Control of Scaphoideus titanus with Natural Products in Organic Vineyards

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Trials
2.2. Laboratory Bioassay
2.3. Statistical Analyses
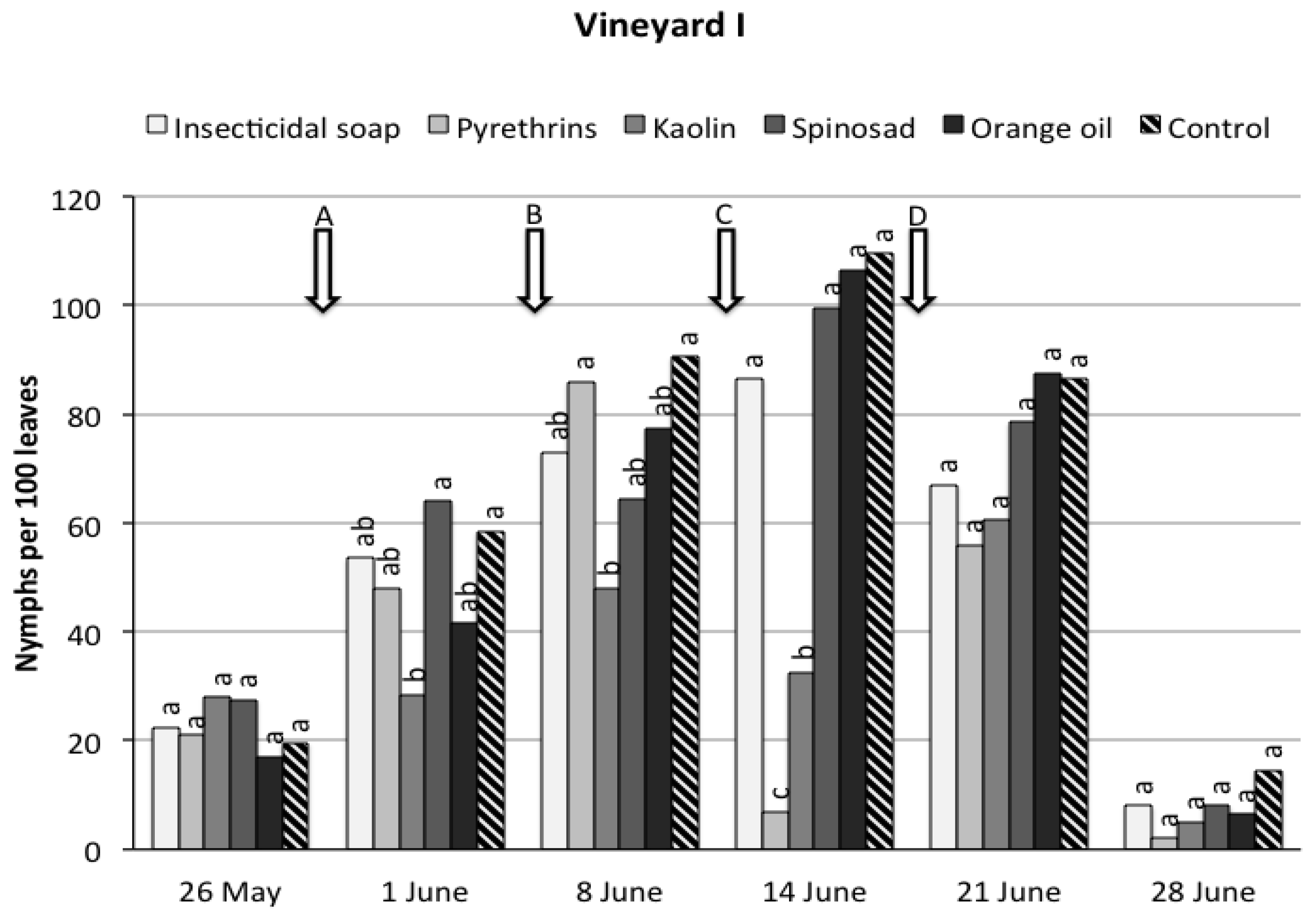
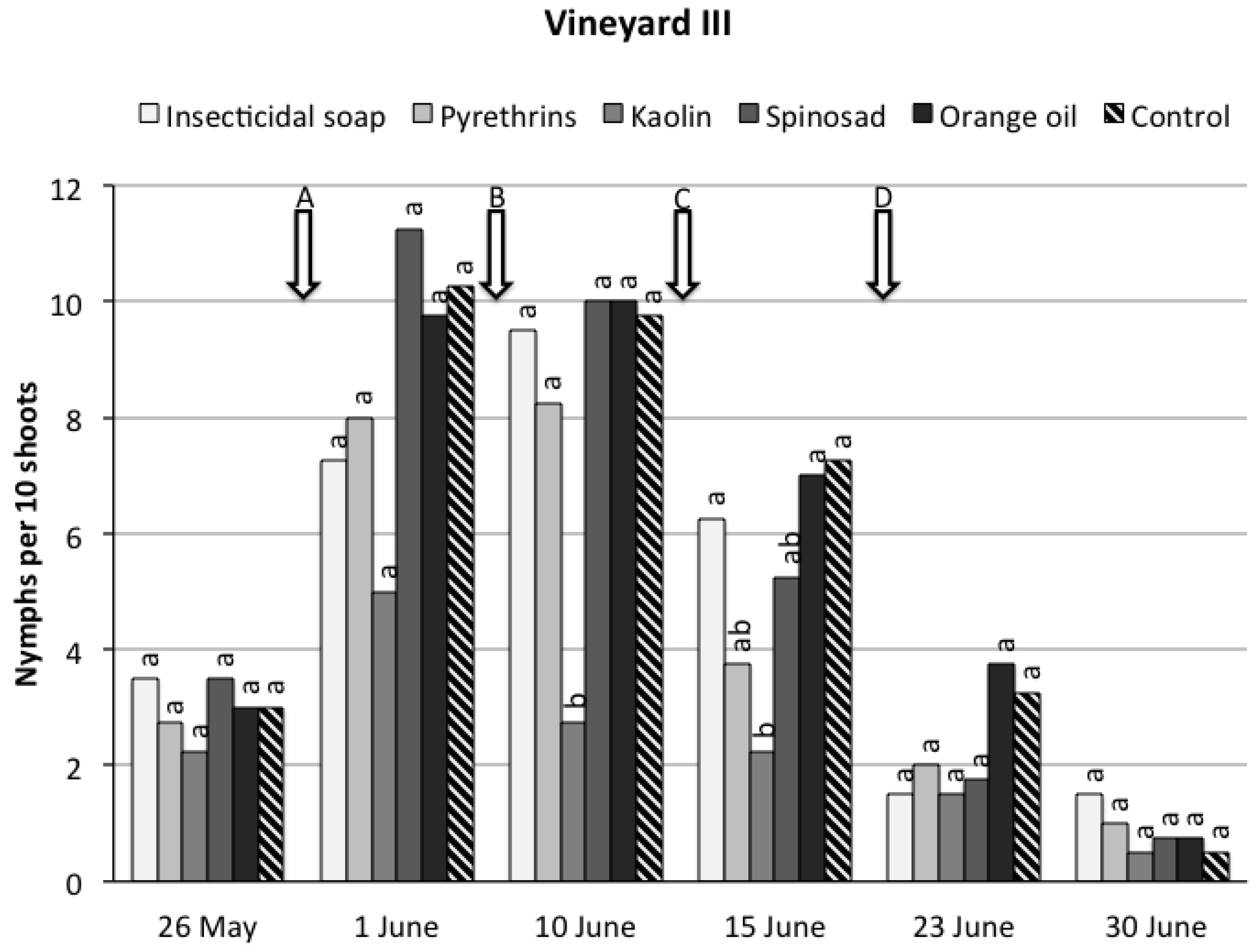
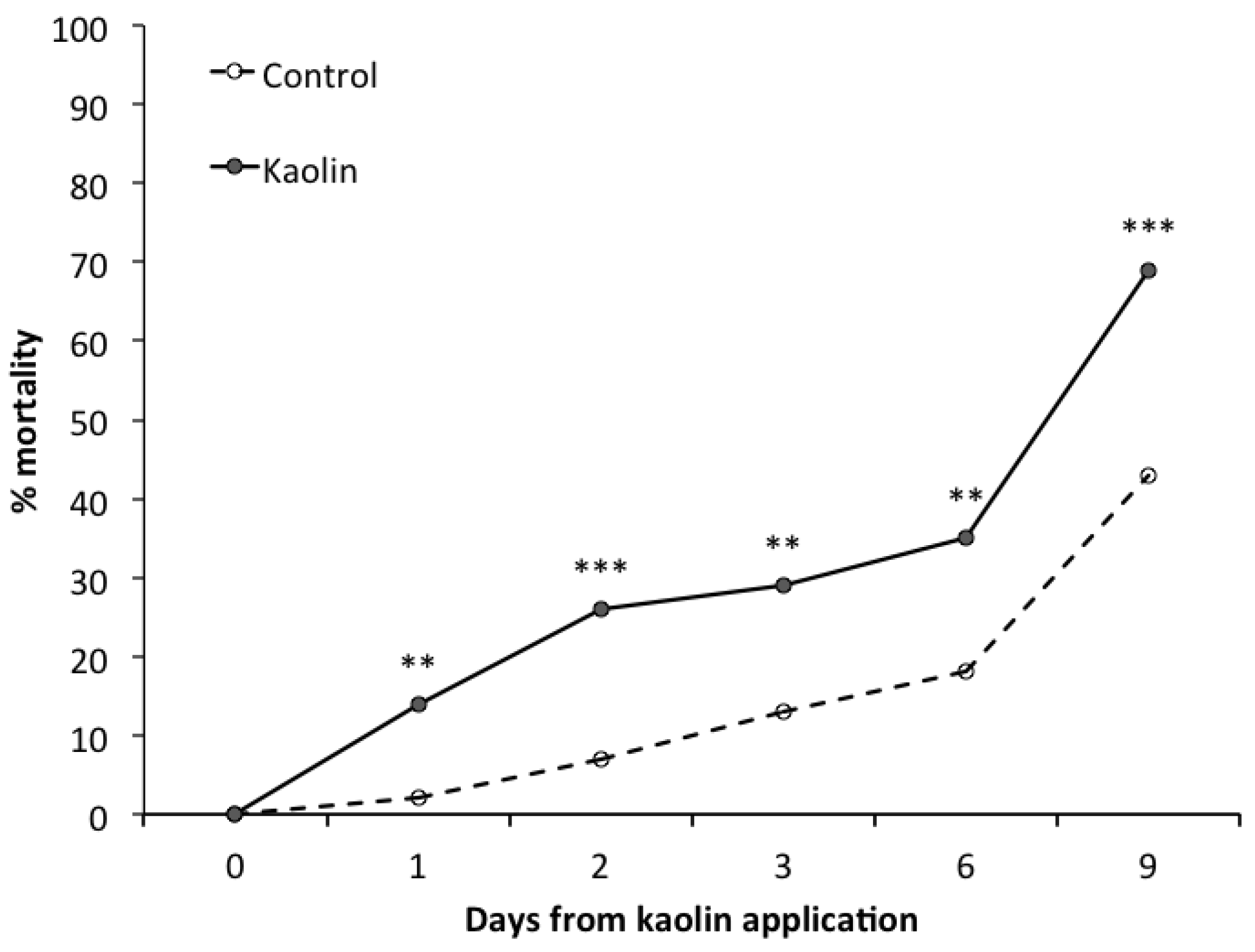
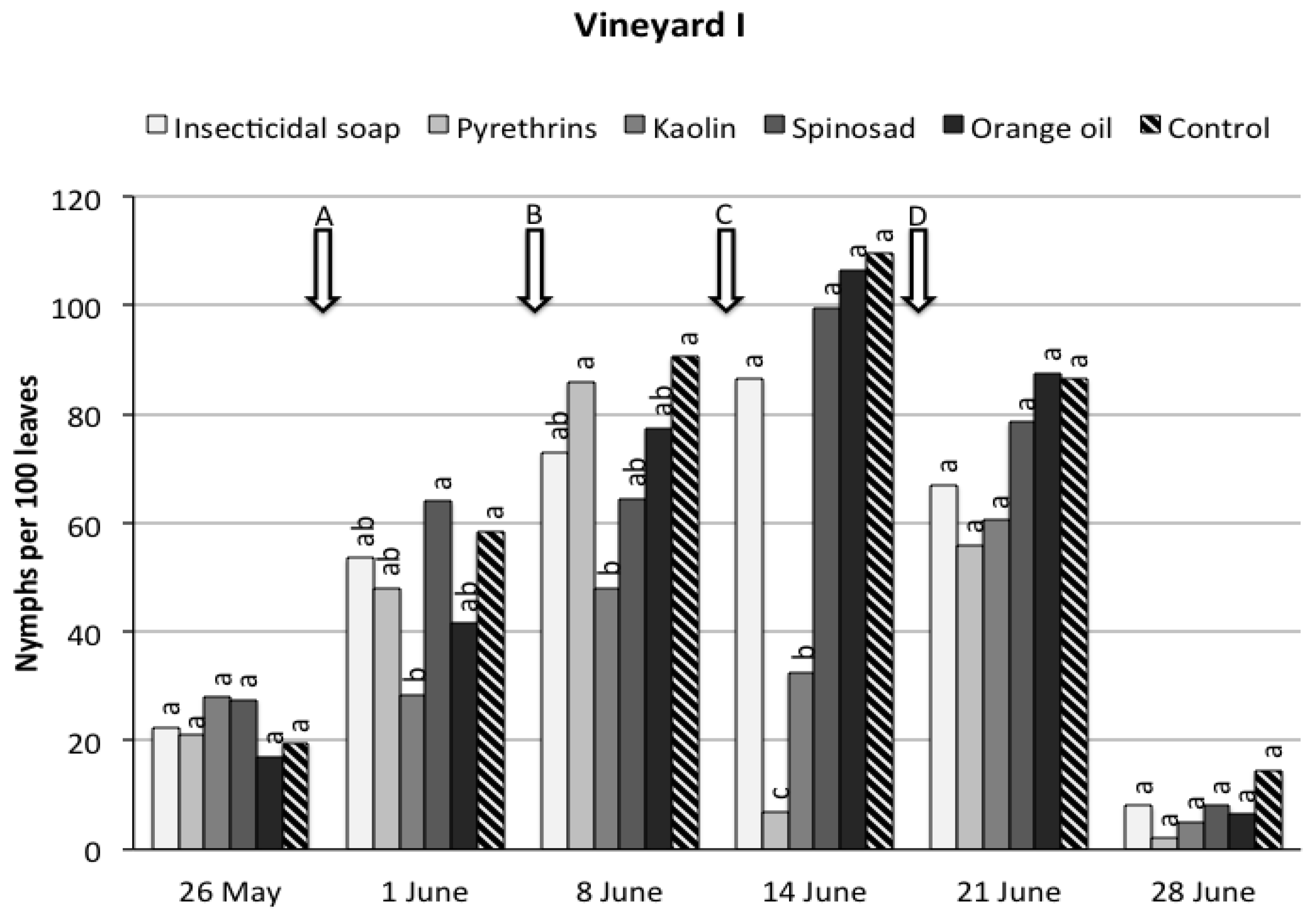
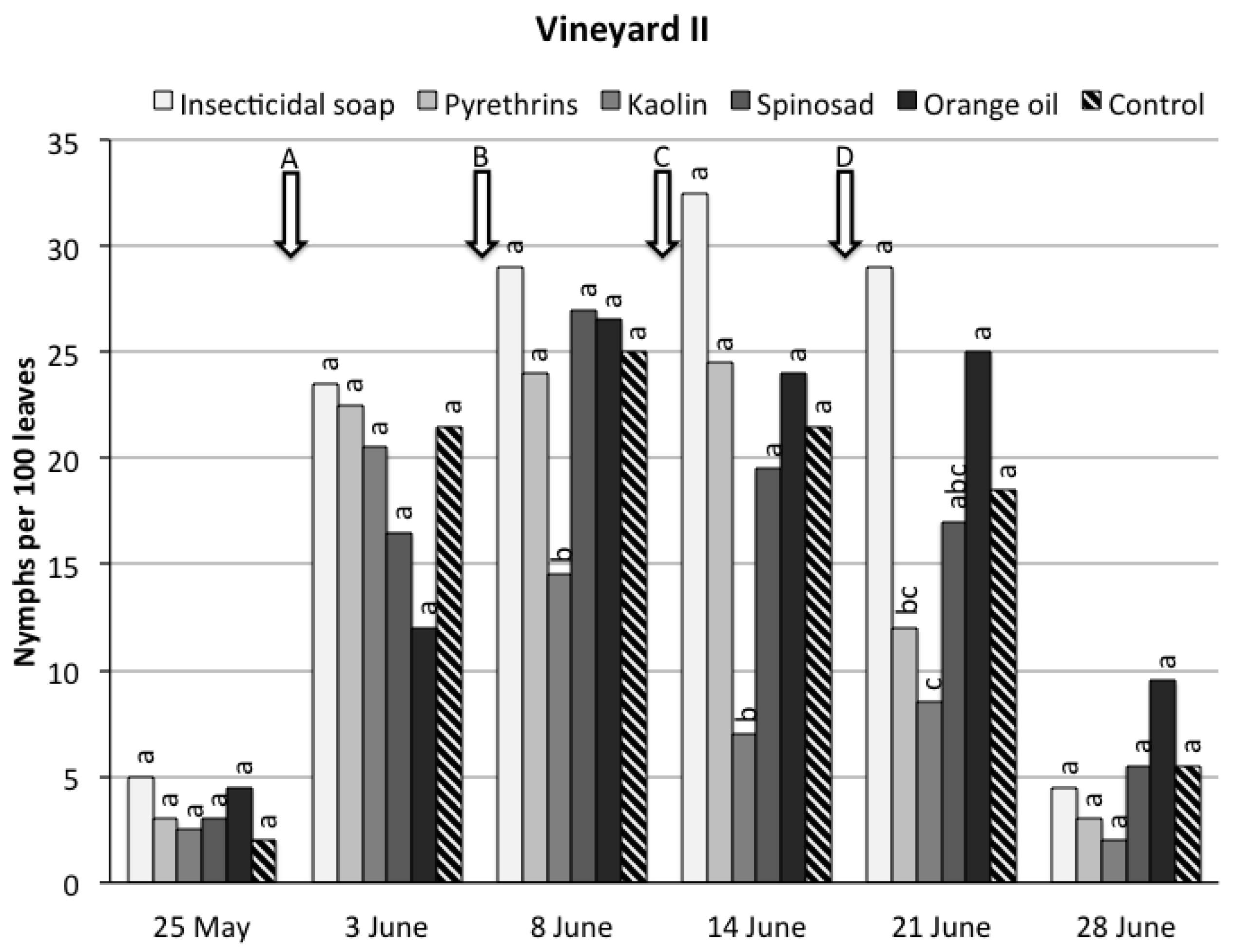
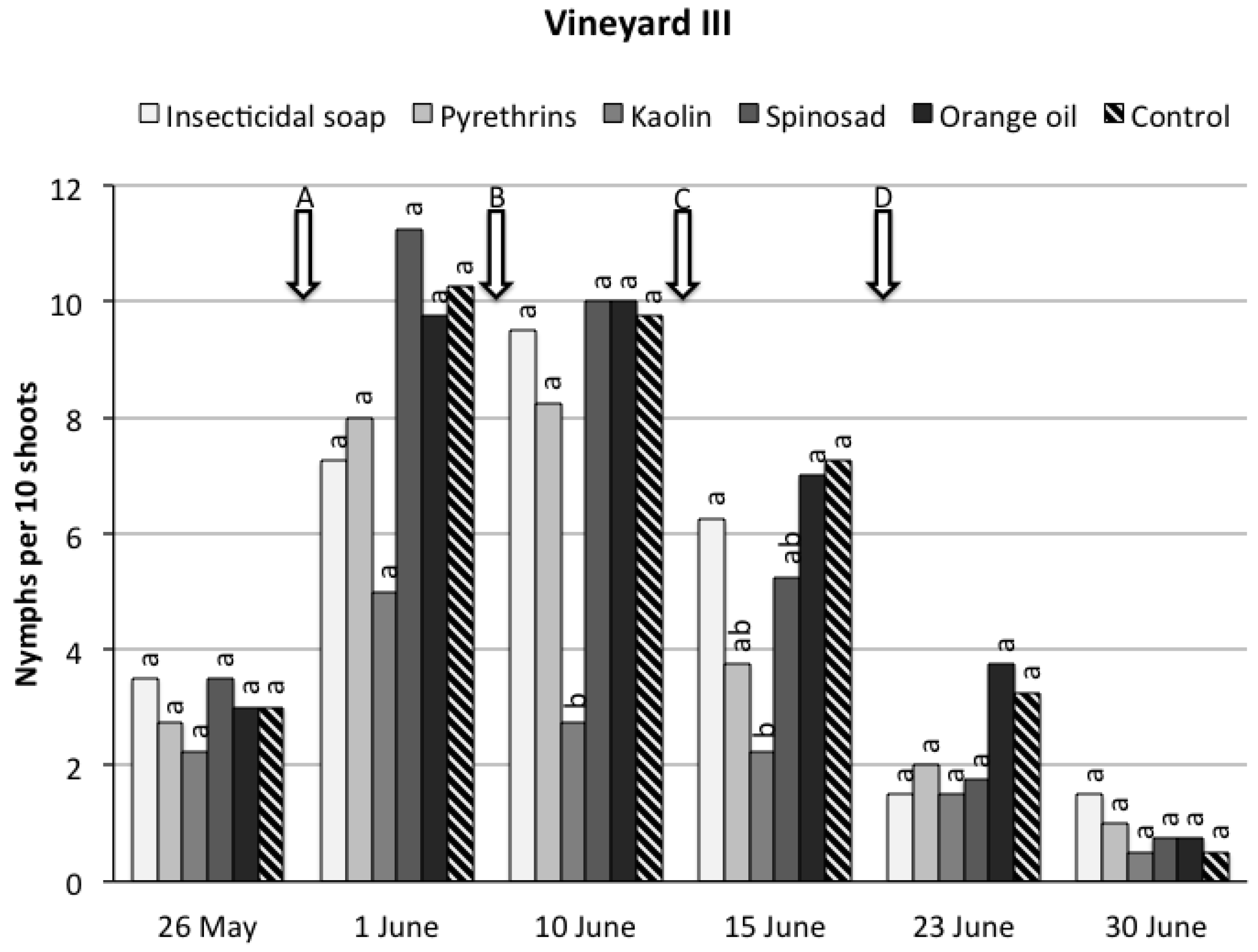
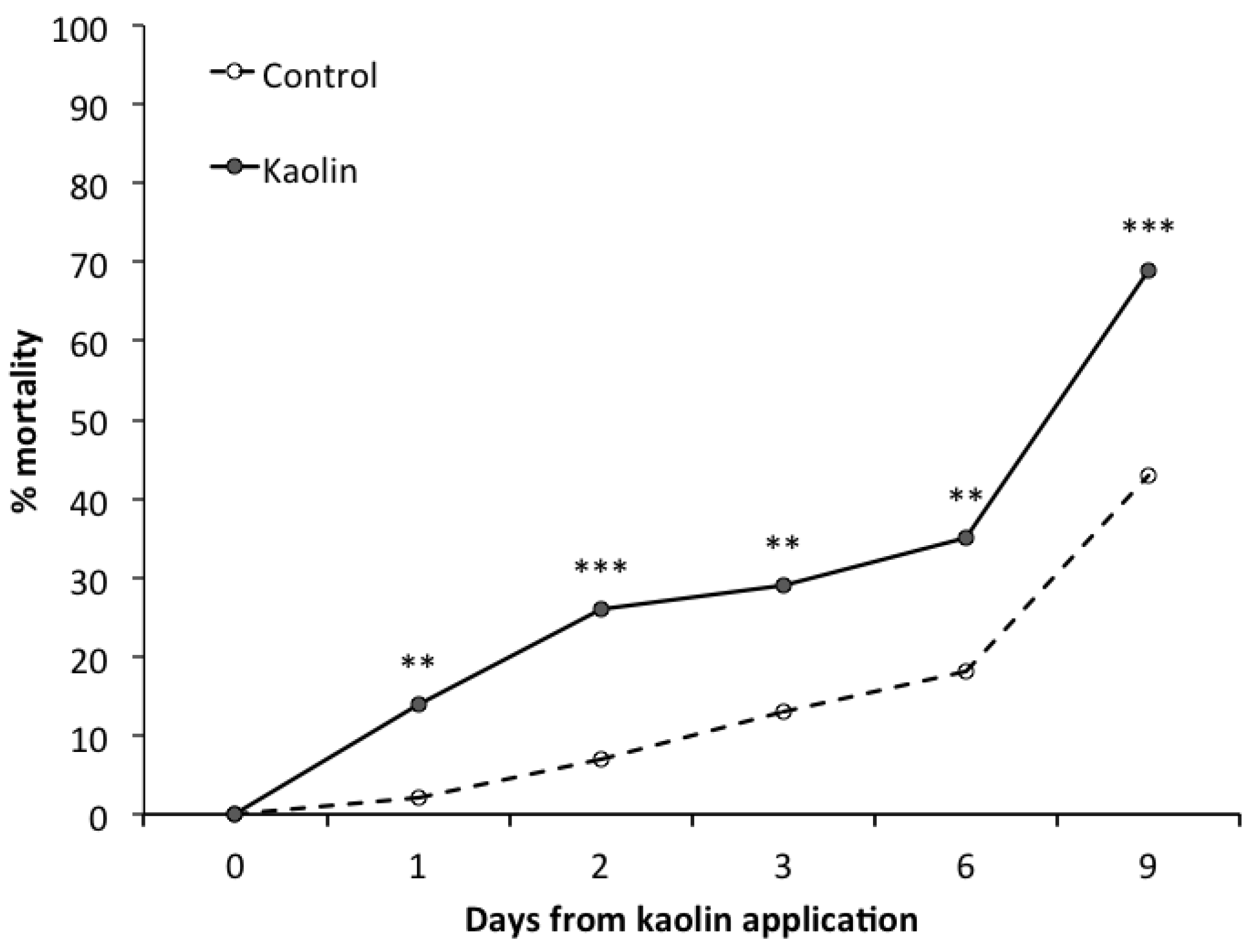
3. Results
3.1. Field Trials
3.2. Laboratory Bioassay
4. Discussion and Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Schvester, A.; Carle, P.; Moutous, G. Transmission de la Flavescence dorée de la vigne par Scaphoideus littoralis Ball. (Homopt. Jassidae). Ann. Épiphyt. 1963, 14, 175–198. (In French) [Google Scholar]
- Carraro, L.; Loi, N.; Kuszala, C.; Clair, D.; Boudon-Padieu, E.; Refatti, E. On the ability-inability of Scaphoideus titanus Ball to transmit different grapevine yellow agents. Vitis 1994, 33, 231–234. [Google Scholar]
- Bianco, P.A.; Alma, A.; Casati, P.; Scattini, G.; Arzone, A. Transmission of 16SrV phytoplasmas by Scaphoideus titanus Ball in Northern Italy. Plant Protect. Sci. 2001, 37, 49–56. [Google Scholar]
- Mori, N.; Bressan, A.; Martini, M.; Guadagnini, M.; Girolami, V.; Bertaccini, A. Experimental transmission by Scaphoideus titanus Ball of two Flavescence dorée-type phytoplasmas. Vitis 2002, 41, 99–102. [Google Scholar]
- EPPO/CABI. Quarantine Pests for Europe; CAB International: Wallingford, UK, 1997. [Google Scholar]
- Chuche, J.; Thiéry, D. Biology and ecology of the Flavescence dorée vector Scaphoideus titanus: A review. Agron. Sustain. Dev. 2014, 34, 381–403. [Google Scholar] [CrossRef]
- Caudwell, A. La flavescence dorée de la vigne en France. Phytoma-La Défense des végétaux 1981, 325, 16–19. (In French) [Google Scholar]
- Barba, M. La lotta obbligatoria alla flavescenza dorata e al suo vettore Scaphoideus titanus. In Flavescenza dorata e altri giallumi della vite in Toscana e in Italia; Quaderno ARSIA 2005; Bertaccini, A., Braccini, P., Eds.; ARSIA Regione Toscana: Firenze, Italy, 2005; Volume 3, pp. 135–138. (In Italian) [Google Scholar]
- Weintraub, P.G.; Beanland, L. Insect vectors of phytoplasmas. Annu. Rev. Entomol. 2006, 51, 91–111. [Google Scholar] [CrossRef] [PubMed]
- Carle, P.; Schvester, D. Nouvelle mise au point su la lutte contre Scaphoideus titanus Ball, cicadelle vectrice de la Flavescence dorée de la vigne. Rev. Zool. Agric. Appl. 1964, 63, 107–114. (In French) [Google Scholar]
- Pavan, F.; Mori, N.; Bigot, G.; Zandigiacomo, P. Border effect in spatial distribution of Flavescence dorée affected grapevines and outside source of Scaphoideus titanus vectors. Bull. Insectol. 2012, 65, 281–290. [Google Scholar]
- Lessio, F.; Portaluri, A.; Paparella, F.; Alma, A. A mathematical model of flavescence dorée epidemiology. Ecol. Model. 2015, 312, 41–53. [Google Scholar] [CrossRef]
- Pavan, F.; Stefanelli, G.; Villani, A.; Mori, N.; Posenato, G.; Bressan, A.; Girolami, V. Controllo della flavescenza dorata attraverso la lotta contro il vettore Scaphoideus titanus Ball. In Flavescenza dorata e altri giallumi della vite in Toscana e in Italia; Quaderno ARSIA 2005; Bertaccini, A., Braccini, P., Eds.; ARSIA Regione Toscana: Firenze, Italy, 2005; Volume 3, pp. 91–108. (In Italian) [Google Scholar]
- Bosco, D.; Mori, N. Flavescence dorée vector control in Italy. Phytopathogenic Mollicutes 2013, 3, 40–43. [Google Scholar] [CrossRef]
- Pavan, F.; Mori, N.; Bressan, S.; Mutton, P. Control strategies for grapevine phytoplasma diseases: Factors influencing the profitability of replacing symptomatic plants. Phytopathol. Mediterr. 2012, 51, 11–22. [Google Scholar]
- Schvester, D.; Moutous, G.; Bonfilis, J.; Carle, P. Étude biologique des cicadelles de la vigne dans le sud-ouest de la France. Ann. Épiphyt. 1962, 13, 205–237. (In French) [Google Scholar]
- Vidano, C. Scoperta in Italia dello Scaphoideus littoralis Ball cicalina americana collegata alla “Flavescence dorée” della vite. L'Italia Agricola 1964, 101, 1031–1049. (In Italian) [Google Scholar]
- Schvester, D.; Carle, P.; Moutous, G. Nouvelles données sur la transmission de la flavescence dorée de la vigne par Scaphoideus littoralis Ball. Ann. Zool. écol. Anim. 1969, 1, 445–465. (In French) [Google Scholar]
- Mori, N.; Posenato, G.; Sancassani, G.; Tosi, L.; Girolami, V. Insetticidi per il controllo delle cicaline nei vigneti. L'Informatore Agrario 1999, 55, 93–97. (In Italian) [Google Scholar]
- Posenato, G.; Mori, N.; Bressan, A.; Stefanelli, G.; Pavan, F.; Girolami, V. Valutazione dell'attività di alcuni insetticidi nei confronti degli adulti di Scaphoideus titanus Ball e Metcalfa pruinosa (Say). Atti Giornate Fitopatologiche 2002, 1, 459–462. (In Italian) [Google Scholar]
- Mazzoni, E.; Colla, R.; Chiusa, B.; Ciampitti, M.; Cravedi, P. Experiences for vector control of grape golden flavescence in Lombardia and Emilia Romagna (Northern Italy) vineyards. IOBC/WPRS Bull. 2003, 26, 221–225. [Google Scholar]
- Jermini, M.; Gusberti, M.; Schaub, L.; Linder, C.; Gugerli, P.; Schärer, S.; Emery, S. Flavescence dorée and Scaphoideus titanus: Distribution and control in Switzerland. IOBC/WPRS Bull. 2008, 36, 107–111. [Google Scholar]
- Žežlina, I.; Škvarč, A.; Bohinc, T.; Trdan, S. Testing the efficacy of single applications of five insecticides against Scaphoideus titanus on common grapevines. Int. J. Pest Manag. 2013, 59, 1–9. [Google Scholar] [CrossRef]
- Caobelli, R.; Carcereri, G. Lotta biologica alla cicalina della vite. L’Informatore Agrario 1995, 51, 75–77. (In Italian) [Google Scholar]
- Bottura, N.; Mori, N.; Posenato, G.; Sancassani, G.P.; Girolami, V. Lotta alle cicaline nei vigneti a conduzione biologica. L'Informatore Agrario 2003, 59, 75–79. (In Italian) [Google Scholar]
- Mori, N.; Bottura, N.; Posenato, G.; Sancassani, G.P.; Girolami, V. Lotta contro Scaphoideus titanus Ball nei vigneti a conduzione biologica. Atti Giornate Fitopatologiche 2004, 1, 111–116. (In Italian) [Google Scholar]
- Mori, N.; Tonello, D.; Posenato, G.; Pozzebon, A.; Duso, C. Efficacy of biopesticides against Scaphoides titanus Ball in different experimental conditions. IOBC/WPRS Bull. 2014, 105, 45–48. [Google Scholar]
- Gusberti, M.; Jermini, M.; Wyss, E.; Linder, C. Efficacité d’insecticides contre Scaphoideus titanus en vignobles biologiques et effets secondaires. Rev. Suisse Vitic. Arboric. Hortic. 2008, 40, 173–177. (In French) [Google Scholar]
- Constant, N. L'utilisation du pyrèthre naturel pour lutter contre la cicadelle de la flavescence dorée en viticulture biologique. Progrès Agricole et Viticole 2009, 126, 334–340. (In French) [Google Scholar]
- Duso, C.; Angeli, G.; Castagnoli, M.; Liguori, M.; Facchin, P.; Malagnini, V.; Pozzebon, A. Pesticides and phytoseiid mites: A synopsis of research carried out on grapes, apples and vegetables in northern and central Italy. In Acarology XI: Proceedings of the International Congress; Merida, Mexico, September 8–13, 2002; Morales-Malacara, J.B., Behan-Pelletier, V., Ueckermann, E., Pérez, T.M., Estrada, E., Gispert, C., Badii, M., Eds.; Merida, Instituto de Biololgía, UNAM; Facultad de Ciencias, UNAM; Sociedad Latinoamericana de Acarología: Ciudada Universitaria, Mexico, 2007; pp. 325–337. [Google Scholar]
- Tubajika, K.M.; Civerolo, E.L.; Puterka, G.J.; Hashim, J.M.; Luvisi, D.A. The effects of kaolin, harpin, and imidacloprid on development of Pierce’s disease in grape. Crop Prot. 2007, 26, 92–99. [Google Scholar] [CrossRef]
- Tacoli, F.; Pavan, F.; Cargnus, E.; Tilatti, E.; Pozzebon, A.; Zandigiacomo, P. Efficacy and mode of action of kaolin in the control of Empoasca vitis and Zygina rhamni (Hemiptera: Cicadellidae) in vineyards. J. Econ. Entomol. 2017, 110, 1164–1178. [Google Scholar] [CrossRef] [PubMed]
- Karamaouna, F.; Kimbaris, F.; Michaelakis, A.; Papachristos, D.; Polissiou, M.; Papatsakona, P.; Tsora, E. Insecticidal activity of plant essential oils against the vine mealybug, Planococcus ficus. J. Insect Sci. 2013, 13, 142. [Google Scholar] [CrossRef] [PubMed]
- Trdan, S.; Žnidarčič, D.; Valič, N. Field efficacy of three insecticides against cabbage stink bugs (Heteroptera: Pentatomidae) on two cultivars of white cabbage. Int. J. Pest Manag. 2006, 52, 79–87. [Google Scholar] [CrossRef]
- Lee, D.H.; Short, B.D.; Nielsen, A.L.; Leskey, T.C. Impact of organic insecticides on the survivorship and mobility of Halyomorpha halys (Hemiptera: Pentatomidae) in the laboratory. Florida Entomol. 2014, 97, 414–421. [Google Scholar] [CrossRef]
- Bagnoli, B.; Gargani, E. Survey on Scaphoideus titanus egg distribution on grapevines. IOBC/WPRS Bull. 2011, 67, 233–237. [Google Scholar]
- Abbott, W.S. A method for computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Pease, C.E.; Lόpez-Olguín, J.F.; Pérez-Moreno, I.; Marco-Mancebόn, V. Effects of kaolin on Lobesia botrana (Lepidoptera: Tortircidae) and its compatibility with the natural enemy, Trichogramma cacoeciae (Hymenoptera: Trichogrammatidae). J. Econ. Entomol. 2016, 109, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Tacoli, F.; Cargnus, E.; Kiaeian Moosavi, F.; Zandigiacomo, P.; Pavan, F.; University of Udine, Udine, Italy. Unpublished Data. 2017.
- Coniberti, A.; Ferrari, V.; Dellacassa, E.; Boido, E.; Carrau, F.; Gepp, V.; Disegna, E. Kaolin over sun-exposed fruit affects berry temperature, must composition and wine sensory attributes of Sauvignon blanc. Eur. J. Agron. 2013, 50, 75–81. [Google Scholar] [CrossRef]
- Brillante, L.; Belfiore, N.; Gaiotti, F.; Lovat, L.; Sansone, L.; Poni, S.; Tomasi, S. Comparing kaolin and pinolene to improve sustainable grapevine production during drought. PLoS ONE 2016, 11, e0156631. [Google Scholar] [CrossRef] [PubMed]
- Jansen, J.P.; Defrance, T.; Warnier, A.M. Effect of organic-farming-compatible insecticides on four aphid natural enemy species. Pest Manag. Sci. 2010, 66, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Pascual, S.; Cobos, G.; Seris, E.; González-Núñez, M. Effects of processed kaolin on pests and non-target arthropods in a Spanish olive grove. J. Pest Sci. 2010, 83, 121–133. [Google Scholar] [CrossRef]
- Pavan, F.; Bellomo, C.; Vidoni, F.; Bigot, G.; Ostan, M.; Boccalon, W.; Bressan, S.; Mutton, P.; Frausin, C.; De Biasio, A.C.; et al. Efficacia della lotta insetticida contro Scaphoideus titanus Ball in Friuli Venezia Giulia. Supplemento. Notiziario. ERSA 2005, 17, 11–21. (In Italian) [Google Scholar]
- Pozzebon, A.; Tirello, P.; Moret, R.; Pederiva, M.; Duso, C. A fundamental step in IPM on grapevine: Evaluating the side effects of pesticides on predatory mites. Insects 2015, 6, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, V.; Prešern, J.; Lucchi, A.; Virant-Doberlet, M. Reproductive strategy of the Nearctic leafhopper Scaphoideus titanus Ball (Hemiptera: Cicadellidae). Bull. Entomol. Res. 2009, 99, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, A.; Anfora, G.; Lucchi, A.; Lanzo, F.; Virant-Doberlet, M.; Mazzoni, V. Exploitation of insect vibrational signals reveals a new method of pest management. PLoS ONE 2012, 7, e32954. [Google Scholar] [CrossRef] [PubMed]
- Polajnar, J.; Eriksson, A.; Rossi Stacconi, M.V.; Lucchi, A.; Anfora, G.; Virant-Doberlet, M.; Mazzoni, V. The process of pair formation mediated by substrate-borne vibrations in a small insect. Behav. Process. 2014, 107, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Polajnar, J.; Eriksson, A.; Virant-Doberlet, M.; Mazzoni, V. Mating disruption of a grapevine pest using mechanical vibrations: From laboratory to the field. J. Pest Sci. 2016, 89, 909–921. [Google Scholar] [CrossRef]
- Alma, A.; Daffonchio, D.; Gonella, E.; Raddadi, N. Microbial symbionts of Auchenorrhyncha transmitting phytoplasmas: A resource for symbiotic control of phytoplasmoses. In Phytoplasmas: Genomes, Plant Hosts and Vectors; Weintraub, P.G., Jones, P., Eds.; CABI: Wallingford, UK, 2010; pp. 272–292. [Google Scholar]
- Gonella, E.; Crotti, E.; Rizzi, A.; Mandrioli, M.; Favia, G.; Daffonchio, D.; Alma, A. Horizontal transmission of the symbiotic bacterium Asaia sp. in the leafhopper Scaphoideus titanus Ball (Hemiptera: Cicadellidae). BMC Microbiol. 2012, 12 (Suppl. 1), S4. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, V.; Ioriatti, C.; Trona, F.; Lucchi, A.; De Cristofaro, A.; Anfora, G. Study on the role of olfaction in host plant detection of Scaphoideus titanus (Hemiptera: Cicadellidae) nymphs. J. Econ. Entomol. 2009, 102, 974–980. [Google Scholar] [CrossRef] [PubMed]
- Cara, C.; Trivellone, V.; Linder, C.; Junkert, J.; Jermini, M. Influence de la gestion des repousses du tronc et du bois de taille sur les densités de Scaphoideus titanus. Rev. Suisse Vitic. Arboric. Hortic. 2013, 45, 114–119. (In French) [Google Scholar]
- Trivellone, V.; Jermini, M.; Posenato, G.; Mori, N. Influence of pruning wood management and suckering on Scaphoideus titanus Ball density in two distinct wine-growing area. In Book of Abstracts IOBC-WPRS Meeting of the Working Group on “Integrated Protection and Production in Viticulture”; IOBC-WPRS: Vienna, Austria, 2015; p. 11. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Active Constituent | Commercial Product | Application Rate in Water | Application Timing * | |
|---|---|---|---|---|
| Name | Formulation | |||
| Kaolin | Surround WP (Tessenderlo Kerley Inc., Phoenix, Arizona, USA) | WP (wettable powder), 95% kaolin | 2% w/v | A, B, C |
| Orange oil | Prev-Am Plus (Nufarm Italia, Milano, Italy) | SL (soluble liquid), 5.88% orange oil | 0.5% v/v | C, D |
| Insecticidal soap | Flipper (Dow Agrosciences Italia, Milano, Italy) | SL (soluble liquid), 47.8% potassium salts of fatty acids | >2% v/v | C, D |
| Spinosad | Laser (Dow Agrosciences Italia, Milano, Italy) | SC (suspension concentrate), 44.2% pure spinosad | 0.02% v/v | C, D |
| Pyrethrins | Biopiren Plus (Copyr, Milano, Italy) | EC (emulsifiable concentrate), 2% pure pyrethrins | 0.16% v/v | C, D |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tacoli, F.; Mori, N.; Pozzebon, A.; Cargnus, E.; Da Vià, S.; Zandigiacomo, P.; Duso, C.; Pavan, F. Control of Scaphoideus titanus with Natural Products in Organic Vineyards. Insects 2017, 8, 129. https://doi.org/10.3390/insects8040129
Tacoli F, Mori N, Pozzebon A, Cargnus E, Da Vià S, Zandigiacomo P, Duso C, Pavan F. Control of Scaphoideus titanus with Natural Products in Organic Vineyards. Insects. 2017; 8(4):129. https://doi.org/10.3390/insects8040129
Chicago/Turabian StyleTacoli, Federico, Nicola Mori, Alberto Pozzebon, Elena Cargnus, Sarah Da Vià, Pietro Zandigiacomo, Carlo Duso, and Francesco Pavan. 2017. "Control of Scaphoideus titanus with Natural Products in Organic Vineyards" Insects 8, no. 4: 129. https://doi.org/10.3390/insects8040129
APA StyleTacoli, F., Mori, N., Pozzebon, A., Cargnus, E., Da Vià, S., Zandigiacomo, P., Duso, C., & Pavan, F. (2017). Control of Scaphoideus titanus with Natural Products in Organic Vineyards. Insects, 8(4), 129. https://doi.org/10.3390/insects8040129