Microfungi Associated with Pteroptyx bearni (Coleoptera: Lampyridae) Eggs and Larvae from Kawang River, Sabah (Northern Borneo)


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Specimen Collection
2.2. Isolation and Identification of Fungi
2.3. Microscopic Examination
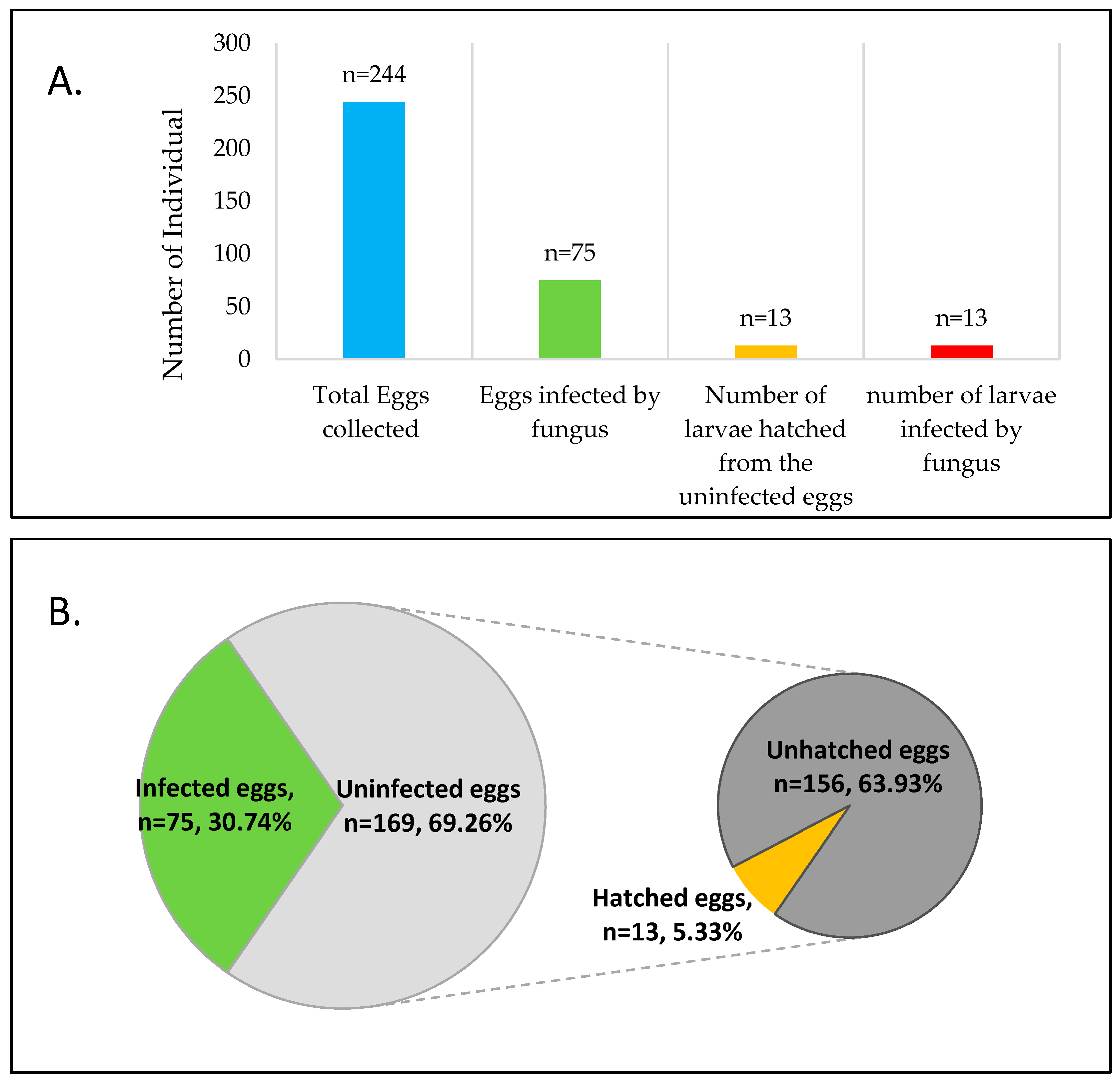
3. Results
3.1. Microfungi Associated with Pteroptyx bearni Eggs and Larvae
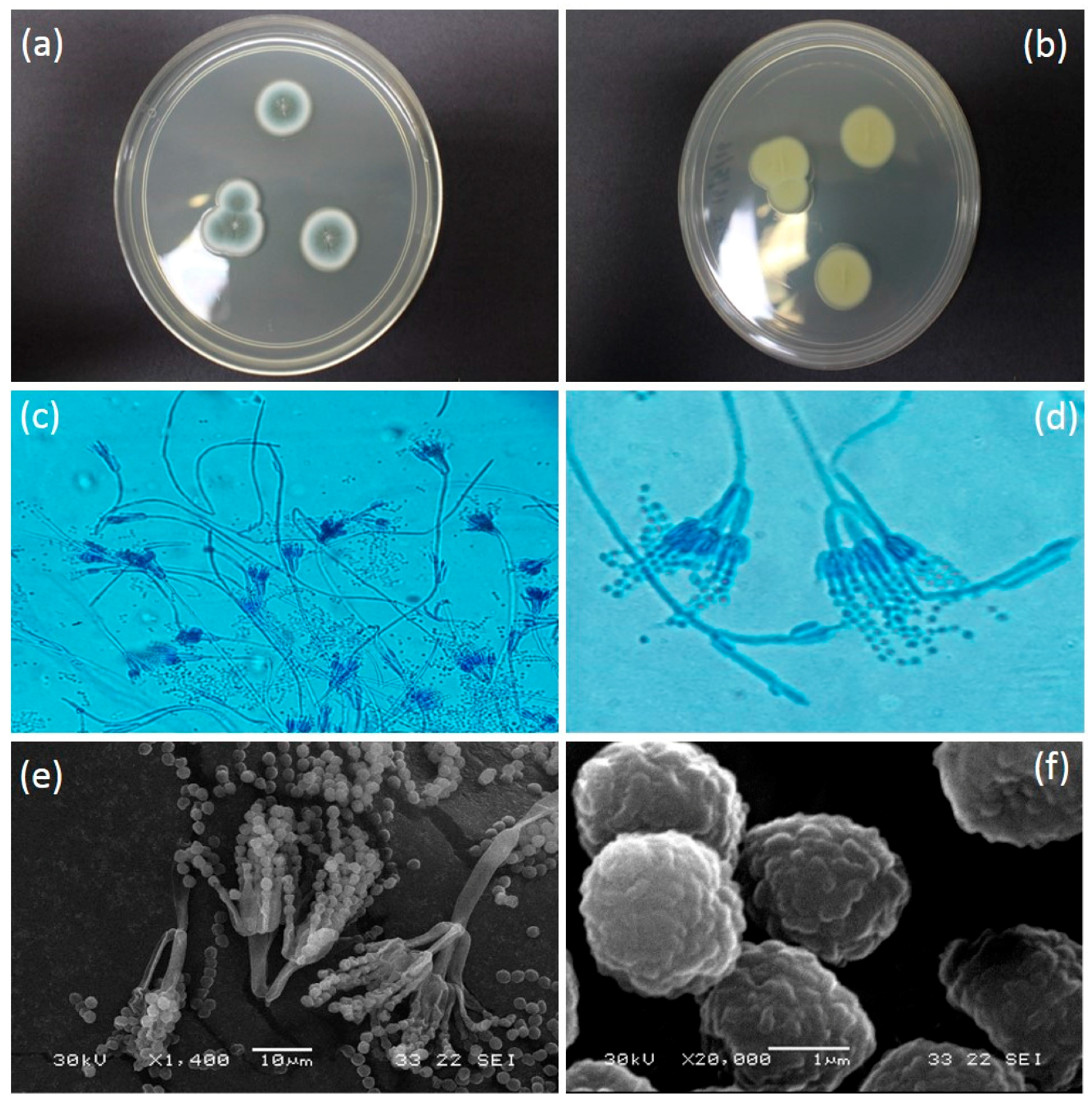
3.2. Penicillium Citrinum Thom
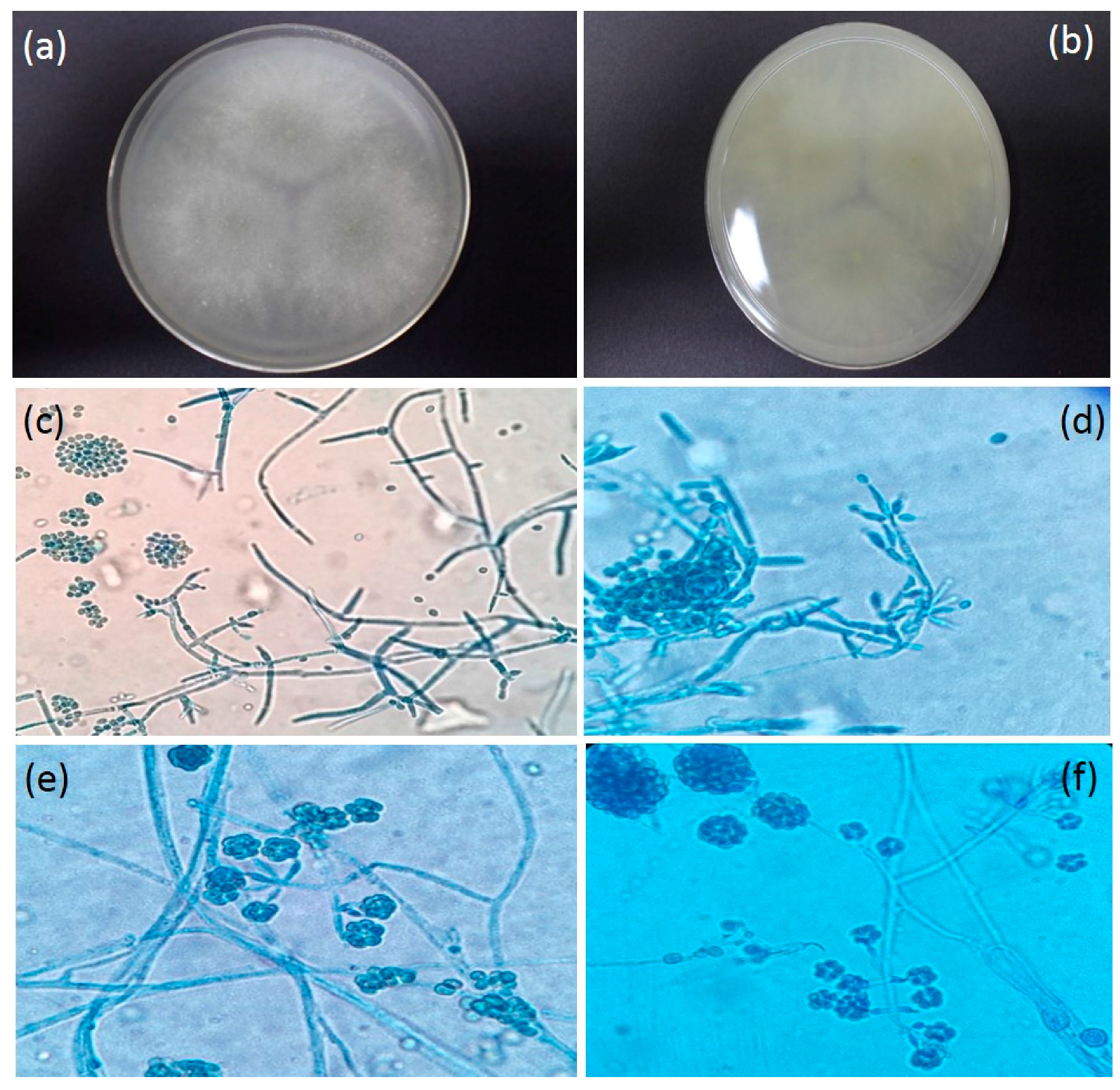
3.3. Trichoderma Harzianum Rifai
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Blackwell, M. The Fungi: 1, 2, 3 … 5.1 million species? Am. J. Bot. 2011, 98, 426–438. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, I.; Pereira, J.A.; Lino-Neto, T.; Bento, A.; Baptista, P. Fungal diversity associated to the olive moth, Prays oleae Bernard: A survey for potential entomopathogenic fungi. Microb. Ecol. 2012, 63, 964–974. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Najera, R.; Ruiz-Estudillo, R.A.; Sanchez-Yanez, J.M.; Molina-Ochoe, J.; Skoda, S.R.; Coutino-Ruiz, R.; Pinto-Ruiz, R.; Guevara-Hernandez, F.; Foster, J.E. Occurrence of entomopathogenic fungi and parasitic nematodes on Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae collected in Central Chiapas, Maxico. Fla. Entomol. 2013, 96, 498–503. [Google Scholar] [CrossRef]
- Rios-Velasco, C.; Gallegos-Morales, G.; Cambero-Campos, J.; Cerna-Chavez, E.; Rincon-Castro, M.C.D.; Valenzuela-Garcia, R. Natural enemies of the fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) In Coahuila, Mexico. Fla. Entomol. 2011, 94, 723–726. [Google Scholar] [CrossRef]
- Pereira, E.S.; Hamada, N.; Ferreira, R.L.M.; Lichtwardt, R.W. Trichomycete fungi (Zygomycota) associated with mosquito larvae (Diptera: Culicidae) in natural and artificial habitats in Manaus, AM, Brazil. Neotrop. Entomol. 2005, 34, 325–329. [Google Scholar] [CrossRef]
- Pereira, E.S.; Sarquis, M.I.M.; Ferreira-Keppler, R.; Hamada, N.; Alencar, Y.B. Filamentous fungi associated with mosquito larvae (Diptera: Culicidae) in municipalities of the Brazilian Amazon. Neotrop. Entomol. 2009, 38, 352–359. [Google Scholar] [CrossRef]
- Rashed, S.S.; Helal, G.A.; Rashad, E.M.; Mostafa, W.A. Pathogenicity of entomopathogenic fungi on larvae of Culex pipiens mosquitoes (Diptera: Culicidae). J. Appl. Sci. Res. 2014, 9, 6636–6642. [Google Scholar]
- Maketon, M.; Amnuaykanjanasin, A.; Kaysorngup, A. A rapid knockdown effect of Penicillium citrinum for control of the mosquito Culex quinquefasciatus in Thailand. World J. Microbiol. Biotechnol. 2014, 30, 727–736. [Google Scholar] [CrossRef] [PubMed]
- Mohd Shahwahid, H.O.; Mohd Iqbal, M.N.; Amira Mas Ayu, A.M.; Farah, M.A. Assessing service quality of community-based ecotourism: A case study from Kampung Kuantan Firefly Park. J. Trop. For. Sci. 2013, 25, 22–33. [Google Scholar]
- Wan Jusoh, W.F.A.; Hashim, N.R.; Ibrahim, Z.Z. Firefly distribution and abundance on mangrove vegetation assemblages in Sepetang Estuary, Peninsular Malaysia. Wetl. Ecol. Manag. 2010, 18, 367–373. [Google Scholar] [CrossRef]
- Wan Jusoh, W.F.A.; Hashim, N.R.; Adam, N.A. Distribution of the Synchronous Flashing Beetle, Pteroptyx tener Olivier (Coleoptera: Lampyridae), in Malaysia. Coleopt. Bull. 2013, 67, 604–605. [Google Scholar] [CrossRef]
- Chey, V.K. Fireflies of Beaufort with special reference to Sungai Garama and Sungai Klias. Sepilok Bull. 2010, 12, 13–19. [Google Scholar]
- Mahadimenakbar, M.D.; Fiffy, H.S.; Godoong, E. Studies on the Potential of Fireflies Watching Tourism for Firefly (Coleoptera: Lampyridae; Pteroptyx spp.) Conservation. In Proceedings of the International Seminar on Wetlands & Sustainability; Wetland & Climate Change: The Need for Intergration, Kota Kinabalu, Malaysia, 26–28 June 2009. [Google Scholar]
- Nallakumar, K. The synchronously flashing aggregative fireflies of peninsular Malaysia. Biodiversity 2003, 4, 11–16. [Google Scholar] [CrossRef]
- Pearce-Kelly, P.; Morgan, R.; Honan, P.; Barrett, P.; Perrotti, L.; Magdich, M.; Daniel, B.A.; Sullivan, E.; Veltman, K.; Clarke, D. The conservation value of insect breeding programmes: Rationale, evaluation tools and example programme case studies. In Insect Conservation Biology: Proceedings of the Royal Entomological Society’s 23rd Symposium; Stewart, A.J.A., New, T.R., Lewis, O.T., Eds.; CABI International: Oxfordshire, UK, 2007. [Google Scholar]
- Ballantyne, L.A.; Menayah, R. A description of larvae and redescription of adults of the firefly Pteroptyx valida Olivier in Selangor, Malaysia (Coleoptera: Lampyridae: Luciolinae) with notes on Luciolinae larvae. Raffles Bull. Zool. 2002, 50, 101–109. [Google Scholar]
- Kadowaki, K.; Leschen, R.A.B.; Beggs, J. Spore consumption and life history of Zearagytodes maculifer (Broun) (Coleoptera: Leiodidae) on Ganoderma, its fungal host. N.Z. J. Ecol. 2011, 35, 61–68. [Google Scholar]
- Six, D.L.; Bentz, B.J. Fungi associated with the North American spruce beetle, Dendroctonus Rufipennis. Can. J. For. Res. 2003, 33, 1815–1820. [Google Scholar] [CrossRef]
- David, M.H.; Mills, R.B.; Sauer, D.B. Development and oviposition of Ahasverus advena (Coleoptera: Silvanidae) on seven species of fungi. J. Stored Prod. Res. 1974, 10, 17–22. [Google Scholar] [CrossRef]
- Prabha, K.C.; Sivadasan, R.; Jose, A. Microflora associated with the red flour beetle, Tribolium castaneum (Coleoptera: Tenebrionidae). J. Agric. Technol. 2011, 7, 1625–1631. [Google Scholar]
- Haelewaters, D.; Minnaar, I.A.; Clusella-Trullas, S. First finding of the parasitic fungus Hesperomyces Virescens (Laboulbeniales) on native and invasive ladybirds (Coleoptera: Coccinellidae) in South Africa. Parasite 2016, 23, 5. [Google Scholar] [CrossRef] [PubMed]
- Henriques, J.; Inacio, M.L.; Sousa, E. Fungi associated to Platypus cylindrus Fab. (Coleoptera:Platypodidae) in cork oak. Revista Ciencias Agrarias 2009, 32, 55–56. [Google Scholar]
- Russell, B.M.; Kay, B.H.; Shipton, W. Survival of Aedes aegypti (Diptera:Culicidae) eggs in surface and subterranean breeding sites during the Northern Queensland dry season. J. Med. Entomol. 2001, 38, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Ballantyne, L.A.; Lambkin, C.L. Systematics and phylogenetics of Indo-Pacific Luciolinae Fireflies (Coleoptera: Lampyridae) and the description of new genera. Zootaxa 2013, 3653, 1–162. [Google Scholar] [CrossRef] [PubMed]
- Gouli, V.; Gouli, S.; Marcelino, J.A.P.; Skinner, M.; Parker, B.L. Entomopathogenic fungi associated with exotic invasive insect pests in Northeastern Forest of the USA. Insects 2013, 4, 631–645. [Google Scholar] [CrossRef] [PubMed]
- Houbraken, J.A.M.P.; Frisvad, J.C.; Samson, R.A. Taxonomy of Penicillium citrinum and related species. Fungal Divers. 2010, 44, 117–133. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Harman, G.E. Trichoderma and Gliocladium: Basic biology, Taxonomy and Genetics; Taylor & Francis e-Library: London, UK, 2002; Volume 1. [Google Scholar]
- Bissett, J. A revision of the genus Trichoderma. Section Pachybasium. Can. J. Bot. 1991, 69, 2373–2417. [Google Scholar] [CrossRef]
- Roy, H.E.; Hails, R.S.; Hesketh, H.; Roy, D.B.; Pell, J.K. Beyond biological control: non-pest insects and their pathogens in a changing world. Insect Conserv. Divers. 2009, 2, 65–72. [Google Scholar] [CrossRef]
- Fonseca, Q.R.; de Moura Sarquis, M.I.; Hamada, N.; Alencar, Y.B. Occurrence of filamentous fungi in Simulium goeldii Cerqueira & Nunes de Mello (Diptera: Simuliidae) larvae in central Amazonia, Brazil. Braz. J. Microbiol. 2008, 39, 282–285. [Google Scholar] [PubMed]
- Lewis, S.M.; Cratsley, C.K.; Rooney, J.A. Nuptial Gifts and Sexual Selection in Photinus Fireflies. Integr. Comp. Biol. 2004, 44, 234–237. [Google Scholar] [CrossRef] [PubMed]
- Tillman, P.G. Parasitism and Predation of Stink Bug (Heteroptera: Pentatomidae) Eggs in Georgia Corn Fields. Environ. Entomol. 2010, 39, 1184–1194. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kellner, R.L.L. The role of Microorganisms for eggs and progeny. In Chemoecology of Insect Eggs and Egg Deposition; Hilker, M., Meiners, T., Eds.; Blackwell Publishing: Oxford, UK, 2002; pp. 149–164. [Google Scholar]
- Trougakos, I.P.; Margaritis, L.H. Novel Morphological and Physiological aspects of Insect Eggs. In Chemoecology of Insect Eggs and Egg Deposition; Hilker, M., Meiners, T., Eds.; Blackwell Publishing: Oxford, UK, 2002; pp. 3–31. [Google Scholar]
- Eisner, T.; Rossini, C.; González, A.; Iyenger, V.K.; Siegler, V.S.; Smedley, S.R. Paternal investment in egg defence. In Chemoecology of Insect Eggs and Egg Deposition; Hilker, M., Meiners, T., Eds.; Blackwell Publishing: Oxford, UK, 2002; pp. 91–111. [Google Scholar]
- Gonzalez, A.; Hare, J.F.; Eisner, T. Chemical egg defense in Photuris firefly “femmes fatales”. Chemoecology 1999, 9, 177–185. [Google Scholar] [CrossRef]
- Hosoe, T.; Saito, K.; Ichikawa, M.; Ohba, N. Chemical defense in the firefly, Rhagophthalmus Ohbai (Coleoptera: Rhagophthalmidae). Appl. Entomol. Zool. 2014, 49, 331–335. [Google Scholar] [CrossRef]
- Matsuura, K.; Tamura, T.; Kobayashi, N.; Yashiro, T.; Tatsumi, S. The antibacterial protein lysozyme identified as the termite egg recognition pheromone. PLoS ONE 2007, 2, e813. [Google Scholar] [CrossRef] [PubMed]
- Tragust, S.; Mitteregger, B.; Barone, V.; Konrad, M.; Ugelvig, L.V.; Cremer, S. Ants disinfect fungus-exposed brood by oral uptake and spread of their poison. Curr. Biol. 2013, 23, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Boos, S.; Meunier, J.; Pichon, S.; Kolliker, M. Maternal care provides antifungal protection to eggs in the European earwig. Behav. Ecol. 2014, 25, 754–761. [Google Scholar] [CrossRef]
- Cristensen, J.T.; Sauriau, P.G.; Richard, P.; Jensen, P.D. Diet in mangrove snails: preliminary data on gut contents and stable isotope analysis. J. Shellfish. Res. 2001, 20, 423–426. [Google Scholar]
- Jacobs, C.G.C.; Spaink, H.P.; Zee, M.V.D. The extraembryonic serosa is a frontier epithelium providing the insect egg with a full-range innate immune respone. eLife 2014, 3. [Google Scholar] [CrossRef] [PubMed]
- Shakeri, J.; Foster, H.A. Proteolytic activity and antibiotic production by Trichoderma harzianum in relation to pathogenicity to insects. Enzyme Microb. Technol. 2007, 40, 961–968. [Google Scholar] [CrossRef]
- Ferguson, A.; Pearce-Kelly, P. Captive management of the Frégate Island giant tenebrionid beetle Polposipus herculeanus. Phelsuma 2005, 13, 25–43. [Google Scholar]
- Smith, S.F.; Krischil, V.A. Effects of systemic Imidacloprid on Coleomegilla maculata (Coleoptera: Coccinellidae). Environ. Entomol. 1999, 28, 1189–1195. [Google Scholar] [CrossRef]
- Zha, C.; Cohen, A.C. Effects of anti-fungal compounds on feeding behavior and nutritional ecology of tobacco budworm and painted lady butterfly larvae. Entomol. Ornithol. Herpetol. 2014, 3, 120. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insect Order (Family) | Species | Microfungi | Host Relationships | Reference |
|---|---|---|---|---|
| Coleoptera (Leiodidae) | Zearagytodes maculifer | Ganoderma cf. applanatum & Ganoderma australe | Fungal as food source for larvae | Kadowaki et al., [17] |
| Coleoptera (Scolytidae) | Dendroctonus rufipennis | Leptographium abietinum, Ophiostoma ips, & Ophiostoma piceae | Symbiotic | Six & Bentz, [18] |
| Coleoptera (Silvanidae) | Ahasverus adrena | Aspergillus amstelodam, Penicillium citrinum, & Cladosporium sp. | Fungal as food source for larvae | David et al., [19] |
| Coleoptera (Tenebrionidae) | Tribolium castaneum | Aspergillus flavus, Aspergillus fumigatus, Penicillium spp., Fusarium spp., & Rhizopus oryzae | Symbiotic | Prabha et al., [20] |
| Coleoptera (Coccinellidae) | Harmonia axyridis & Cheilomenes propinquo | Hesperomyces virescens | Parasitic | Haelewaters et al., [21] |
| Coleoptera (Platypodidae) | Platypus cylindrus | Fungus genus: Aspergillus, Paecilomyces, Penicillium, Botrytis, Acremonium, Beauveria, Fusarium, Gliocladium, Trichoderma, Raffaelea, Geotrichum, Chaetomium, Scytalidium, Nodulusporium, & Streptomyces | Symbiotic | Henriques et al., [22] |
| Diptera (Culicidae) | Aedes aegypti | Penicillium citrinum | Entomopathogenic fungal | Russell et al., [23] |
| Diptera (Culicidae) | Culex quinquefasciatus | Penicillium citrinum | Entomopathogenic fungal | Maketon et al., [16] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foo, K.; Sathiya Seelan, J.S.; Dawood, M.M. Microfungi Associated with Pteroptyx bearni (Coleoptera: Lampyridae) Eggs and Larvae from Kawang River, Sabah (Northern Borneo). Insects 2017, 8, 66. https://doi.org/10.3390/insects8030066
Foo K, Sathiya Seelan JS, Dawood MM. Microfungi Associated with Pteroptyx bearni (Coleoptera: Lampyridae) Eggs and Larvae from Kawang River, Sabah (Northern Borneo). Insects. 2017; 8(3):66. https://doi.org/10.3390/insects8030066
Chicago/Turabian StyleFoo, Kevin, Jaya Seelan Sathiya Seelan, and Mahadimenakbar M. Dawood. 2017. "Microfungi Associated with Pteroptyx bearni (Coleoptera: Lampyridae) Eggs and Larvae from Kawang River, Sabah (Northern Borneo)" Insects 8, no. 3: 66. https://doi.org/10.3390/insects8030066
APA StyleFoo, K., Sathiya Seelan, J. S., & Dawood, M. M. (2017). Microfungi Associated with Pteroptyx bearni (Coleoptera: Lampyridae) Eggs and Larvae from Kawang River, Sabah (Northern Borneo). Insects, 8(3), 66. https://doi.org/10.3390/insects8030066