
Isolation and Characterization of Microsatellite Loci for Cotesia plutellae (Hymenoptera: Braconidae)

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Primer Design
2.2. Proved the Effectiveness of Primers
2.3. Data Analysis
3. Results and Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Furlong, M.J.; Wright, D.J.; Dosdall, M. Diamondback moth ecology and management: problems, progress, and prospects. Annu. Rev. Entomol. 2013, 58, 517–541. [Google Scholar] [CrossRef] [PubMed]
- Delvare, G.; Kirk, A.A.; Bordet, D. The taxonomic status and role of Hymenoptera in biological control of DBM, Plutella xylostella (L.) (Lepidoptera: Plutellidae) Improving biocontrol of Plutella xylostella. In Proceedings of the International Symposium on CIRAD, Montpellier, France, 22–24 October 2004; pp. 17–49. [Google Scholar]
- Shi, Z.H.; Liu, S.S.; Li, Y.X. Cotesia plutellae parasitizing Plutella xylostella: Host-age dependent parasitism and its effect on host development and food consumption. Biocontrol 2002, 47, 499–511. [Google Scholar] [CrossRef]
- Ali, M.R.; Seo, J.; Lee, D.; Kim, Y. Teratocyte-secreting proteins of an endoparasitoid wasp, Cotesia plutellae, prevent host metamorphosis by altering endocrine signals. Comp. Biochem. Phys. 2013, 166, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Hepat, R.; Lee, D.; Kim, Y. Protein tyrosine phosphatase encoded in Cotesia plutellae bracovirus suppresses a larva-to-pupa metamorphosis of the diamondback moth, Plutella xylostella. Comp. Biochem. Phys. 2013, 166, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Gariepy, T.D.; Kuhlmann, U.; Gillott, C.; Erlandson, M. Parasitoids, predators and PCR: The use of diagnostic molecular markers in biological control of Arthropods. J. Appl. Entomol. 2007, 131, 225–240. [Google Scholar] [CrossRef]
- Selkoe, K.A.; Toonen, R.J. Microsatellites for ecologists: A practical guide to using and evaluating microsatellite markers. Ecol. Lett. 2006, 9, 615–629. [Google Scholar] [CrossRef] [PubMed]
- Vaiman, D.; Pailhoux, E.; Payen, E.; Saidi-Mehtar, N.; Cotinot, C. Evolutionary conservation of a microsatellite in the Wilms Tumour (WT) gene: Mapping in sheep and cattle. Cytogenet. Genome. Res. 1995, 70, 112–115. [Google Scholar] [CrossRef]
- Roderick, G.K.; Navajas, M. Genes in new environments: Genetics and evolution in biological control. Nat. Rev. Genet. 2003, 4, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Cullingham, C.I.; Roe, A.D.; Sperling, F.A.H.; Coltman, D.W. Phylogeographic insights into an irruptive pest outbreak. Ecol. Evol. 2012, 2, 908–919. [Google Scholar] [CrossRef] [PubMed]
- Thiel, T.; Michalek, W.; Varshney, R.K.; Graner, A. Exploiting EST databases for the development and characterization of gene-derived SSR-markers in barley (Hordeum Vulgare L.). Theor. Appl. Genet. 2003, 106, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, O.; Brillada, C.; Yazawa, S.; Maffei, M.E.; Arimura, G.I. Transcriptome pyrosequencing of the parasitoid wasp Cotesia plutellae: genes involved in the antennal odorant-sensory system. PLoS ONE 2012, 7, e50664. [Google Scholar] [CrossRef] [PubMed]
- Rozen, S.; Skaletsky, H. Primer 3. Available online: http://primer3.sourceforge.net/ (accessed on 6 September 2015).
- Cook, J.M. Sex determination in the Hymenoptera: A review of models and evidence. Heredity 1993, 71, 421–435. [Google Scholar] [CrossRef]
- Heimpel, G.E.; De Boer, J.G. Sex determination in the Hymenoptera. Annu. Rev. Entomol. 2008, 53, 209–230. [Google Scholar] [CrossRef] [PubMed]
- Rousset, F. Genepop’007: A complete re-implementation of the genepop software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Tian, E.W.; Yu, H.; Zhang, D.Y.; Nason, J.D. Development of microsatellite loci for Blastophaga javana (Agaonidae), the pollinating wasp of Ficus hirta (Moraceae). Am. J. Bot. 2011, 98, E41–E43. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.C.; Bergamaschi, A.B.; Sanches, A.; Hatanaka, T.; Del Lama, M.A. Isolation and characterization of microsatellite loci for the mud-dauber wasp Trypoxylon (Trypargilum) albitarse (Hymenoptera: Crabronidae). Eur. J. Entomol. 2013, 110, 541–543. [Google Scholar] [CrossRef]
- Chen, W.; Fang, L.; Liu, J.; He, Z.; Hu, H. Isolation and characterization of polymorphic microsatellite loci for Pachycrepoideus vindemmiae (Rondani) (Hymenoptera: Pteromalidae). Genet. Mol. Res. 2015, 14, 1798–1801. [Google Scholar] [CrossRef] [PubMed]
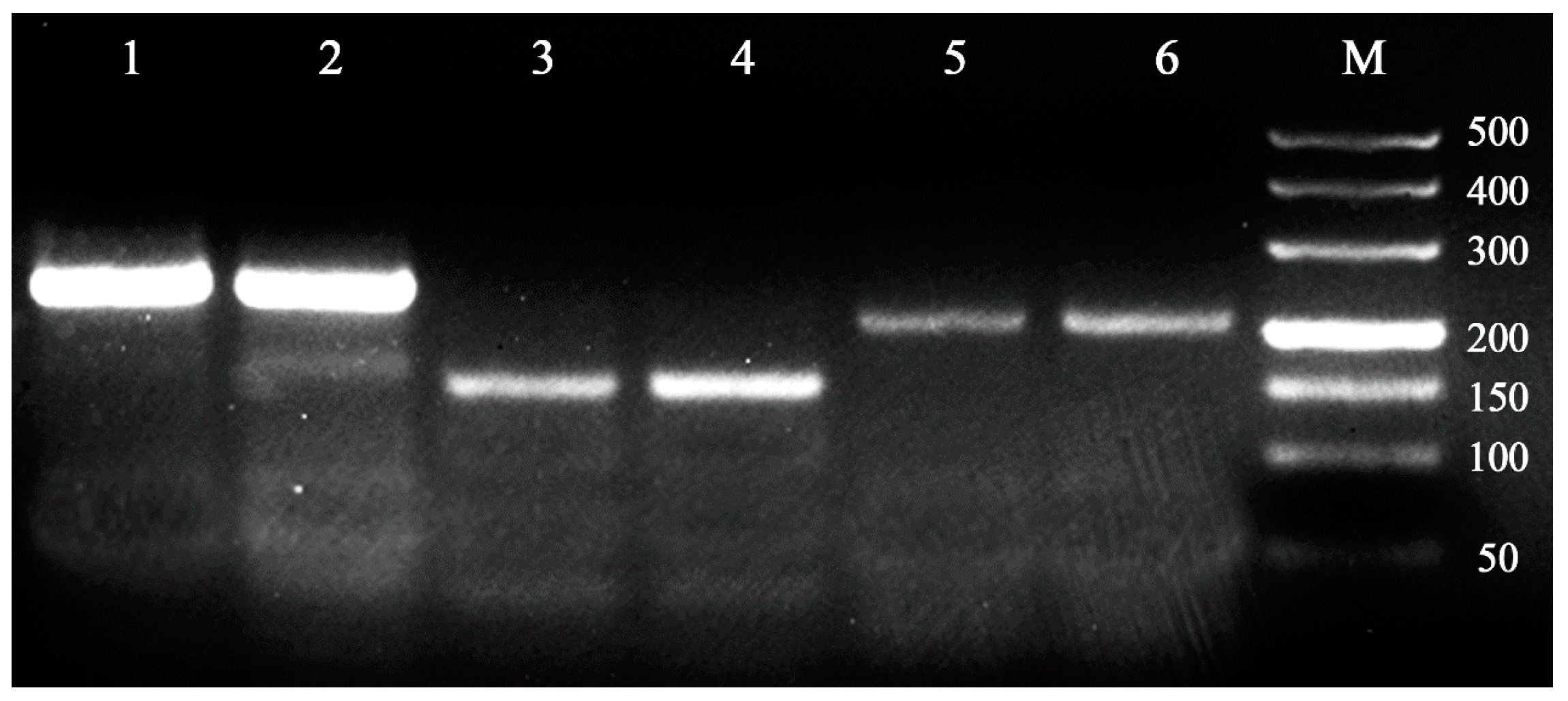

{kind=link}
| Locus | Repeat Motif | Primer Sequences (5’–3’) | Tm (°C) | Na | Size Range (bp) | HO/HE | Fis | Annotation |
|---|---|---|---|---|---|---|---|---|
| C6 | CTG | F: AGAGCGGCAGTATCGTGAGT R: AGGAAAAGTCCTCAGCCTCC | 53 | 5 | 245–260 | 0.250/0.316 | 0.2114 * | Putative uncharacterized protein |
| C19 | AAT | F: CGCGAAAGAACGAATTTGAG R: TCACAGTATACGTCATTCCCAAG | 54 | 5 | 129–141 | 0.141/0.129 | −0.0941 | Bifunctional protein FolD |
| C21 | AAT | F: TCGCTAGAAAAAGTTTCGGC R: AATGAAGCAGGGTGAAATGC | 53 | 4 | 232–241 | 0.172/0.129 | 0.1852 | Putative uncharacterized protein |
| C22 | CTG | F: CGCGACTCTCTGGCTCTACT R: TCAGGAGTCAGGAGTGGCTT | 56 | 3 | 155–161 | 0.203/0.234 | 0.1352 | cAMP responsive element-binding protein-like 2 |
| C31 | GAA | F: AAAACGTGACCAAAAGCTGG R: GGCCCGAGTACAAACAACTC | 55 | 2 | 215–218 | 0.141/0.123 | −0.1482 | Putative uncharacterized protein |
| C32 | CTG | F: TATGGGCGATAAAGGTGCTC R: AGGAAAAGTCCTCAGCCTCC | 55 | 4 | 291–300 | 0.218/0.279 | 0.2202 * | Ceramide kinase |
| C51 | TAT | F: AAAGGACGGGATAGATCGGT R: ACACTCAGGAATCCCACGAC | 53 | 2 | 296–299 | 0.172/0.164 | −0.0460 | Putative uncharacterized protein |
| C53 | TAT | F: GGCGAATTGGTTATGCTGAT R: TCGAAACATTGAGACAGCGT | 53 | 6 | 210–257 | 0.188/0.276 | 0.3243 * | Putative uncharacterized protein |
| C54 | TAT | F: TATCCTCTTCGCGCGTTATT R: AGGAACTCGTTTCCAACAGC | 53 | 4 | 188–197 | 0.188/0.273 | 0.4572 | Putative uncharacterized protein |
| C57 | TCG | F: CCGGAACTGTTTTGTCACG R: CCGGAGTACGCTCTCAAGAC | 52 | 4 | 124–133 | 0.250/0.266 | 0.0615 | Putative uncharacterized protein |
| gi11 | TTC | F: TTAATATAAACTGGCGGCGG R: CTCGGTCGACCAATGAAAAT | 53 | 2 | 216–219 | 0.189/0.218 | 0.1429 | Putative uncharacterized protein |
| gi16 | TCT | F: TCCACTGCAAGCCATACAAG R: TGGTGATGTTGAGAAACCGA | 53 | 2 | 126–129 | 0.203/0.248 | 0.1826 | Putative uncharacterized protein |
| gi27 | TAC | F: TGGATTTGCCACTACCATCA R: GTTGAAAGGGCCAATTTTGA | 53 | 3 | 194–200 | 0.219/0.290 | 0.2485 | Putative uncharacterized protein |
| gi29 | TAG | F: TTAGTGGCGGCAGTGATAAT R: CCGTGTAACAAACCCCTGAT | 53 | 3 | 244–250 | 0.281/0.298 | 0.0558 | Putative uncharacterized protein |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, T.; Ke, F.; You, S.; Chen, W.; He, W.; You, M. Isolation and Characterization of Microsatellite Loci for Cotesia plutellae (Hymenoptera: Braconidae). Insects 2017, 8, 63. https://doi.org/10.3390/insects8020063
Liu T, Ke F, You S, Chen W, He W, You M. Isolation and Characterization of Microsatellite Loci for Cotesia plutellae (Hymenoptera: Braconidae). Insects. 2017; 8(2):63. https://doi.org/10.3390/insects8020063
Chicago/Turabian StyleLiu, Tiansheng, Fushi Ke, Shijun You, Wenbin Chen, Weiyi He, and Minsheng You. 2017. "Isolation and Characterization of Microsatellite Loci for Cotesia plutellae (Hymenoptera: Braconidae)" Insects 8, no. 2: 63. https://doi.org/10.3390/insects8020063
APA StyleLiu, T., Ke, F., You, S., Chen, W., He, W., & You, M. (2017). Isolation and Characterization of Microsatellite Loci for Cotesia plutellae (Hymenoptera: Braconidae). Insects, 8(2), 63. https://doi.org/10.3390/insects8020063