; Hursting, S.D.; Leitzmann, M.; Margetts, B.; et al. Energy balance and obesity: What are the main drivers? Cancer Causes Control 2017, 28, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Rejeki, P.S.; Pranoto, A.; Rahmanto, I.; Izzatunnisa, N.; Yosika, G.F.; Hernaningsih, Y.; Wungu, C.D.K.; Halim, S. The Positive Effect of Four-Week Combined Aerobic–Resistance Training on Body Composition and Adipokine Levels in Obese Females. Sports 2023, 11, 90. [Google Scholar] [CrossRef] [PubMed]
- Merawati, D.; Sugiharto; Susanto, H.; Taufiq, A.; Pranoto, A.; Amelia, D.; Rejeki, P.S. Dynamic of irisin secretion change after moderate-intensity chronic physical exercise on obese female. J. Basic. Clin. Physiol. Pharmacol. 2023, 34, 539–547. [Google Scholar] [CrossRef]
- Susanto, H.; Sugiharto; Taufiq, A.; Pranoto, A.; Purnomo, J.D.T. Dynamic alteration of plasma levels of betatrophin in younger female onset obesity post acute moderate-intensity exercise training. Saudi J. Biol. Sci. 2023, 30, 103546. [Google Scholar] [CrossRef] [PubMed]
- Pranoto, A.; Cahyono, M.B.A.; Yakobus, R.; Izzatunnisa, N.; Ramadhan, R.N.; Rejeki, P.S.; Miftahussurur, M.; Effendi, W.I.; Wungu, C.D.K.; Yamaoka, Y. Long-Term Resistance–Endurance Combined Training Reduces Pro-Inflammatory Cytokines in Young Adult Females with Obesity. Sports 2023, 11, 54. [Google Scholar] [CrossRef]
- Park, H.J.; Rhie, S.J.; Shim, I. The effects of physical exercise therapy on weight control: Its regulation of adipocyte physiology and metabolic capacity. J. Exerc. Rehabil. 2023, 19, 141–148. [Google Scholar] [CrossRef]
- Welton, S.; Minty, R.; O’Driscoll, T.; Willms, H.; Poirier, D.; Madden, S.; Kelly, L. Intermittent fasting and weight loss: Systematic review. Can. Fam. Physician 2020, 66, 117–125. [Google Scholar]
- Alhamdan, B.A.; Garcia-Alvarez, A.; Alzahrnai, A.H.; Karanxha, J.; Stretchberry, D.R.; Contrera, K.J.; Utria, A.F.; Cheskin, L.J. Alternate-day versus daily energy restriction diets: Which is more effective for weight loss? A systematic review and meta-analysis. Obes. Sci. Pract. 2016, 2, 293–302. [Google Scholar] [PubMed]
- Mattson, M.P.; Longo, V.D.; Harvie, M. Impact of intermittent fasting on health and disease processes. Ageing Res. Rev. 2017, 39, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Song, D.K.; Kim, Y.W. Beneficial effects of intermittent fasting: A narrative review. J. Yeungnam Med. Sci. 2023, 40, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.D.; Di Tano, M.; Mattson, M.P.; Guidi, N. Intermittent and periodic fasting, longevity and disease. Nat. Aging 2021, 1, 47–59. [Google Scholar] [CrossRef]
- Qian, S.; Wei, Z.; Yang, W.; Huang, J.; Yang, Y.; Wang, J. The role of BCL-2 family proteins in regulating apoptosis and cancer therapy. Front. Oncol. 2022, 12, 985363. [Google Scholar] [CrossRef]
- Kaloni, D.; Diepstraten, S.T.; Strasser, A.; Kelly, G.L. BCL-2 protein family: Attractive targets for cancer therapy. Apoptosis 2023, 28, 20–38. [Google Scholar] [CrossRef] [PubMed]
- Koohshoori, Y.S.; Marandi, S.M.; Kargarfard, M.; Vaseghi, G.; Moshtaghian, S.J. The Effect of 4 Weeks Aerobic Exercise Training with Detraining Courses in Various Prevention Phases on BCl-2 and BAX Genes Expression and Proteins. Int. J. Prev. Med. 2023, 14, 79. [Google Scholar] [PubMed]
- Raji-Amirhasani, A.; Khaksari, M.; Soltani, Z.; Saberi, S.; Iranpour, M.; Darvishzadeh Mahani, F.; Hajializadeh, Z.; Sabet, N. Beneficial effects of time and energy restriction diets on the development of experimental acute kidney injury in Rat: Bax/Bcl-2 and histopathological evaluation. BMC Nephrol. 2023, 24, 59. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, B.K.; Lamming, D.W. The Mechanistic Target of Rapamycin: The Grand ConducTOR of Metabolism and Aging. Cell Metab. 2016, 23, 990–1003. [Google Scholar] [CrossRef]
- Lee, H. Phosphorylated mTOR Expression Profiles in Human Normal and Carcinoma Tissues. Dis. Markers 2017, 2017, 1397063. [Google Scholar] [CrossRef]
- Cai, H.; Dong, L.Q.; Liu, F. Recent Advances in Adipose mTOR Signaling and Function: Therapeutic Prospects. Trends Pharmacol. Sci. 2016, 37, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Kjøbsted, R.; Hingst, J.R.; Fentz, J.; Foretz, M.; Sanz, M.N.; Pehmøller, C.; Shum, M.; Marette, A.; Mounier, R.; Treebak, J.T.; et al. AMPK in skeletal muscle function and metabolism. FASEB J. 2018, 32, 1741–1777. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.K.; Kleimbaun, A.P. Design Methodology for Randomized Clinical Trials; Family Health International: Arlington, VA, USA, 1985; pp. 24–25. [Google Scholar]
- Bae, J.Y.; Shin, K.O.; Woo, J.; Woo, S.H.; Jang, K.S.; Lee, Y.H.; Kang, S. Exercise and dietary change ameliorate high fat diet induced obesity and insulin resistance via mTOR signaling pathway. J. Exerc. Nutr. Biochem. 2016, 20, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Sari, A.R.; Risdayanto, R.D.; Pradipta, M.H.; Qorni, U.A.; Rejeki, P.S.; Argarini, R.; Halim, S.; Pranoto, A. Impact of Time-Resricted Feeding and Aerobic Exercise Combination on Promotes Myokine Levels and Improve Body Composition in Obese Women. Retos 2024, 53, 1–10. [Google Scholar] [CrossRef]
- Delli Paoli, G.; van de Laarschot, D.; Friesema, E.C.H.; Verkaik, R.; Giacco, A.; Senese, R.; Arp, P.P.; Jhamai, P.M.; Pagnotta, S.M.; Broer, L.; et al. Short-Term, Combined Fasting and Exercise Improves Body Composition in Healthy Males. Int. J. Sport. Nutr. Exerc. Metab. 2020, 30, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Sbrignadello, S.; Göbl, C.; Tura, A. Bioelectrical Impedance Analysis for the Assessment of Body Composition in Sarcopenia and Type 2 Diabetes. Nutrients 2022, 14, 1864. [Google Scholar] [CrossRef]
- Pranoto, A.; Rejeki, P.S.; Miftahussurur, M.; Setiawan, H.K.; Yosika, G.F.; Munir, M.; Maesaroh, S.; Purwoto, S.P.; Waritsu, C.; Yamaoka, Y. Single 30 min treadmill exercise session suppresses the production of pro-inflammatory cytokines and oxidative stress in obese female adolescents. J. Basic. Clin. Physiol. Pharmacol. 2023, 34, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Min, K.; Jang, J.; Kim, S.C.; Kang, M.S.; Jang, S.J.; Lee, J.Y.; Kim, S.H.; Kim, M.K.; An, S.A.; et al. Involvement of Immune Responses in the Efficacy of Cord Blood Cell Therapy for Cerebral Palsy. Stem Cells Dev. 2015, 24, 2259–2268. [Google Scholar] [CrossRef]
- Zhang, S.; Li, Y.; Li, T.; Zhang, Y.; Li, H.; Cheng, Z.; Peng, N.; Liu, Y.; Xu, J.; He, H. Activable Targeted Protein Degradation Platform Based on Light-triggered Singlet Oxygen. J. Med. Chem. 2022, 65, 3632–3643. [Google Scholar] [CrossRef]
- Longo, V.D.; Mattson, M.P. Fasting: Molecular mechanisms and clinical applications. Cell Metab. 2014, 19, 181–192. [Google Scholar] [CrossRef]
- Yin, W.; Sun, L.; Liang, Y.; Luo, C.; Feng, T.; Zhang, Y.; Zhang, W.; Yin, Y. Maternal intermittent fasting deteriorates offspring metabolism via suppression of hepatic mTORC1 signaling. FASEB J. 2023, 37, e22831. [Google Scholar] [CrossRef] [PubMed]
- Vendelbo, M.H.; Møller, A.B.; Christensen, B.; Nellemann, B.; Clasen, B.F.; Nair, K.S.; Jørgensen, J.O.; Jessen, N.; Møller, N. Fasting increases human skeletal muscle net phenylalanine release and this is associated with decreased mTOR signaling. PLoS ONE 2014, 9, e102031. [Google Scholar] [CrossRef] [PubMed]
- Dethlefsen, C.; Lillelund, C.; Midtgaard, J.; Andersen, C.; Pedersen, B.K.; Christensen, J.F.; Hojman, P. Exercise regulates breast cancer cell viability: Systemic training adaptations versus acute exercise responses. Breast Cancer Res. Treat. 2016, 159, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Liang, J.; Wu, L.; Zhang, H.; Lv, J.; Chen, N. Exercise-Induced Autophagy Suppresses Sarcopenia Through Akt/mTOR and Akt/FoxO3a Signal Pathways and AMPK-Mediated Mitochondrial Quality Control. Front. Physiol. 2020, 11, 583478. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, L.; Wu, W.; Liu, Z.; Huang, Y.; Yang, L.; Luo, Q.; Chen, J.; Hou, Y.; Song, G. Exercise protects proliferative muscle satellite cells against exhaustion via the Igfbp7-Akt-mTOR axis. Theranostics 2020, 10, 6448–6466. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.D.C.; Morais, G.P.; Ropelle, E.R.; de Moura, L.P.; Cintra, D.E.; Pauli, J.R.; de Freitas, E.C.; Rorato, R.; da Silva, A.S.R. Using Intermittent Fasting as a Non-pharmacological Strategy to Alleviate Obesity-Induced Hypothalamic Molecular Pathway Disruption. Front. Nutr. 2022, 9, 858320. [Google Scholar] [CrossRef] [PubMed]
- Jaspers, R.T.; Zillikens, M.C.; Friesema, E.C.; delli Paoli, G.; Bloch, W.; Uitterlinden, A.G.; Goglia, F.; Lanni, A.; de Lange, P. Exercise, fasting, and mimetics: Toward beneficial combinations? FASEB J. 2017, 31, 14–28. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Xu, Y.; Liu, F. Hypothalamic roles of mTOR complex I: Integration of nutrient and hormone signals to regulate energy homeostasis. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E994–E1002. [Google Scholar] [CrossRef] [PubMed]
- Tsubai, T.; Noda, Y.; Ito, K.; Nakao, M.; Seino, Y.; Oiso, Y.; Hamada, Y. Insulin elevates leptin secretion and mRNA levels via cyclic AMP in 3T3-L1 adipocytes deprived of glucose. Heliyon 2016, 2, e00194. [Google Scholar] [CrossRef][Green Version]
- Wang, B.; Cheng, K.K.-Y. Hypothalamic AMPK as a Mediator of Hormonal Regulation of Energy Balance. Int. J. Mol. Sci. 2018, 19, 3552. [Google Scholar] [CrossRef]
- Evans, M.C.; Lord, R.A.; Anderson, G.M. Multiple Leptin Signalling Pathways in the Control of Metabolism and Fertility: A Means to Different Ends? Int. J. Mol. Sci. 2021, 22, 9210. [Google Scholar] [CrossRef] [PubMed]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [PubMed]
- Pena-Leon, V.; Perez-Lois, R.; Seoane, L.M. mTOR Pathway is Involved in Energy Homeostasis Regulation as a Part of the Gut–Brain Axis. Int. J. Mol. Sci. 2020, 21, 5715. [Google Scholar] [CrossRef] [PubMed]
- Szwed, A.; Kim, E.; Jacinto, E. Regulation and metabolic functions of mTORC1 and mTORC2. Physiol. Rev. 2021, 101, 1371–1426. [Google Scholar] [CrossRef] [PubMed]
- Noureldein, M.H.; Eid, A.A. Gut microbiota and mTOR signaling: Insight on a new pathophysiological interaction. Microb. Pathog. 2018, 118, 98–104. [Google Scholar] [CrossRef]
- Ye, Y.; Liu, H.; Zhang, F.; Hu, F. mTOR signaling in Brown and Beige adipocytes: Implications for thermogenesis and obesity. Nutr. Metab. 2019, 16, 74. [Google Scholar] [CrossRef] [PubMed]
- Widiatmaja, D.M.; Lutvyani, A.; Sari, D.R.; Kurniasari, H.; Meiliana, I.D.; Fasitasari, M.; Yamaoka, Y.; Rejeki, P.S. The effect of long-term ketogenic diet on serum adiponectin and insulin-like growth factor-1 levels in mice. J. Basic Clin. Physiol. Pharmacol. 2021, 33, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Fulco, M.; Sartorelli, V. Comparing and contrasting the roles of AMPK and SIRT1 in metabolic tissues. Cell Cycle 2008, 7, 3669–3679. [Google Scholar] [CrossRef] [PubMed]
- Jamshed, H.; Beyl, R.A.; Della Manna, D.L.; Yang, E.S.; Ravussin, E.; Peterson, C.M. Early Time-Restricted Feeding Improves 24-Hour Glucose Levels and Affects Markers of the Circadian Clock, Aging, and Autophagy in Humans. Nutrients 2019, 11, 1234. [Google Scholar] [CrossRef]
- Huynh, M.K.; Kinyua, A.W.; Yang, D.J.; Kim, K.W. Hypothalamic AMPK as a Regulator of Energy Homeostasis. Neural Plast. 2016, 2016, 2754078. [Google Scholar] [CrossRef]
- Green, C.L.; Lamming, D.W.; Fontana, L. Molecular mechanisms of dietary restriction promoting health and longevity. Nat. Rev. Mol. Cell Biol. 2022, 23, 56–73. [Google Scholar] [CrossRef] [PubMed]
- Mani, K.; Javaheri, A.; Diwan, A. Lysosomes Mediate Benefits of Intermittent Fasting in Cardiometabolic Disease: The Janitor Is the Undercover Boss. Compr. Physiol. 2018, 8, 1639–1667. [Google Scholar] [PubMed]
- Hu, D.; Xie, Z.; Ye, Y.; Bahijri, S.; Chen, M. The beneficial effects of intermittent fasting: An update on mechanism, and the role of circadian rhythm and gut microbiota. Hepatobil. Surg. Nutr. 2020, 9, 597–602. [Google Scholar]
- Gnoni, M.; Beas, R.; Vásquez-Garagatti, R. Is there any role of intermittent fasting in the prevention and improving clinical outcomes of COVID-19? Intersection between inflammation, mTOR pathway, autophagy and calorie restriction. Virusdisease 2021, 32, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Hong, N.; Kim, K.-W.; Cho, S.J.; Lee, M.; Lee, Y.-H.; Lee, Y.-H.; Kang, E.S.; Cha, B.-S.; Lee, B.-W. The Effectiveness of Intermittent Fasting to Reduce Body Mass Index and Glucose Metabolism: A Systematic Review and Meta-Analysis. J. Clin. Med. 2019, 8, 1645. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.; Alkhawam, H.; Madanieh, R.; Shah, N.; Kosmas, C.E.; Vittorio, T.J. Aerobic vs anaerobic exercise training effects on the cardiovascular system. World J. Cardiol. 2017, 9, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Flack, K.D.; Ufholz, K.; Johnson, L.; Fitzgerald, J.S.; Roemmich, J.N. Energy compensation in response to aerobic exercise training in overweight adults. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R619–R626. [Google Scholar] [CrossRef] [PubMed]
- Muscella, A.; Stefàno, E.; Lunetti, P.; Capobianco, L.; Marsigliante, S. The Regulation of Fat Metabolism during Aerobic Exercise. Biomolecules 2020, 10, 1699. [Google Scholar] [CrossRef]
- Spaulding, H.R.; Yan, Z. AMPK and the Adaptation to Exercise. Annu. Rev. Physiol. 2022, 84, 209–227. [Google Scholar] [CrossRef]
- Said, M.A.; Abdelmoneem, M.; Almaqhawi, A.; Hamid Kotob, A.A.; Alibrahim, M.C.; Bougmiza, I. Multidisciplinary approach to obesity: Aerobic or resistance physical exercise? J. Exerc. Sci. Fit. 2018, 16, 118–123. [Google Scholar] [CrossRef]
- Marandi, S.M.; Abadi, N.G.; Esfarjani, F.; Mojtahedi, H.; Ghasemi, G. Effects of intensity of aerobics on body composition and blood lipid profile in obese/overweight females. Int. J. Prev. Med. 2013, 4, S118–S125. [Google Scholar] [PubMed]
- Al Saif, A.; Alsenany, S. Aerobic and anaerobic exercise training in obese adults. J. Phys. Ther. Sci. 2015, 27, 1697–1700. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.H.; Ko, M.C.; Wu, L.S.; Yeh, D.P.; Kan, N.W.; Lee, P.F.; Hsieh, J.W.; Tseng, C.Y.; Ho, C.C. Benefits of different intensity of aerobic exercise in modulating body composition among obese young adults: A pilot randomized controlled trial. Health Qual. Life Outcomes 2017, 15, 168. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.H.; Lee, J.K. Effect of Different Intensities of Aerobic Exercise Combined with Resistance Exercise on Body Fat, Lipid Profiles, and Adipokines in Middle-Aged Women with Obesity. Int. J. Environ. Res. Public Health 2023, 20, 3991. [Google Scholar] [CrossRef] [PubMed]
- Jafari, A.; Pourrazi, H.; Nikookheslat, S.; Baradaran, B. Effect of Exercise Training on Bcl-2 and Bax Gene Expression in the Rat Heart. Gene Cell Tissue 2015, 2, e60174. [Google Scholar] [CrossRef]
- Roberts, C.J.; Campbell, I.C.; Troop, N. Increases in weight during chronic stress are partially associated with a switch in food choice towards increased carbohydrate and saturated fat intake. Eur. Eat. Disord. Rev. 2014, 22, 77–82. [Google Scholar] [CrossRef] [PubMed]
- López-Domínguez, J.A.; Khraiwesh, H.; González-Reyes, J.A.; López-Lluch, G.; Navas, P.; Ramsey, J.J.; de Cabo, R.; Burón, M.I.; Villalba, J.M. Dietary fat and aging modulate apoptotic signaling in liver of calorie-restricted mice. J. Gerontol. A Biol. Sci. Med. Sci. 2015, 70, 399–409. [Google Scholar] [CrossRef]
- Katare, R.G.; Kakinuma, Y.; Arikawa, M.; Yamasaki, F.; Sato, T. Chronic intermittent fasting improves the survival following large myocardial ischemia by activation of BDNF/VEGF/PI3K signaling pathway. J. Mol. Cell Cardiol. 2009, 46, 405–412. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef]
- Shamas-Din, A.; Kale, J.; Leber, B.; Andrews, D.W. Mechanisms of action of Bcl-2 family proteins. Cold Spring Harb. Perspect. Biol. 2013, 5, a008714. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, I.; Bodo, J.; Hill, B.T.; Hsi, E.D.; Almasan, A. Targeting BCL-2 in B-cell malignancies and overcoming therapeutic resistance. Cell Death Dis. 2020, 11, 941. [Google Scholar] [CrossRef]
- Campbell, K.L.; McTiernan, A.; Li, S.S.; Sorensen, B.E.; Yasui, Y.; Lampe, J.W.; King, I.B.; Ulrich, C.M.; Rudolph, R.E.; Irwin, M.L.; et al. Effect of a 12-month exercise intervention on the apoptotic regulating proteins Bax and Bcl-2 in colon crypts: A randomized controlled trial. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1767–1774. [Google Scholar] [CrossRef] [PubMed]
- Rami, M.; Rahdar, S.; Ahmadi Hekmatikar, A.; Awang Daud, D.M. Highlighting the novel effects of high-intensity interval training on some histopathological and molecular indices in the heart of type 2 diabetic rats. Front. Endocrinol. 2023, 14, 1175585. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.-W.; Yoo, S.-Z.; No, M.-H.; Park, D.-H.; Kang, J.-H.; Kim, T.-W.; Kim, C.-J.; Seo, D.-Y.; Han, J.; Yoon, J.-H.; et al. Exercise Training Attenuates Obesity-Induced Skeletal Muscle Remodeling and Mitochondria-Mediated Apoptosis in the Skeletal Muscle. Int. J. Environ. Res. Public Health 2018, 15, 2301. [Google Scholar] [CrossRef]
- Podhorska-Okolow, M.; Sandri, M.; Zampieri, S.; Brun, B.; Rossini, K.; Carraro, U. Apoptosis of myofibres and satellite cells: Exercise-induced damage in skeletal muscle of the mouse. Neuropathol. Appl. Neurobiol. 1998, 24, 518–531. [Google Scholar] [CrossRef]
- Cory, S.; Huang, D.C.; Adams, J.M. The Bcl-2 family: Roles in cell survival and oncogenesis. Oncogene 2003, 22, 8590–8607. [Google Scholar] [CrossRef]
- Evangelisti, C.; Cenni, V.; Lattanzi, G. Potential therapeutic effects of the MTOR inhibitors for preventing ageing and progeria-related disorders. Br. J. Clin. Pharmacol. 2016, 82, 1229–1244. [Google Scholar] [CrossRef]
- Sunjaya, A.P.; Sunjaya, A.F. Targeting ageing and preventing organ degeneration with metformin. Diabetes Metab. 2021, 47, 101203. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameters | (CG; n = 10) | (IFG; n = 10) | (EXG; n = 10) | (IFEXG; n = 10) | p-Value |
|---|---|---|---|---|---|
| Age (yrs) | 23.90 ± 0.99 | 24.20 ± 2.69 | 23.30 ± 1.57 | 22.90 ± 1.59 | 0.390 † |
| SBP (mmHg) | 111.45 ± 5.86 | 116.65 ± 12.37 | 119.95 ± 15.65 | 116.20 ± 3.43 | 0.360 † |
| DBP (mmHg) | 67.68 ± 3.24 | 73.95 ± 8.55 | 75.30 ± 8.44 | 74.50 ± 7.13 | 0.086 † |
| HR (bpm) | 76.80 ± 7.87 | 81.50 ± 5.72 | 79.30 ± 10.94 | 75.60 ± 4.79 | 0.336 † |
| SpO2 (%) | 98.00 ± 1.05 | 98.80 ± 0.42 | 97.80 ± 1.23 | 98.40 ± 0.69 | 0.085 † |
| BT (°C) | 35.96 ± 0.83 | 35.87 ± 0.58 | 35.58 ± 0.80 | 36.22 ± 0.67 | 0.285 † |
| FBG (mg/dL) | 95.60 ± 2.50 | 94.90 ± 2.96 | 94.60 ± 2.01 | 95.60 ± 2.67 | 0.761 † |
| Hb (g/dL) | 14.27 ± 2.02 | 13.81 ± 1.29 | 13.78 ± 1.50 | 14.04 ± 1.63 | 0.899 † |
| Height (m) | 1.55 ± 0.03 | 1.57 ± 0.05 | 1.57 ± 0.06 | 1.55 ± 0.04 | 0.566 † |
| Weight (kg) | 69.54 ± 6.10 | 75.22 ± 13.04 | 70.10 ± 9.88 | 76.26 ± 10.21 | 0.337 † |
| BMI (kg/m2) | 29.19 ± 2.49 | 31.22 ± 5.01 | 28.36 ± 2.87 | 31.57 ± 3.61 | 0.157 † |
| FAT (%) | 44.24 ± 2.66 | 45.84 ± 4.15 | 43.56 ± 3.54 | 47.21 ± 3.82 | 0.119 † |
| FM (kg) | 31.72 ± 3.68 | 34.93 ± 9.11 | 30.78 ± 6.52 | 36.30 ± 7.49 | 0.260 † |
| FFM (kg) | 38.65 ± 2.30 | 40.29 ± 4.31 | 39.32 ± 3.78 | 39.96 ± 3.10 | 0.725 † |
| MM (kg) | 36.44 ± 2.09 | 37.94 ± 3.93 | 37.05 ± 3.47 | 37.62 ± 2.83 | 0.727 † |
| TBW (%) | 28.42 ± 2.52 | 30.43 ± 5.68 | 28.83 ± 3.49 | 29.73 ± 3.14 | 0.659 † |
| TBW (kg) | 41.03 ± 3.75 | 40.63 ± 2.53 | 41.27 ± 2.53 | 39.18 ± 2.96 | 0.411 † |
| BM (kg) | 2.21 ± 0.21 | 2.35 ± 0.39 | 2.27 ± 0.32 | 2.34 ± 0.28 | 0.708 † |
| BMR (kcal) | 1268.30 ± 80.96 | 1336.20 ± 168.73 | 1285.10 ± 146.22 | 1338.80 ± 125.68 | 0.553 † |
| MA (yrs) | 48.70 ± 0.95 | 49.10 ± 2.88 | 48.90 ± 2.73 | 48.10 ± 2.18 | 0.790 † |
| Assessment | (CG; n = 10) | (IFG; n = 10) | (EXG; n = 10) | (IFEXG; n = 10) | p-Value |
|---|---|---|---|---|---|
| Pre-mTOR (ng/mL) | 2.15 ± 1.22 | 2.23 ± 1.22 | 2.12 ± 0.85 | 2.09 ± 0.83 | 0.992 † |
| Post-mTOR (ng/mL) | 2.07 ± 0.68 | 1.42 ± 0.83 * | 1.36 ± 0.64 * | 0.82 ± 0.53 * | 0.003 † |
| ∆-mTOR (ng/mL) | −0.08 ± 1.33 | −0.81 ± 0.97 | −0.76 ± 0.59 | −1.26 ± 0.79 * | 0.047 † |
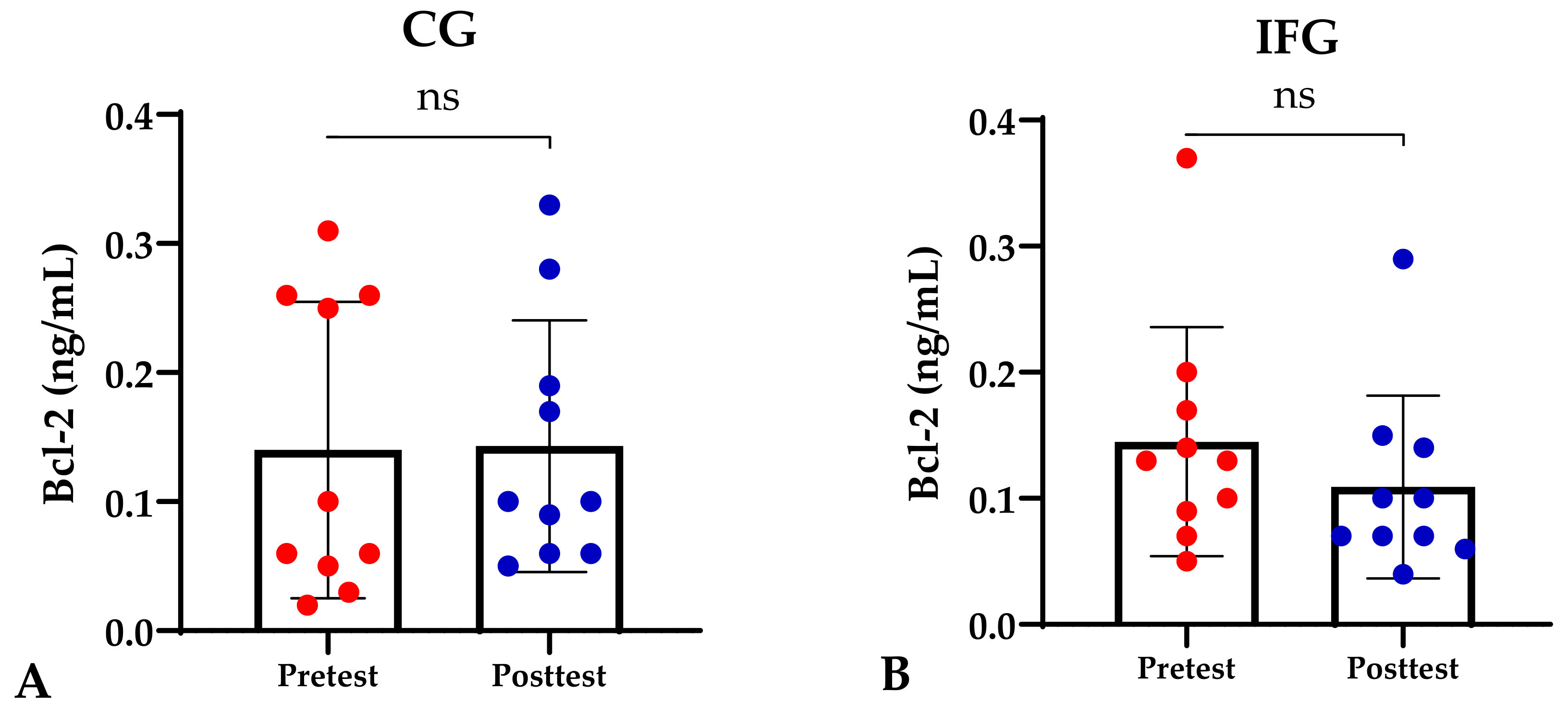
| Pre-Bcl-2 (ng/mL) | 0.14 ± 0.12 | 0.15 ± 0.10 | 0.14 ± 0.11 | 0.12 ± 0.08 | 0.932 ≠ |
| Post-Bcl-2 (ng/mL) | 0.14 ± 0.16 | 0.11 ± 0.07 | 0.11 ± 0.07 | 0.05 ± 0.04 *^$ | 0.015 ≠ |
| ∆-Bcl-2 (ng/mL) | 0.01 ± 0.17 | −0.04 ± 0.14 | −0.04 ± 0.15 | −0.07 ± 0.09 | 0.676 ≠ |
| ∆-Weight (kg) | 0.75 ± 0.87 | −0.59 ± 1.13 * | −0.78 ± 0.58 * | −1.10 ± 0.42 * | 0.001 † |
| ∆-BMI (kg/m2) | 0.71 ± 1.25 | −0.25 ± 0.47 | −0.32 ± 0.24 | −0.46 ± 0.19 | 0.002 † |
| ∆-FAT (%) | 0.58 ± 0.50 | −0.30 ± 0.71 | −0.96 ± 1.21 | −1.31 ± 0.38 *^ | 0.001 † |
| ∆-FM (kg) | 0.42 ± 0.43 | −0.49 ± 0.88 * | −0.70 ± 1.03 * | −1.16 ± 0.21 *^ | 0.001 † |
| ∆-MM (kg) | −0.14 ± 0.88 | 0.39 ± 0.55 | 0.59 ± 0.69 * | 1.15 ± 0.33 *^ | 0.001 † |
| ∆-MA (yrs) | 0.20 ± 0.63 | −0.60 ± 1.27 | −0.70 ± 1.57 | −1.40 ± 0.69 * | 0.026 † |
| Parameters | ∆-mTOR (ng/mL) | ∆-Bcl-2 (ng/mL) | ||
|---|---|---|---|---|
| r | p-Value | r | p-Value | |
| ∆-Weight (kg) | 0.315 * | 0.048 | 0.397 * | 0.023 |
| ∆-BMI (kg/m2) | 0.537 ** | p ≤ 0.001 | 0.331 * | 0.037 |
| ∆-FAT (%) | 0.316 * | 0.047 | 0.369 * | 0.044 |
| ∆-FM (kg) | 0.358 * | 0.023 | 0.367 * | 0.035 |
| ∆-MM (kg) | −0.418 ** | p ≤ 0.001 | −0.501 ** | p ≤ 0.001 |
| ∆-MA (yrs) | 0.500 ** | p ≤ 0.001 | 0.736** | p ≤ 0.001 |
| Parameters | ∆-MA (yrs) | |
|---|---|---|
| r | p-Value | |
| ∆-FAT (%) | 0.523 ** | p ≤ 0.001 |
| ∆-FM (kg) | 0.514 ** | p ≤ 0.001 |
| ∆-MM (kg) | −0.529 ** | p ≤ 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rejeki, P.S.; Pranoto, A.; Widiatmaja, D.M.; Utami, D.M.; Izzatunnisa, N.; Sugiharto; Lesmana, R.; Halim, S. Combined Aerobic Exercise with Intermittent Fasting Is Effective for Reducing mTOR and Bcl-2 Levels in Obese Females. Sports 2024, 12, 116. https://doi.org/10.3390/sports12050116
Rejeki PS, Pranoto A, Widiatmaja DM, Utami DM, Izzatunnisa N, Sugiharto, Lesmana R, Halim S. Combined Aerobic Exercise with Intermittent Fasting Is Effective for Reducing mTOR and Bcl-2 Levels in Obese Females. Sports. 2024; 12(5):116. https://doi.org/10.3390/sports12050116
Chicago/Turabian StyleRejeki, Purwo Sri, Adi Pranoto, Deandra Maharani Widiatmaja, Dita Mega Utami, Nabilah Izzatunnisa, Sugiharto, Ronny Lesmana, and Shariff Halim. 2024. "Combined Aerobic Exercise with Intermittent Fasting Is Effective for Reducing mTOR and Bcl-2 Levels in Obese Females" Sports 12, no. 5: 116. https://doi.org/10.3390/sports12050116
APA StyleRejeki, P. S., Pranoto, A., Widiatmaja, D. M., Utami, D. M., Izzatunnisa, N., Sugiharto, Lesmana, R., & Halim, S. (2024). Combined Aerobic Exercise with Intermittent Fasting Is Effective for Reducing mTOR and Bcl-2 Levels in Obese Females. Sports, 12(5), 116. https://doi.org/10.3390/sports12050116