

Agreement between Ventilatory Thresholds and Bilaterally Measured Vastus Lateralis Muscle Oxygen Saturation Breakpoints in Trained Cyclists: Effects of Age and Performance


Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Design
2.3. Procedures and Measurements
2.4. Computations and Measurements
2.5. Statistical Analysis
3. Results
3.1. Performance Characteristics
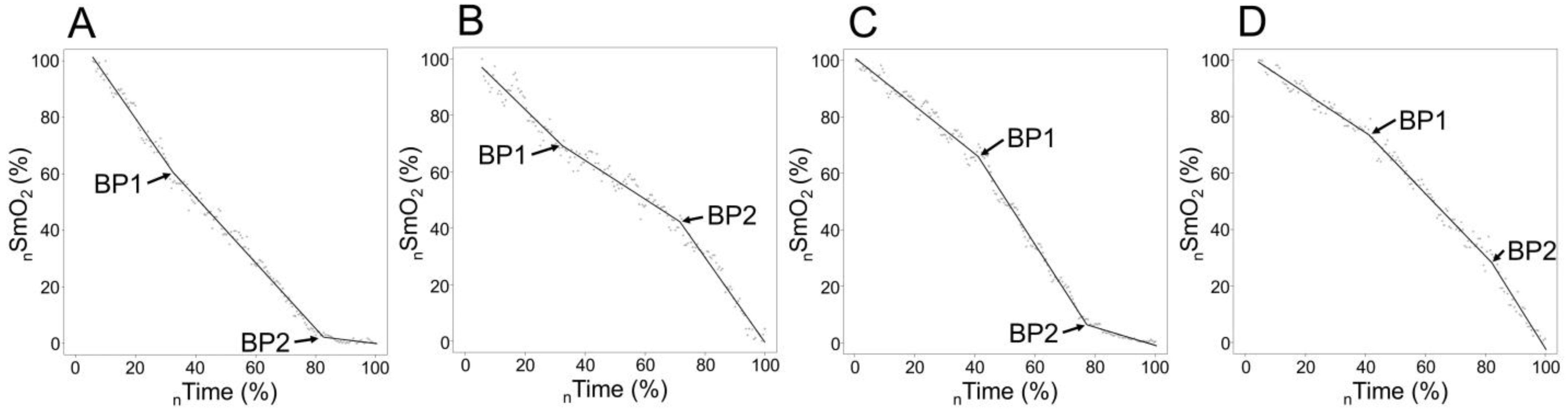
3.2. Typology and Breakpoints of SmO2 signal sloping according to Age Group
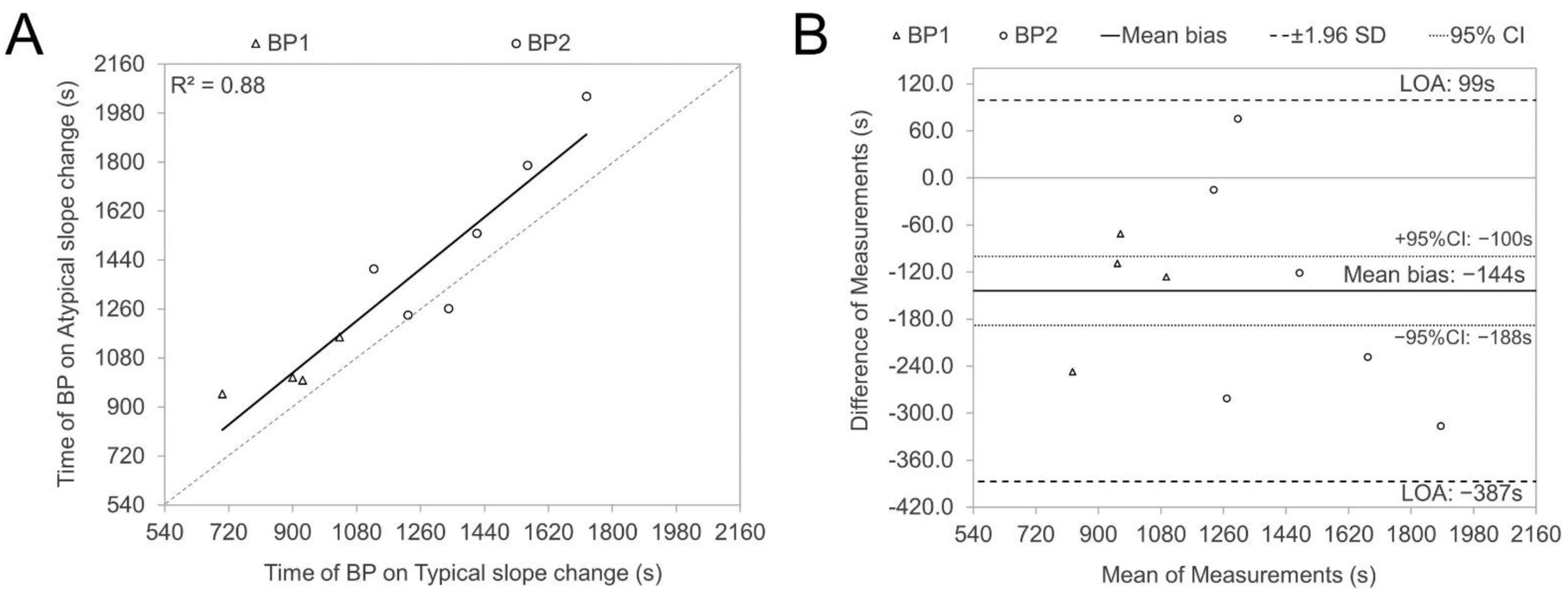
3.3. Agreement between Ventilatory Thresholds and Bilaterally Measured SmO2 Breakpoints and between ND- and DO-Side SmO2 Breakpoints among Different Age Groups
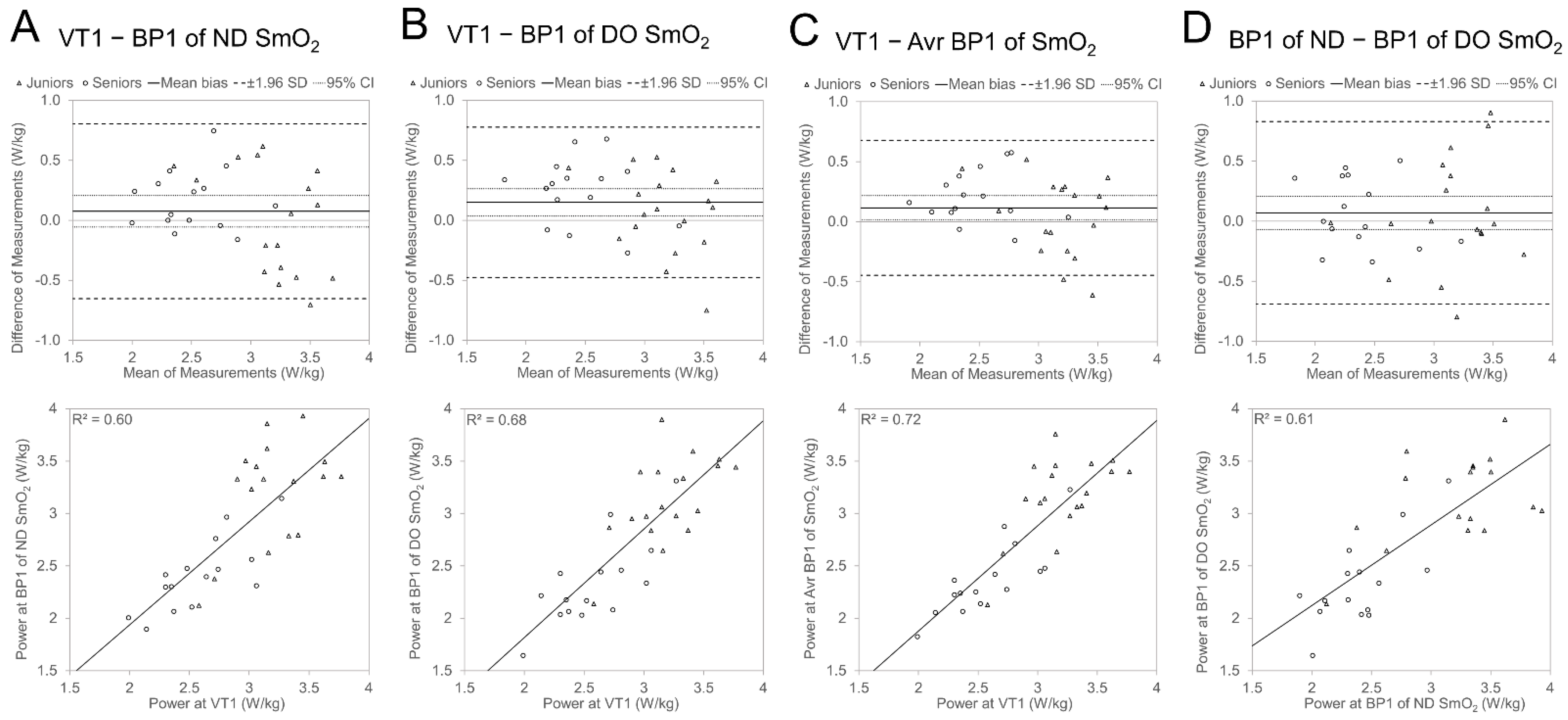
3.3.1. Agreement between VT1 and BP1
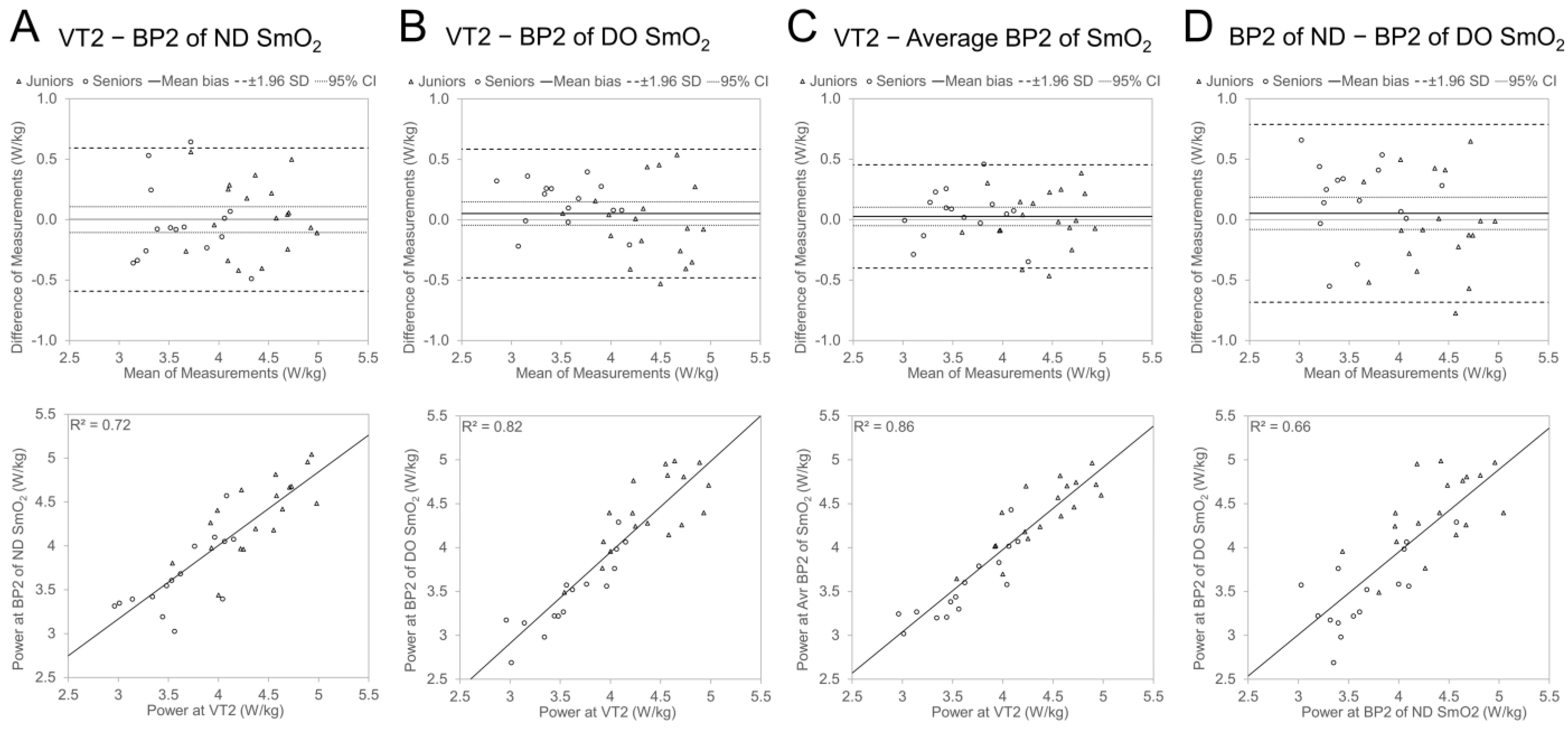
3.3.2. Agreement between VT2 and BP2
3.4. Effect of Cyclist’s Performance Characteristics on Differences between Ventilatory Thresholds and SmO2 Breakpoints
4. Discussion
4.1. Breakpoint Evaluation and Signal Sloping
4.2. Agreement between VT and SmO2 BP and Effect of Age Group
4.3. The Effect of Cycling Performance Measures to Agreement between VT and SmO2 BP
4.4. Future Research and Practical Implications
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seiler, S. What is best practice for training intensity and duration distribution in endurance athletes? Int. J. Sports Physiol. Perform. 2010, 5, 276–291. [Google Scholar] [CrossRef] [PubMed]
- West, J.B. A century of pulmonary gas exchange. Am. J. Respir. Crit. Care Med. 2004, 169, 897–902. [Google Scholar] [CrossRef] [PubMed]
- Perrey, S.; Ferrari, M. Muscle Oximetry in Sports Science: A Systematic Review. Sports Med. 2018, 48, 597–616. [Google Scholar] [CrossRef] [PubMed]
- Perrey, S. Muscle oxygenation unlocks the secrets of physiological responses to exercise: Time to exploit it in the training monitoring. Front. Sports Act. Living 2022, 4, 864825. [Google Scholar] [CrossRef]
- Joyner, M.J.; Coyle, E.F. Endurance exercise performance: The physiology of champions. J. Physiol. 2008, 586, 35–44. [Google Scholar] [CrossRef] [PubMed]
- van Der Zwaard, S.; Brocherie, F.; Jaspers, R.T. Under the hood: Skeletal muscle determinants of endurance performance. Front. Sports Act. Living 2021, 3, 719434. [Google Scholar] [CrossRef]
- Bassett, D.R.; Howley, E.T. Limiting factors for maximum oxygen uptake and determinants of endurance performance. Med. Sci. Sports Exerc. 2000, 32, 70–84. [Google Scholar] [CrossRef]
- Amann, M. Central and peripheral fatigue: Interaction during cycling exercise in humans. Med. Sci. Sports Exerc. 2011, 43, 2039–2045. [Google Scholar] [CrossRef]
- Feldner, H.A.; Howell, D.; Kelly, V.E.; McCoy, S.W.; Steele, K.M. “Look, your muscles are firing!”: A qualitative study of clinician perspectives on the use of surface electromyography in neurorehabilitation. Arch. Phys. Med. Rehabil. 2019, 100, 663–675. [Google Scholar] [CrossRef]
- Barstow, T.J. Understanding near infrared spectroscopy and its application to skeletal muscle research. J. Appl. Physiol. 2019, 126, 1360–1376. [Google Scholar] [CrossRef]
- Crum, E.M.; O’Connor, W.J.; Van Loo, L.; Valckx, M.; Stannard, S.R. Validity and reliability of the Moxy oxygen monitor during incremental cycling exercise. Eur. J. Sport. Sci. 2017, 17, 1037–1043. [Google Scholar] [CrossRef]
- Yogev, A.; Arnold, J.; Clarke, D.; Guenette, J.A.; Sporer, B.C.; Koehle, M.S. Comparing the Respiratory Compensation Point With Muscle Oxygen Saturation in Locomotor and Non-locomotor Muscles Using Wearable NIRS Spectroscopy During Whole-Body Exercise. Front. Physiol. 2022, 13, 818733. [Google Scholar] [CrossRef]
- van der Zwaard, S.; Jaspers, R.T.; Blokland, I.J.; Achterberg, C.; Visser, J.M.; den Uil, A.R.; Hofmijster, M.J.; Levels, K.; Noordhof, D.A.; de Haan, A.; et al. Oxygenation Threshold Derived from Near-Infrared Spectroscopy: Reliability and Its Relationship with the First Ventilatory Threshold. PLoS ONE 2016, 11, e0162914. [Google Scholar] [CrossRef]
- Murias, J.M.; Spencer, M.D.; Keir, D.A.; Paterson, D.H. Systemic and vastus lateralis muscle blood flow and O2 extraction during ramp incremental cycle exercise. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R720–R725. [Google Scholar] [CrossRef]
- Iannetta, D.; Qahtani, A.; Maturana, F.M.; Murias, J.M. The near-infrared spectroscopy-derived deoxygenated haemoglobin breaking-point is a repeatable measure that demarcates exercise intensity domains. J. Sci. Med. Sport 2017, 20, 873–877. [Google Scholar] [CrossRef]
- Keir, D.A.; Fontana, F.Y.; Robertson, T.C.; Murias, J.M.; Paterson, D.H.; Kowalchuk, J.M.; Pogliaghi, S. Exercise intensity thresholds: Identifying the boundaries of sustainable performance. Med. Sci. Sports Exerc. 2015, 47, 1932–1940. [Google Scholar] [CrossRef]
- Feldmann, A.; Ammann, L.; Gachter, F.; Zibung, M.; Erlacher, D. Muscle Oxygen Saturation Breakpoints Reflect Ventilatory Thresholds in Both Cycling and Running. J. Hum. Kinet. 2022, 83, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Briceño, F.; Espinosa-Ramirez, M.; Keim-Bagnara, V.; Carreño-Román, M.; Rodríguez-Villagra, R.; Villegas-Belmar, F.; Viscor, G.; Gabrielli, L.; Andía, M.E.; Araneda, O.F. Determination of the respiratory compensation point by detecting changes in intercostal muscles oxygenation by using Near-Infrared Spectroscopy. Life 2022, 12, 444. [Google Scholar] [CrossRef] [PubMed]
- Bhambhani, Y.N. Muscle oxygenation trends during dynamic exercise measured by near infrared spectroscopy. Can. J. Appl. Physiol. 2004, 29, 504–523. [Google Scholar] [CrossRef] [PubMed]
- Farzam, P.; Starkweather, Z.; Franceschini, M.A. Validation of a novel wearable, wireless technology to estimate oxygen levels and lactate threshold power in the exercising muscle. Physiol. Rep. 2018, 6, e13664. [Google Scholar] [CrossRef] [PubMed]
- Spencer, M.D.; Murias, J.M.; Lamb, H.P.; Kowalchuk, J.M.; Paterson, D.H. Are the parameters of VO2, heart rate and muscle deoxygenation kinetics affected by serial moderate-intensity exercise transitions in a single day? Eur. J. Appl. Physiol. 2011, 111, 591–600. [Google Scholar] [CrossRef]
- Alejo, L.B.; Montalvo-Pérez, A.; Valenzuela, P.L.; Revuelta, C.; Ozcoidi, L.M.; de la Calle, V.; Mateo-March, M.; Lucia, A.; Santalla, A.; Barranco-Gil, D. Comparative analysis of endurance, strength and body composition indicators in professional, under-23 and junior cyclists. Front. Physiol. 2022, 13, 945552. [Google Scholar] [CrossRef]
- Meredith, C.; Frontera, W.; Fisher, E.; Hughes, V.; Herland, J.; Edwards, J.; Evans, W. Peripheral effects of endurance training in young and old subjects. J. Appl. Physiol. 1989, 66, 2844–2849. [Google Scholar] [CrossRef]
- Letnes, J.M.; Dalen, H.; Aspenes, S.T.; Salvesen, Ø.; Wisløff, U.; Nes, B.M. Age-related change in peak oxygen uptake and change of cardiovascular risk factors. The HUNT Study. Prog. Cardiovasc. Dis. 2020, 63, 730–737. [Google Scholar] [CrossRef]
- Grassi, B.; Quaresima, V. Near-infrared spectroscopy and skeletal muscle oxidative function in vivo in health and disease: A review from an exercise physiology perspective. J. Biomed. Opt. 2016, 21, 091313. [Google Scholar] [CrossRef] [PubMed]
- Bottinelli, R.; Reggiani, C. Human skeletal muscle fibres: Molecular and functional diversity. Prog. Biophys. Mol. Biol. 2000, 73, 195–262. [Google Scholar] [CrossRef]
- Padilla, S.; Mujika, I.; Cuesta, G.; Goiriena, J.J. Level ground and uphill cycling ability in professional road cycling. Med. Sci. Sports Exerc. 1999, 31, 878–885. [Google Scholar] [CrossRef] [PubMed]
- Koga, S.; Poole, D.C.; Ferreira, L.F.; Whipp, B.J.; Kondo, N.; Saitoh, T.; Ohmae, E.; Barstow, T.J. Spatial heterogeneity of quadriceps muscle deoxygenation kinetics during cycle exercise. J. Appl. Physiol. 2007, 103, 2049–2056. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, T.; Ferreira, L.F.; Barstow, T.J.; Poole, D.C.; Ooue, A.; Kondo, N.; Koga, S. Effects of prior heavy exercise on heterogeneity of muscle deoxygenation kinetics during subsequent heavy exercise. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, R615–R621. [Google Scholar] [CrossRef] [PubMed]
- Carpes, F.P.; Mota, C.B.; Faria, I.E. On the bilateral asymmetry during running and cycling—A review considering leg preference. Phys. Ther. Sport 2010, 11, 136–142. [Google Scholar] [CrossRef]
- Smak, W.; Neptune, R.; Hull, M. The influence of pedaling rate on bilateral asymmetry in cycling. J. Biomech. 1999, 32, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Reinpõld, K.; Rannama, I. Oxygen Uptake and Bilaterally Measured Vastus Lateralis Muscle Oxygen Desaturation Kinetics in Well-Trained Endurance Cyclists. J. Funct. Morphol. Kinesiol. 2023, 8, 64. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, A.; Schmitz, R.; Erlacher, D. Near-infrared spectroscopy-derived muscle oxygen saturation on a 0% to 100% scale: Reliability and validity of the Moxy Monitor. J. Biomed. Opt. 2019, 24, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sendra-Pérez, C.; Sanchez-Jimenez, J.L.; Marzano-Felisatti, J.M.; Encarnación-Martínez, A.; Salvador-Palmer, R.; Priego-Quesada, J.I. Reliability of threshold determination using portable muscle oxygenation monitors during exercise testing: A systematic review and meta-analysis. Sci. Rep. 2023, 13, 12649. [Google Scholar] [CrossRef] [PubMed]
- De Pauw, K.; Roelands, B.; Cheung, S.S.; de Geus, B.; Rietjens, G.; Meeusen, R. Guidelines to classify subject groups in sport-science research. Int. J. Sports Physiol. Perform. 2013, 8, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Geraskin, D.; Boeth, H.; Kohl-Bareis, M. Optical measurement of adipose tissue thickness and comparison with ultrasound, magnetic resonance imging, and callipers. J. Biomed. Opt. 2009, 14, 044017. [Google Scholar] [CrossRef] [PubMed]
- Rodger, S.M.; Plews, D.J.; McQuillan, J.; Driller, M.W. Evaluation of the Cyclus cycle ergometer and the Stages power meter for measurement of power output in cycling. J. Sci. Cycl. 2016, 5, 16–22. [Google Scholar]
- Wasserman, K.; Hansen, J.E.; Sue, D.Y.; Stringer, W.W.; Whipp, B.J. Principles of exercise testing and interpretation: Including pathophysiology and clinical applications. Med. Sci. Sports Exerc. 2005, 37, 1249. [Google Scholar]
- Muggeo, V.M. Segmented: An R package to fit regression models with broken-line relationships. R News 2008, 8, 20–25. [Google Scholar]
- Hopkins, W.; Marshall, S.; Batterham, A.; Hanin, J. Progressive statistics for studies in sports medicine and exercise science. Med. Sci. Sports Exerc. 2009, 41, 3. [Google Scholar] [CrossRef]
- Rodrigo-Carranza, V.; González-Mohíno, F.; Turner, A.P.; Rodriguez-Barbero, S.; González-Ravé, J.M. Using a portable near-infrared spectroscopy device to estimate the second ventilatory threshold. Int. J. Sports Med. 2021, 42, 905–910. [Google Scholar] [CrossRef]
- Ducrocq, G.P.; Kaufman, M.P. Inorganic phosphate and lactate potentiate the pressor response to acidic stimuli in rats. Exp. Physiol. 2020, 105, 613–621. [Google Scholar] [CrossRef]
- Grotle, A.-K.; Macefield, V.G.; Farquhar, W.B.; O’Leary, D.S.; Stone, A.J. Recent advances in exercise pressor reflex function in health and disease. Auton. Neurosci. 2020, 228, 102698. [Google Scholar] [CrossRef] [PubMed]
- Faude, O.; Kindermann, W.; Meyer, T. Lactate threshold concepts: How valid are they? Sports Med. 2009, 39, 469–490. [Google Scholar] [CrossRef]
- Kindermann, W.; Simon, G.; Keul, J. The significance of the aerobic-anaerobic transition for the determination of work load intensities during endurance training. Eur. J. Appl. Physiol. Occup. Physiol. 1979, 42, 25–34. [Google Scholar] [CrossRef]
- Radegran, G. Ultrasound Doppler estimates of femoral artery blood flow during dynamic knee extensor exercise in humans. J. Appl. Physiol. 1997, 83, 1383–1388. [Google Scholar] [CrossRef]
- Batterson, P.M.; Kirby, B.S.; Hasselmann, G.; Feldmann, A. Muscle oxygen saturation rates coincide with lactate-based exercise thresholds. Eur. J. Appl. Physiol. 2023, 123, 2249–2258. [Google Scholar] [CrossRef] [PubMed]
- Zoladz, J.A.; Gladden, L.B.; Hogan, M.C.; Nieckarz, Z.; Grassi, B. Progressive recruitment of muscle fibers is not necessary for the slow component of VO2 kinetics. J. Appl. Physiol. 2008, 105, 575–580. [Google Scholar] [CrossRef]
- Poole, D.C.; Rossiter, H.B.; Brooks, G.A.; Gladden, L.B. The anaerobic threshold: 50+ years of controversy. J. Physiol. 2021, 599, 737–767. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Grassi, B.; Christensen, P.M.; Krustrup, P.; Bangsbo, J.; Poole, D.C. Slow component of VO2 kinetics: Mechanistic bases and practical applications. Med. Sci. Sports Exerc. 2011, 43, 2046–2062. [Google Scholar] [CrossRef]
- Broker, J.P.; Gregor, R.J. Mechanical energy management in cycling: Source relations and energy expenditure. Med. Sci. Sports Exerc. 1994, 26, 64–74. [Google Scholar] [CrossRef] [PubMed]
- So, R.C.; Ng, J.K.-F.; Ng, G.Y. Muscle recruitment pattern in cycling: A review. Phys. Ther. Sport 2005, 6, 89–96. [Google Scholar] [CrossRef]
- Baum, B.S.; Li, L. Lower extremity muscle activities during cycling are influenced by load and frequency. J. Electromyogr. Kinesiol. 2003, 13, 181–190. [Google Scholar] [CrossRef]
- Elmer, S.; Barratt, P.; Korff, T.; Martin, J. Joint-specific power production during submaximal and maximal cycling. In Proceedings of the ISBS-Conference Proceedings Archive, Marquette, MI, USA, 19–23 July 2010. [Google Scholar]
- Turpin, N.A.; Costes, A.; Moretto, P.; Watier, B. Can muscle coordination explain the advantage of using the standing position during intense cycling? J. Sci. Med. Sport 2017, 20, 611–616. [Google Scholar] [CrossRef]
- Hug, F.; Bendahan, D.; Le Fur, Y.; Cozzone, P.J.; Grelot, L. Heterogeneity of muscle recruitment pattern during pedaling in professional road cyclists: A magnetic resonance imaging and electromyography study. Eur. J. Appl. Physiol. 2004, 92, 334–342. [Google Scholar] [CrossRef]
- Hug, F.; Drouet, J.M.; Champoux, Y.; Couturier, A.; Dorel, S. Interindividual variability of electromyographic patterns and pedal force profiles in trained cyclists. Eur. J. Appl. Physiol. 2008, 104, 667–678. [Google Scholar] [CrossRef]
- Hug, F.; Turpin, N.A.; Guével, A.; Dorel, S. Is interindividual variability of EMG patterns in trained cyclists related to different muscle synergies? J. Appl. Physiol. 2010, 108, 1727–1736. [Google Scholar] [CrossRef]
- Chapman, A.R.; Vicenzino, B.; Blanch, P.; Hodges, P.W. Patterns of leg muscle recruitment vary between novice and highly trained cyclists. J. Electromyogr. Kinesiol. 2008, 18, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Priego, J.I.; Bini, R.R.; Lanferdini, F.; Carpes, F.P. Effects of workload level on muscle recruitment in cycling. Hum. Mov. 2014, 15, 45–50. [Google Scholar] [CrossRef][Green Version]
- Dingwell, J.B.; Joubert, J.E.; Diefenthaeler, F.; Trinity, J.D. Changes in muscle activity and kinematics of highly trained cyclists during fatigue. IEEE Trans. Biomed. Eng. 2008, 55, 2666–2674. [Google Scholar] [CrossRef] [PubMed]
- Hawley, J.A.; Stepto, N.K. Adaptations to training in endurance cyclists: Implications for performance. Sports Med. 2001, 31, 511–520. [Google Scholar] [CrossRef]
- Coyle, E.; Feltner, M.; Kautz, S.; Hamilton, M.; Montain, S.; Baylor, A.; Abraham, L.; Petrek, G. Physiological and biomechanical factors associated with elite endurance cycling performance. Med. Sci. Sports Exerc. 1991, 23, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Mezzani, A.; Grassi, B.; Giordano, A.; Corrà, U.; Colombo, S.; Giannuzzi, P. Age-related prolongation of phase I of VO2 on-kinetics in healthy humans. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2010, 299, R968–R976. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Raleigh, C.; Donne, B.; Fleming, N. Association between different Non-Invasively Derived Thresholds with Lactate Threshold during graded incremental exercise. Int. J. Exerc. Sci. 2018, 11, 391. [Google Scholar] [PubMed]
- Salas-Montoro, J.-A.; Mateo-March, M.; Sánchez-Muñoz, C.; Zabala, M. Determination of second lactate threshold using near-infrared spectroscopy in elite cyclists. Int. J. Sports Med. 2022, 43, 721–728. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Juniors | Seniors | ||||||
|---|---|---|---|---|---|---|---|---|
| N | Mean | SD | CoV | N | Mean | SD | CoV | |
| Age (y) | 18 | 18.2 | 1.6 | 0.09 | 15 | 43.8 | 5.2 | 0.12 * |
| Height (m) | 18 | 1.847 | 0.052 | 0.03 | 15 | 1.825 | 0.052 | 0.03 |
| Body mass (kg) | 18 | 73.2 | 7.0 | 0.10 | 15 | 81.5 | 6.7 | 0.08 * |
| BMI (body mass index) | 18 | 21.5 | 2.0 | 0.09 | 15 | 24.4 | 1.6 | 0.07 * |
| ATT (adipose tissue thickness) (mm) | 18 | 3.9 | 1.3 | 0.34 | 15 | 6.7 | 1.7 | 0.26 * |
| Cycling stasis (years) | 18 | 5.3 | 1.3 | 0.24 | 15 | 18.1 | 6.1 | 0.34 * |
| Cycling distance during past season (km) | 18 | 17,061 | 3607 | 0.21 | 15 | 10,273 | 2964 | 0.29 * |
| Variable | Group | N | Absolute Values | Relative Values (*/kg) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | CoV | p | Cohen’s d | Mean | SD | CoV | p | Cohen’s d | |||
| P@VT1 (W) | Juniors | 18 | 235.0 | 36.1 | 0.15 | 0.032 | 0.79 | 3.20 | 0.31 | 0.10 | <0.001 | 1.86 |
| Seniors | 15 | 209.4 | 27.3 | 0.13 | 2.58 | 0.36 | 0.14 | |||||
| P@VT2 (W) | Juniors | 18 | 321.1 | 40.6 | 0.13 | 0.050 | 0.71 | 4.39 | 0.41 | 0.09 | <0.001 | 1.95 |
| Seniors | 15 | 293.7 | 36.0 | 0.12 | 3.61 | 0.39 | 0.11 | |||||
| PP (W) | Juniors | 18 | 389.3 | 41.9 | 0.11 | 0.039 | 0.75 | 5.33 | 0.37 | 0.07 | <0.001 | 2.23 |
| Seniors | 15 | 358.7 | 39.0 | 0.11 | 4.41 | 0.46 | 0.10 | |||||
| VO2@VT1 (mL/min) | Juniors | 18 | 3260 | 422 | 0.13 | 0.326 | 0.35 | 44.5 | 3.2 | 0.07 | <0.001 | 1.62 |
| Seniors | 15 | 3120 | 376 | 0.12 | 38.4 | 4.4 | 0.11 | |||||
| VO2@VT2 (mL/min) | Juniors | 18 | 4341 | 495 | 0.11 | 0.507 | 0.24 | 59.3 | 4.3 | 0.07 | <0.001 | 1.60 |
| Seniors | 15 | 4223 | 512 | 0.12 | 51.9 | 5.1 | 0.10 | |||||
| VO2max (mL/min) | Juniors | 18 | 4950 | 528 | 0.11 | 0.892 | 0.05 | 67.7 | 3.9 | 0.06 | <0.001 | 1.34 |
| Seniors | 15 | 4924 | 536 | 0.11 | 60.6 | 6.5 | 0.11 | |||||
| Pmax5s (W) | Juniors | 18 | 1236.5 | 146.0 | 0.12 | <0.001 | 1.40 | 16.93 | 1.66 | 0.10 | <0.001 | 2.51 |
| Seniors | 15 | 1011.5 | 177.1 | 0.18 | 12.42 | 1.96 | 0.16 | |||||
| Pmax30s (W) | Juniors | 18 | 912.9 | 117.9 | 0.13 | 0.01 | 0.96 | 12.48 | 1.10 | 0.09 | <0.001 | 2.46 |
| Seniors | 15 | 797.9 | 122.2 | 0.15 | 9.78 | 1.11 | 0.11 | |||||
| Phenotype (P5s/P@VT2 (a.u.)) | Juniors | 18 | 3.89 | 0.53 | 0.14 | 0.018 | 0.87 | |||||
| Seniors | 15 | 3.45 | 0.46 | 0.134 | ||||||||
| Variable | Leg | Group | N | Mean | SD | Min | Max | CoV | p | Cohen’s d |
|---|---|---|---|---|---|---|---|---|---|---|
| Min SmO2 (%) | ND | Juniors | 18 | 12.8 | 6.4 | 1.7 | 26.7 | 0.50 | 0.18 | 0.48 |
| Seniors | 15 | 9.8 | 5.8 | 2.0 | 23.0 | 0.59 | ||||
| DO | Juniors | 18 | 14.0 | 8.9 | 3.0 | 31.0 | 0.64 | 0.12 | 0.33 | |
| Seniors | 15 | 8.9 | 4.0 | 1.3 | 17.2 | 0.46 | ||||
| Max SmO2 (%) | ND | Juniors | 18 | 61.1 | 8.6 | 46.1 | 79.7 | 0.14 | 0.16 | 0.51 |
| Seniors | 15 | 57.2 | 6.0 | 50.6 | 72.9 | 0.11 | ||||
| DO | Juniors | 18 | 61.3 | 8.9 | 50.1 | 80.7 | 0.15 | 0.24 | 0.24 | |
| Seniors | 15 | 56.8 | 5.5 | 44.2 | 66.3 | 0.10 | ||||
| BP1 SmO2 (%) | ND | Juniors | 17 | 40.2 | 11.3 | 18.2 | 61.7 | 0.28 | 0.73 | −0.12 |
| Seniors | 15 | 41.5 | 10.4 | 24.5 | 61.3 | 0.25 | ||||
| DO | Juniors | 18 | 43.2 | 13.9 | 21.0 | 72.3 | 0.32 | 0.88 | −0.05 | |
| Seniors | 15 | 43.9 | 10.7 | 25.9 | 63.0 | 0.24 | ||||
| BP2 SmO2 (%) | ND | Juniors | 18 | 19.0 | 9.1 | 3.6 | 41.4 | 0.48 | 0.94 | 0.03 |
| Seniors | 15 | 18.7 | 9.4 | 5.5 | 34.8 | 0.50 | ||||
| DO | Juniors | 18 | 21.3 | 11.5 | 4.7 | 48.8 | 0.54 | 0.26 | 0.40 | |
| Seniors | 15 | 16.9 | 10.1 | 2.4 | 39.1 | 0.60 |
| Variable | Group | N | Mean | SD | CoV | p | Cohen’s d |
|---|---|---|---|---|---|---|---|
| Pkg@BP1 of ND SmO2 (W/kg) | Juniors | 17 | 3.20 | 0.50 | 0.16 | <0.001 | 1.82 |
| Seniors | 15 | 2.41 | 0.35 | 0.14 | |||
| Pkg@BP1 of DO SmO2 (W/kg) | Juniors | 18 | 3.13 | 0.41 | 0.13 | <0.001 | 1.92 |
| Seniors | 15 | 2.34 | 0.41 | 0.18 | |||
| Pkg@BP1 of Avr SmO2 (W/kg) | Juniors | 18 | 3.16 | 0.39 | 0.12 | <0.001 | 2.11 |
| Seniors | 15 | 2.38 | 0.35 | 0.15 | |||
| Pkg@BP2 of ND SmO2 (W/kg) | Juniors | 18 | 4.36 | 0.42 | 0.10 | <0.001 | 1.68 |
| Seniors | 15 | 3.65 | 0.42 | 0.12 | |||
| Pkg@BP2 of DO SmO2 (W/kg) | Juniors | 18 | 4.41 | 0.44 | 0.10 | <0.001 | 2.17 |
| Seniors | 15 | 3.47 | 0.43 | 0.13 | |||
| Pkg@BP2 of Avr SmO2 (W/kg) | Juniors | 18 | 4.39 | 0.38 | 0.09 | <0.001 | 2.13 |
| Seniors | 15 | 3.56 | 0.40 | 0.11 |
| VT1—BP1 Power Difference (W/kg) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Variable | Group | N | Mean | SD | Limits of Agreement | Cohen’s d | ||
| Lower | Upper | p | ||||||
| VT1—BP1 of ND SmO2 | Juniors | 17 | −0.01 | 0.45 | −0.88 | 0.87 | 0.195 | −0.47 |
| Seniors | 15 | 0.17 | 0.25 * | −0.32 | 0.65 | |||
| VT1—BP1 of DO SmO2 | Juniors | 18 | 0.07 | 0.34 | −0.60 | 0.74 | 0.127 | −0.55 |
| Seniors | 15 | 0.24 | 0.28 * | −0.30 | 0.79 | |||
| VT1—Avr BP1 of SmO2 | Juniors | 18 | 0.04 | 0.32 | −0.59 | 0.68 | 0.103 | −0.59 |
| Seniors | 15 | 0.21 | 0.22 * | −0.22 | 0.63 | |||
| VT2—BP2 Power Difference (W/kg) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Variable | Group | N | Mean | SD | Limits of Agreement | Cohen’s d | ||
| Lower | Upper | p | ||||||
| VT2—BP2 of ND SmO2 | Juniors | 18 | 0.03 | 0.30 | −0.55 | 0.62 | 0.503 | 0.24 |
| Seniors | 15 | −0.04 | 0.31 | −0.66 | 0.58 | |||
| VT2—BP2 of DO SmO2 | Juniors | 18 | −0.02 | 0.31 | −0.63 | 0.59 | 0.096 | −0.60 |
| Seniors | 15 | 0.14 | 0.19 * | −0.23 | 0.51 | |||
| VT2—Avr BP2 of SmO2 | Juniors | 18 | 0.01 | 0.23 | −0.45 | 0.46 | 0.58 | −0.20 |
| Seniors | 15 | 0.05 | 0.20 | −0.35 | 0.45 | |||
| Variable | VT1-Avr BP1 Power Difference (W/kg) | VT2-Avr BP2 Power Difference (W/kg) | ||||
|---|---|---|---|---|---|---|
| Juniors (n = 17) | Seniors (n = 15) | All # (n = 32) | Juniors (n = 18) | Seniors (n = 15) | All # (n = 33) | |
| Pkg@VT1 (W/kg) | 0.263 | 0.335 | 0.288 | 0.268 | −0.022 | 0.136 |
| Pkg@VT2 (W/kg) | 0.084 | 0.423 | 0.219 | 0.388 | 0.249 | 0.331 |
| PPkg (W/kg) | 0.09 | 0.366 | 0.208 | 0.184 | 0.103 | 0.143 |
| VO2kg@VT1 (mL/min/kg) | 0.079 | 0.291 | 0.172 | 0.062 | 0.18 | 0.117 |
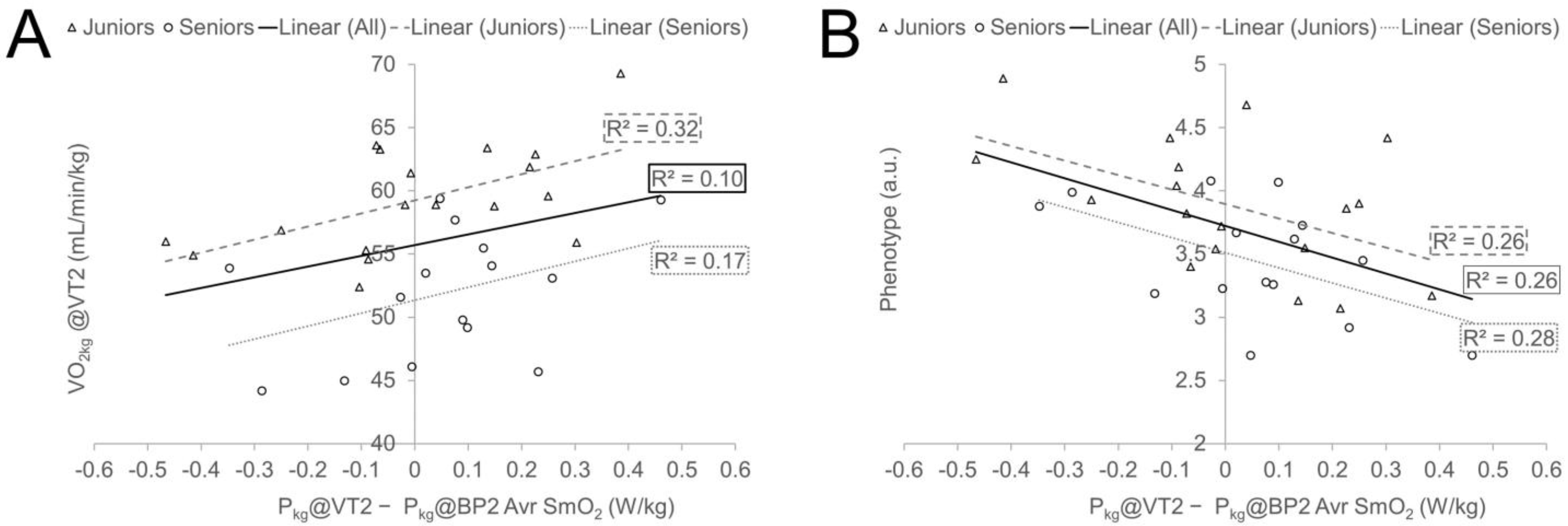
| VO2kg@VT2 (mL/min/kg) | 0.12 | 0.41 | 0.249 | 0.567 * | 0.409 | 0.488 ** |
| VO2kgmax (mL/min/kg) | 0.08 | 0.455 | 0.258 | 0.429 | 0.227 | 0.303 |
| Pkgmax5s (W/kg) | −0.293 | −0.238 | −0.264 | −0.342 | −0.295 | −0.317 |
| Pkgmax30s (W/kg) | −0.06 | 0.059 | −0.022 | −0.215 | 0.088 | −0.088 |
| Phenotype (P5s/P@VT2 (a.u.)) | −0.273 | −0.575 * | −0.385 * | −0.508 * | −0.526 * | −0.515 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reinpõld, K.; Rannama, I.; Port, K. Agreement between Ventilatory Thresholds and Bilaterally Measured Vastus Lateralis Muscle Oxygen Saturation Breakpoints in Trained Cyclists: Effects of Age and Performance. Sports 2024, 12, 40. https://doi.org/10.3390/sports12020040
Reinpõld K, Rannama I, Port K. Agreement between Ventilatory Thresholds and Bilaterally Measured Vastus Lateralis Muscle Oxygen Saturation Breakpoints in Trained Cyclists: Effects of Age and Performance. Sports. 2024; 12(2):40. https://doi.org/10.3390/sports12020040
Chicago/Turabian StyleReinpõld, Karmen, Indrek Rannama, and Kristjan Port. 2024. "Agreement between Ventilatory Thresholds and Bilaterally Measured Vastus Lateralis Muscle Oxygen Saturation Breakpoints in Trained Cyclists: Effects of Age and Performance" Sports 12, no. 2: 40. https://doi.org/10.3390/sports12020040
APA StyleReinpõld, K., Rannama, I., & Port, K. (2024). Agreement between Ventilatory Thresholds and Bilaterally Measured Vastus Lateralis Muscle Oxygen Saturation Breakpoints in Trained Cyclists: Effects of Age and Performance. Sports, 12(2), 40. https://doi.org/10.3390/sports12020040