
Jamaica’s Critically Endangered Butterfly: A Review of the Biology and Conservation Status of the Homerus Swallowtail (Papilio (Pterourus) homerus Fabricius)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Papilio homerus Historical Records
3. Morphology and Life History Traits
3.1. Eggs
3.2. Larval Stages
3.3. Pre-pupae
3.4. Pupae
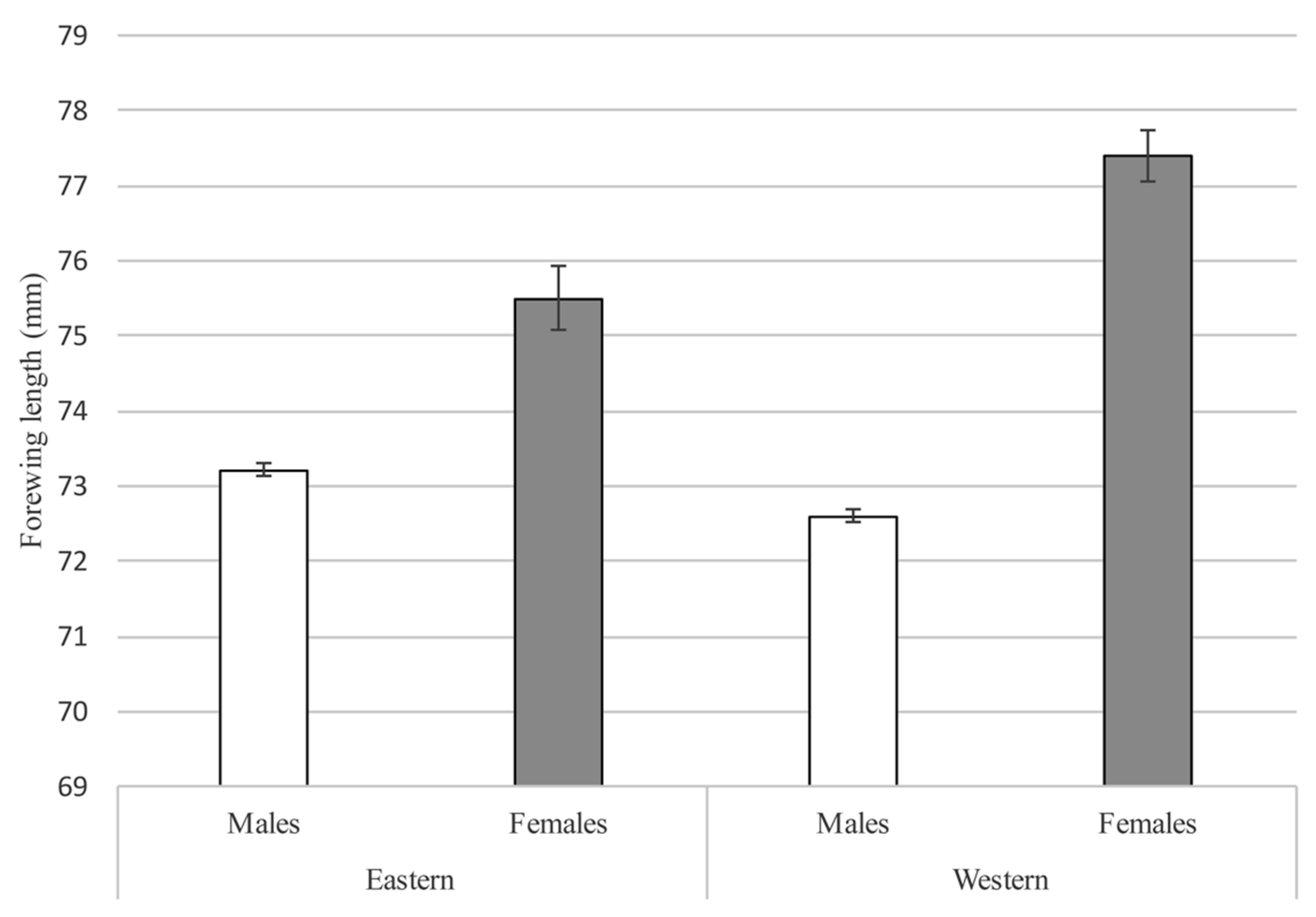
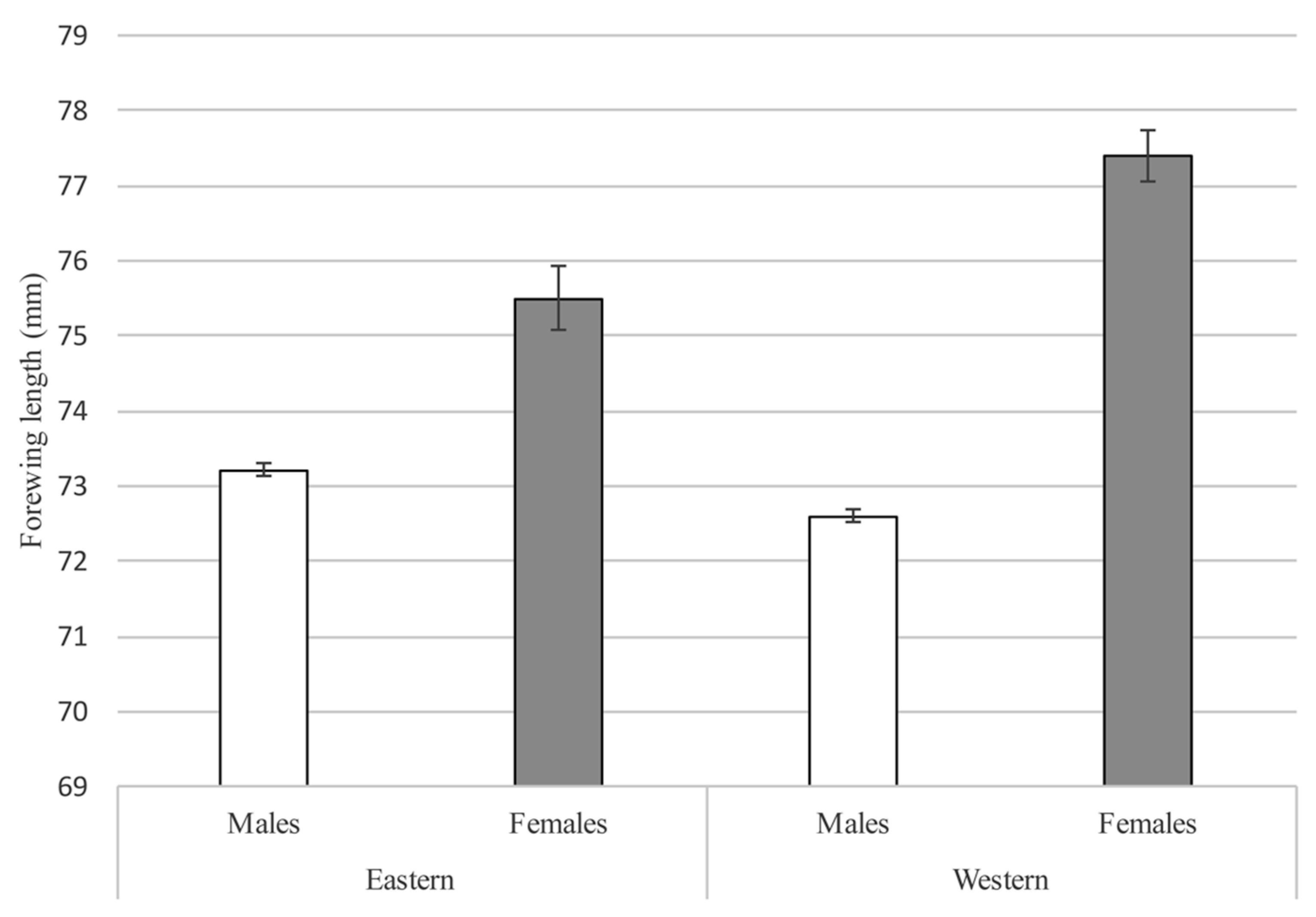
3.5. Adults
4. Papilio homerus Biology
4.1. Host Plants and Behavior of Larvae
4.2. Adult Population Size Estimates and Behavior
5. Phylogeny and Biogeography
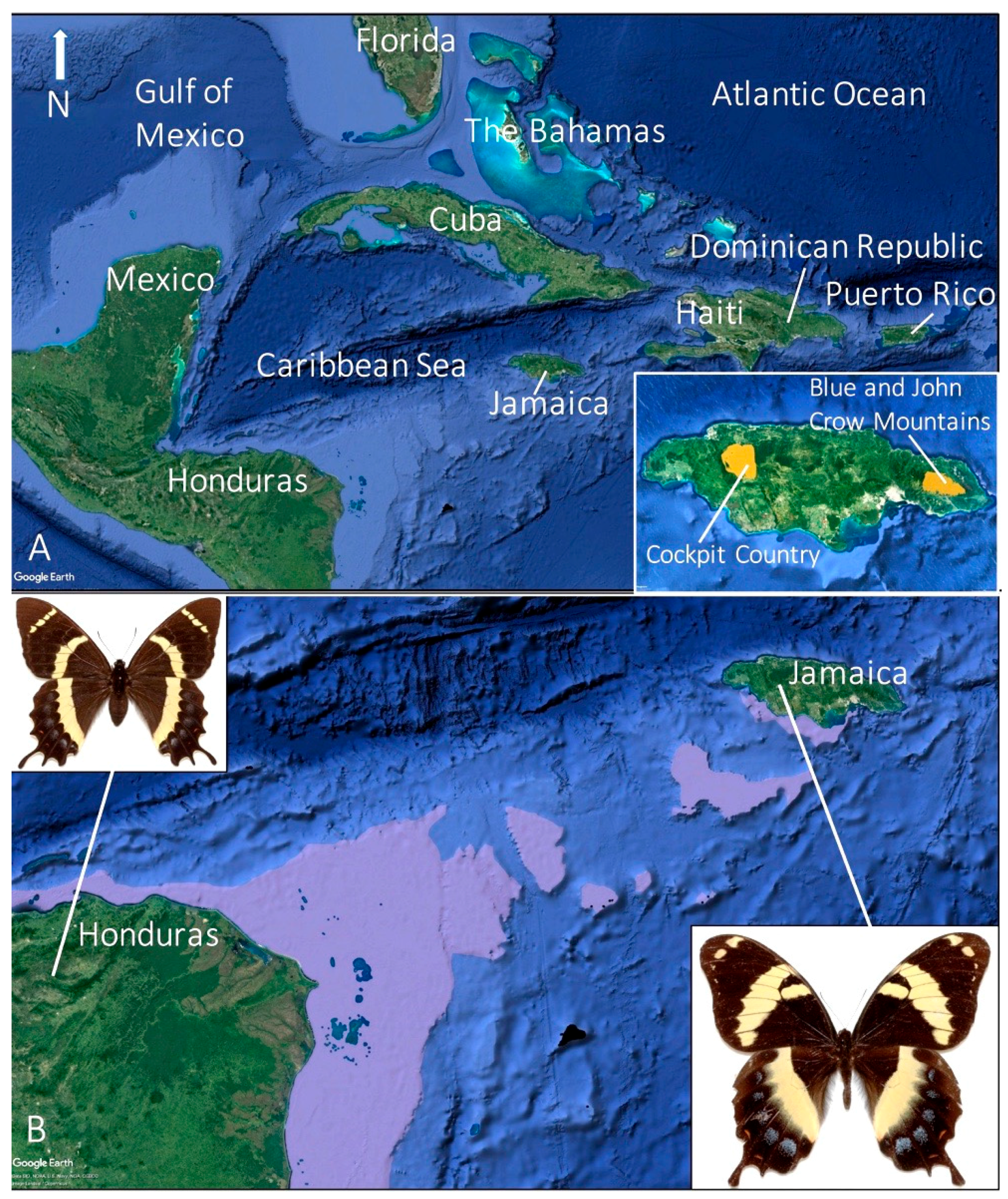
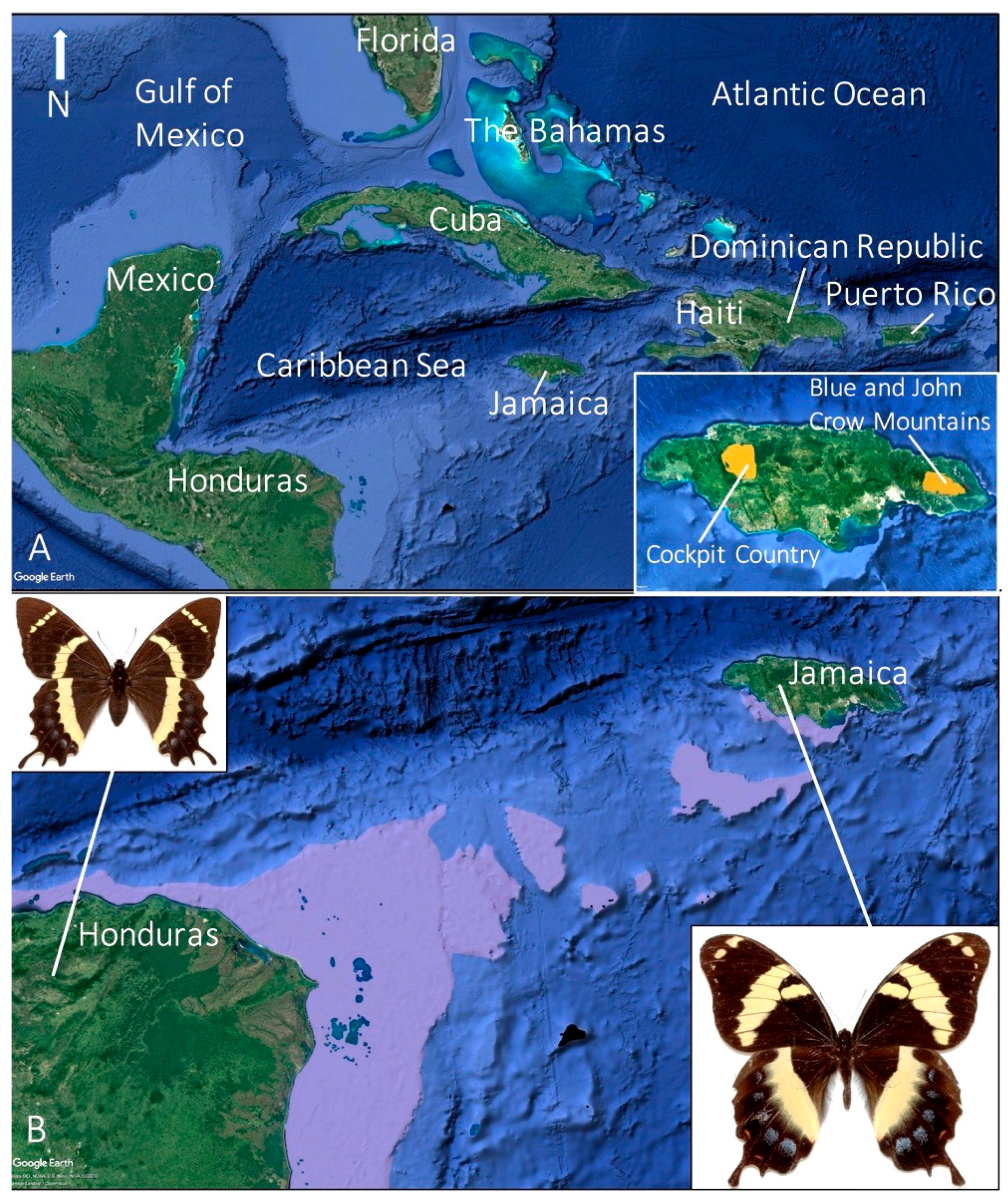
6. Past and Present Distributions and Habitat Preference
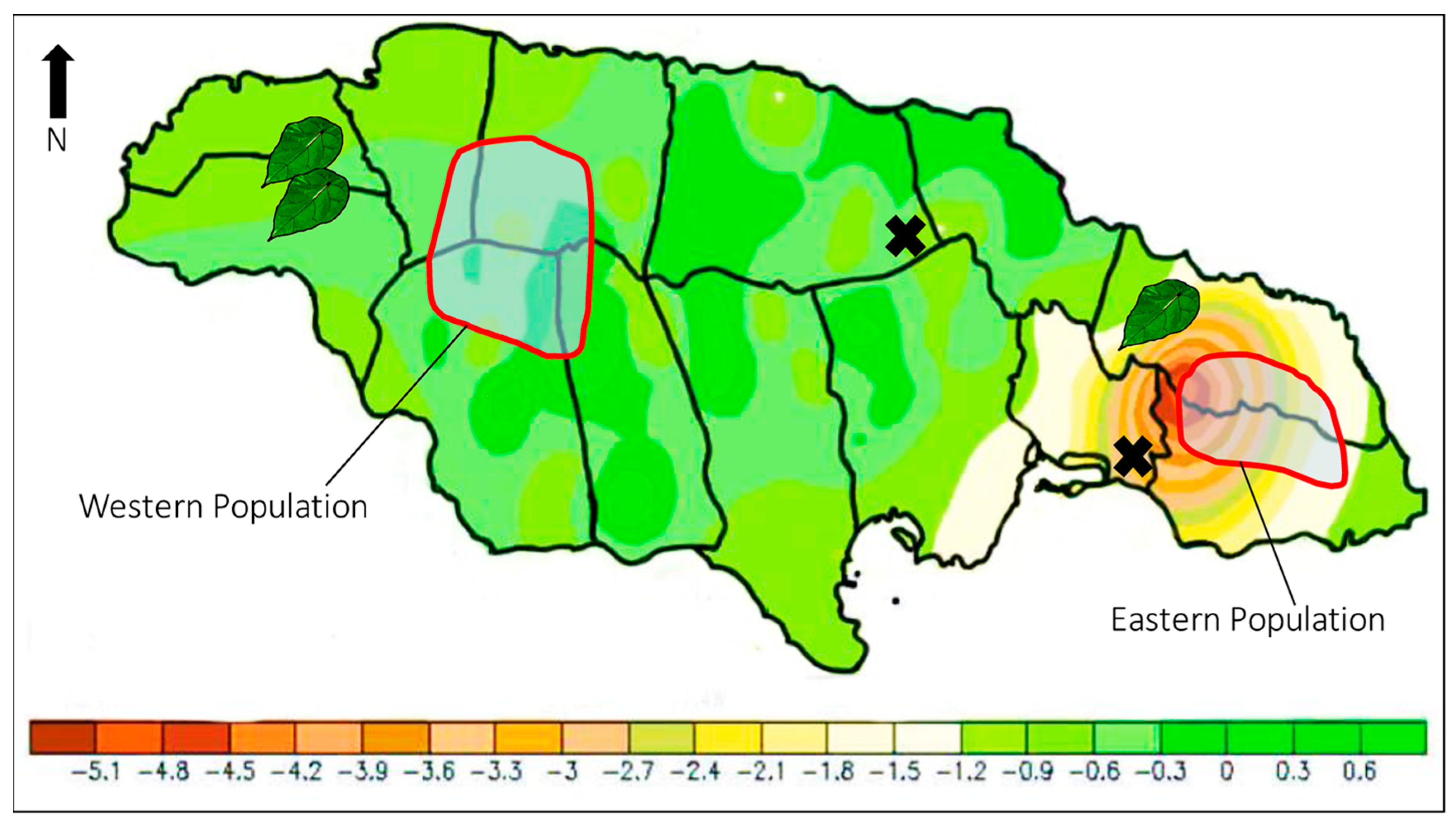
6.1. Central Population
6.2. Eastern Population
6.3. Western Population
7. Threats to Populations of Papilio homerus
7.1. Habitat Destruction
7.2. Predators and Parasitoids
7.3. Poaching
7.4. Climate Change
8. Invasive Species
9. Knowledge Gaps and Objectives for a Conservation Plan
9.1. Determine the Current Geographic Distribution and Estimate the Total Number of Individuals in the Stronghold Populations
9.2. Determine the Spatial and Temporal Dynamics of P. homerus Subpopulations
9.3. Survey Areas to Locate Suitable Habitat for the Potential Release of Captive-Bred P. homerus
9.4. Regularly Monitor Habitat Quality in Areas where the Current Populations Exist and where Future Populations Could Be Introduced
9.5. Create Protected Areas of Montane Forest
9.6. Conduct Research to Determine Ecological Threats to P. homerus and Potential Mitigation Strategies
9.7. Develop Ex Situ Conservation and Recovery Program
9.8. Assess Level of Genetic Differentiation between Spatially Discrete Populations
9.9. Educate the Local Public on the Status of the Populations, Conservation Efforts, and the Importance of Keeping These Populations Protected
10. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Smith, D.S.; Miller, L.D.; Miller, J.Y. Butterflies of the West Indies and South Florida; Oxford University Press: Oxford, UK, 1994; p. 264. [Google Scholar]
- Tyler, H.; Brown, K.S.; Wilson, K.H. Swallowtail Butterflies of the Americas. A Study in Biological Dynamics, Ecological Diversity, Biosystematics, and Conservation; Scientific Publishers: Gainesville, FL, USA, 1994; p. 376. [Google Scholar]
- Collins, N.M.; Morris, M.G. Threatened Swallowtail Butterflies of the World: The IUCN Red Data Book; International Union for Conservation of Nature: Gland, Switzerland; Cambridge, UK, 1985; p. 401. [Google Scholar]
- Lehnert, M.S. The population biology and ecology of the Homerus swallowtail, Papilio (Pterourus) homerus, in the Cockpit Country, Jamaica. J. Insect Conserv. 2008, 12, 179–188. [Google Scholar] [CrossRef]
- Emmel, T.C. Saving endangered swallowtails: The conservation biology of Papilio aristodemus ponceanus in Florida and P. homerus in Jamaica. In Swallowtail Butterflies: Their Ecology and Evolutionary Biology; Scriber, J.M., Tsubaki, Y., Lederhouse, R.C., Eds.; Scientific Publishers: Gainesville, FL, USA, 1995; pp. 359–369. [Google Scholar]
- Emmel, T.C.; Garraway, E. Ecology and conservation biology of the Homerus swallowtail in Jamaica (Lepidoptera: Papilionidae). Trop. Lepidoptera 1990, 1, 63–76. [Google Scholar]
- Garraway, E.; Bailey, A.J.A.; Freemen, B.E.; Parnell, J.R.; Emmel, T.C. Population studies and conservation of Jamaica’s endangered swallowtail butterfly Papilio (Pterourus) homerus. J. Insect Conserv. 2008, 12, 383–397. [Google Scholar] [CrossRef]
- Barua, M.; Gurdak, D.J.; Ahmed, R.A.; Tamuly, J. Selecting flagships for invertebrate conservation. Biodivers. Conserv. 2012, 21, 1457–1476. [Google Scholar] [CrossRef]
- Neufville, Z. Environment-Jamaica: Bauxite Mining Blamed for Deforestation. Inter Press Service. 2001. Available online: http://www.ipsnews.net/2001/04/environment-jamaica-bauxite-mining-blamed-for-deforestation/ (accessed on 20 June 2017).
- Turner, T.W. Papilio homerus (Papilionidae) in Jamaica, West Indies: field observations and a description of the immature stages. J. Lepidopterists Soc. 1991, 45, 259–271. [Google Scholar]
- Garraway, E.; Herlitz, A.D.; Snyder, N.; Bailey, A.J.A. New populations of the Jamaican giant swallowtail, Papilio (Pterourus) homerus (Lepidoptera: Papilionidae). Trop. Lepidoptera Res. 2008, 18, 43–45. [Google Scholar]
- Fabricius, J.C. Entomologia Systematica Emendata Et Aucta. Secundum Classes, Ordines, Genera, Species Adjectis Synonimis, Locis, Observationibus, Descriptionibus; Impensis Christ. Gottl. Proft: Hafniae, 1793; p. 519. [Google Scholar]
- Vane-Wright, R.I.; Hughes, H.W.D. The first images of Homer’s Swallowtail (Lepidoptera: Papilionidae). Linnean 2004, 20, 31–41. [Google Scholar]
- Esper, E.J.C. Die ausländische oder die ausserhalb Europa zur Zeit in den übrigen Welttheilen vorgefundene Schmetterlinge in Abbildungen nach der Natur mit Beschreibungen; Wolfgang Walther: Erlangen, Germany, 1796. [Google Scholar]
- Rutherford, D.G. Notes regarding some rare papilios. Entomol. Mag. 1878, 15, 28–31. [Google Scholar]
- Gosse, P.H. On Papilio homerus, its ovum and larva. Proc. Entomol. Soc. Lond. 1879, 1879, lv–lviii. [Google Scholar]
- Panton, E.S. A description of the larva of Papilio homerus. J. Inst. Jamaica (Kingston) 1893, 1, 375–376. [Google Scholar]
- Taylor, C.B. Description of the larva and pupa of Papilio homerus, Fab. Trans. Entomol. Soc. Lond. 1894, 42, 409–410. [Google Scholar] [CrossRef]
- Avinoff, A.; Shoumatoff, N. Jamaican summer. Carnegie Mag. 1940, 14, 175–182. [Google Scholar]
- Brown, F.M.; Heineman, B. Jamaica and Its Butterflies; E.W. Classey: London, UK, 1972; p. 478. [Google Scholar]
- Garraway, E.; Bailey, A. Parasitoid induced mortality in the eggs of the endangered giant swallowtail butterfly Papilio homerus (Papilionidae). J. Lepidopterists Soc. 1992, 46, 233–234. [Google Scholar]
- Garraway, E.; Parnell, J.R. Notes on the osmeteria of Papilio homerus (Lepidoptera: Papilionidae). Trop. Lepidoptera 1993, 4, 29–30. [Google Scholar]
- Garraway, E.; Bailey, A.J.A.; Emmel, T.C. Contribution to the ecology and conservation biology of the endangered Papilio homerus (Lepidoptera: Papilionidae). Trop. Lepidoptera 1993, 4, 83–91. [Google Scholar]
- Lehnert, M.S.; Emmel, T.C.; Garraway, E. Male-Male interactions in the endangered Homerus swallowtail, Papilio homerus (Lepidoptera: Papilionidae), in Jamaica. Caribb. J. Sci. 2013, 47, 57–66. [Google Scholar] [CrossRef]
- Miller, W.E. Wing measure as a size index in Lepidoptera: The family Olethreutidae. Ann. Entomol. Soc. Am. 1977, 70, 253–256. [Google Scholar] [CrossRef]
- Futahashi, R.; Fujiwara, H. Identification of stage-specific larval camouflage associated genes in the swallowtail butterfly, Papilio xuthus. Dev. Genes Evol. 2008, 218, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Bailey, A.J.A. The Biology and Ecology of the Endangered Giant Swallowtail Butterfly, Papilio (Pterourus) homerus (Lepidoptera: Papilionidae) in Jamaica. Ph.D. Thesis, University of the West Indies, Mona, Jamaica, 2003. [Google Scholar]
- Lewis, C.B. Butterfly notes. Nat. Hist. Notes, Nat. His. Soc. Jamaica 1949, 4, 48. [Google Scholar]
- Swainson, E.M. Notes on lepidopterous larvae from Jamaica. J. N. Y. Entomol. Soc. 1901, 9, 77–82. [Google Scholar]
- Parnell, J. Papilio homerus; the Vanishing Swallowtail; Documentary Video; University of the West Indies: Mona, Jamaica, 1984. [Google Scholar]
- Lewis, C.B. Butterfly notes. Nat. Hist. Notes, Nat. His. Soc. Jamaica 1948, 3, 204. [Google Scholar]
- Lehnert, M.S.; Monaenkova, D.; Andrukh, T.; Beard, C.E.; Adler, P.H.; Kornev, K.G. Hydrophobic–hydrophilic dichotomy of the butterfly proboscis. J. R. Soc. Interface 2013, 10, doi–10. [Google Scholar] [CrossRef] [PubMed]
- Lehnert, M.S.; Mulvane, C.P.; Brothers, A. Mouthpart separation does not impede butterfly feeding. Arthropod Struct. Dev. 2014, 43, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Lehnert, M.S.; Beard, C.E.; Gerard, P.D.; Kornev, K.G.; Adler, P.H. Structure of the lepidopteran proboscis in relation to feeding guild. J. Morphol. 2016, 277, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Arms, K.; Feeny, P.; Lederhouse, R.C. Sodium: Stimulus for puddling behavior by tiger swallowtail butterflies, Papilio glaucus. Science 1974, 185, 372–374. [Google Scholar] [CrossRef] [PubMed]
- Adler, P.H.; Pearson, D.L. Why do male butterflies visit mud puddles? Can. J. Zool. 1982, 60, 322–325. [Google Scholar] [CrossRef]
- Frietas, A.V.L.; Benson, W.W.; Marini-Filho, O.J.; de Carvalho, R.M. Territoriality by the dawn’s early light: The neotropical owl butterfly Caligo idomeneus (Nymphalidae: Brassolinae). J. Res. Lepidoptera 1997, 34, 14–20. [Google Scholar]
- Wickman, P.O.; Wiklund, C. Territorial defence and its seasonal decline in the speckled wood butterfly (Pararge aegeria). Anim. Behav. 1983, 31, 1206–1216. [Google Scholar] [CrossRef]
- Davies, N.B. Territorial defence in the speckled wood butterfly (Pararge aegeria): The resident always wins. Anim. Behav. 1978, 26, 138–147. [Google Scholar] [CrossRef]
- Lehnert, M.S. New protocol for measuring Lepidoptera wing damage. J. Lepidopterists Soc. 2010, 64, 29–32. [Google Scholar] [CrossRef]
- Zakharov, E.V.; Caterino, M.S.; Sperling, F.A. Molecular phylogeny, historical biogeography, and divergence time estimates for swallowtail butterflies of the genus Papilio (Lepidoptera: Papilionidae). Syst. Biol. 2004, 53, 193–215. [Google Scholar] [CrossRef] [PubMed]
- Häuser, C.; de Jong, R.; Lamas, G.; Robbins, R.; Smith, C.; Vane-Wright, R.I. Papilionidae–Revised GloBIS/GART Species Checklist (2nd Draft). 2005. Available online: http://www.insects-online.de/frames/papilio.htm (accessed on 20 June 2017).
- Simonsen, T.J.; Zakharov, E.V.; Djernaes, M.; Cotton, A.M.; Vane-Wright, R.I.; Sperling, F.A.H. Phylogenetics and divergence times of Papilioninae (Lepidoptera) with special reference to the enigmatic genera Teinopalpus and Meandrusa. Cladistics 2010, 27, 113–137. [Google Scholar] [CrossRef]
- Wu, L.-W.; Yen, S.-H.; Lees, D.C.; Lu, C.-C.; Yang, P.-S.; Hsu, Y.-F. Phylogeny and historical biogeography of Asian Pterourus butterflies (Lepidoptera: Papilionidae): A case of intercontinental dispersal from North America to East Asia. PLoS ONE 2015, 10, e0140933. [Google Scholar] [CrossRef] [PubMed]
- Beutelspacher, C.R.; Howe, W.H. Mariposas de Mexico; La Prensa Medica Mexicana: Mexico City, Mexico, 1984; p. 127. [Google Scholar]
- Morgan, G.S. Quaternary land vertebrates of Jamaica. Geol. Soc. Am. Mem. 1993, 182, 417–442. [Google Scholar]
- Kaye, W.J. The butterflies of Jamaica. Trans. Entomol. Soc. Lond. 1926, 73, 455–504. [Google Scholar]
- Robinson, W. A trip after Papilio homerus. Entomol. News 1903, 14, 17–21. [Google Scholar]
- Longstaff, G.B.; Müller, F. Butterfly-Hunting in Many Lands: Notes of a Field Naturalist; Longmans, Green, and Co.: London, UK, 1912; p. 729. [Google Scholar]
- Barry, A. Butterfly Notes. Nat. Hist. Notes, Nat. Hist. Soc. Jam. (Kingston) 1946, 11, 161–163. [Google Scholar]
- Headley, M.; Evelyn, O.B. Jamaica Country Report. Proceedings of the Sub-Regional Workshop on Data Collection and Outlook Effort for Forestry in the Caribbean. 2000. Available online: http://www.fao.org/docrep/003/x6689e/X6689E02.htm (accessed on 20 June 2017).
- Cockpit Country Stakeholders Group in Association with Jamaica Environmental Advocacy Network (JEAN). Save Cockpit Country Fact Sheet. Cockpit Country Stakeholders Group. 2007. Available online: www.jamaicancaves.ord/Save_Cockpit_Country_Fact_Sheet_2007.pdf (accessed on 20 June 2017).
- Lovejoy, T.E.; Bierregaard, R.O.; Rylands, A.B.; Malcolm, J.R.; Quintela, C.E.; Harper, L.H.; Brown, K.S.; Powell, A.H.; Powell, G.V.N.; Schubart, H.O.R.; et al. Edge and other effects of isolation on Amazon forest fragments. In Conservation Biology: The Science of Scarcity and Diversity; Soule, M.E., Ed.; Sinauer: Sunderland, MA, USA, 1986; pp. 257–285. [Google Scholar]
- Laurance, W.F. Forest-climate interactions in fragmented tropical landscapes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Kapos, V.; Ganade, G.; Matsui, E.; Victoria, R.L. ∂^13C as an indicator of edge effects in tropical rainforest reserves. J. Ecol. 1993, 81, 425–432. [Google Scholar] [CrossRef]
- Bruna, E.M. Effects of forest fragmentation on Heliconia acuminata seedling recruitment in central Amazonia. Oecologia 2002, 132, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Lehnert, M.S. Ecology and Population Biology of the Jamaican Giant Swallowtail, Papilio (Pterourus) homerus Fabricius (Lepidoptera: Papilionidae), in the Cockpit Country, Jamaica. Master’s Thesis, University of Florida, Gainesville, FL, USA, 2005. [Google Scholar]
- Mimura, N.; Nurse, L.; McLean, R.; Agard, J.; Briguglio, L.; Lefale, P.; Payet, R.; Sem, G. Small islands. In Climate Change: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Parry, M.L., Canziani, O.F., Palutikof, J.P., van der Linden, P.J., Hanson, C.E., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 687–716. [Google Scholar]
- Singh, B. Climate-related global changes in the southern Caribbean: Trinidad and Tobago. Glob. Planet. Chang. 1997, 15, 93–111. [Google Scholar] [CrossRef]
- Peterson, A.T.; Ortega-Huerta, M.A.; Bartley, J.; Sánchez-Cordero, V.; Soberón, J.; Buddemeier, R.H.; Stockwell, D.R. Future projections for Mexican faunas under global climate change scenarios. Nature 2002, 416, 626–629. [Google Scholar] [CrossRef] [PubMed]
- Ángeles, M.E.; González, J.E.; Erickson, D.J.; Hernández, J.L. Predictions of future climate change in the Caribbean region using global general circulation models. Int. J. Climatol. 2007, 27, 555–569. [Google Scholar] [CrossRef]
- Gamble, D.W.; Parnell, D.B.; Curtis, S. Caribbean vulnerability: Development of an appropriate climatic framework. In Global Change and Caribbean Vulnerability: Environment, Economy and Society at Risk? McGregor, D., Dodman, D., Barker, D., Eds.; University of the West Indies Press: Kingston, Jamaica, 2009; pp. 22–46. [Google Scholar]
- MacPhee, R.D.E.; Greenwood, A.D. Infectious disease, endangerment, and extinction. Int. J. Evol. Biol. 2013. [Google Scholar] [CrossRef]



© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lehnert, M.S.; Kramer, V.R.; Rawlins, J.E.; Verdecia, V.; Daniels, J.C. Jamaica’s Critically Endangered Butterfly: A Review of the Biology and Conservation Status of the Homerus Swallowtail (Papilio (Pterourus) homerus Fabricius). Insects 2017, 8, 68. https://doi.org/10.3390/insects8030068
Lehnert MS, Kramer VR, Rawlins JE, Verdecia V, Daniels JC. Jamaica’s Critically Endangered Butterfly: A Review of the Biology and Conservation Status of the Homerus Swallowtail (Papilio (Pterourus) homerus Fabricius). Insects. 2017; 8(3):68. https://doi.org/10.3390/insects8030068
Chicago/Turabian StyleLehnert, Matthew S., Valerie R. Kramer, John E. Rawlins, Vanessa Verdecia, and Jaret C. Daniels. 2017. "Jamaica’s Critically Endangered Butterfly: A Review of the Biology and Conservation Status of the Homerus Swallowtail (Papilio (Pterourus) homerus Fabricius)" Insects 8, no. 3: 68. https://doi.org/10.3390/insects8030068
APA StyleLehnert, M. S., Kramer, V. R., Rawlins, J. E., Verdecia, V., & Daniels, J. C. (2017). Jamaica’s Critically Endangered Butterfly: A Review of the Biology and Conservation Status of the Homerus Swallowtail (Papilio (Pterourus) homerus Fabricius). Insects, 8(3), 68. https://doi.org/10.3390/insects8030068