
Insect Pathogenic Bacteria in Integrated Pest Management


Abstract
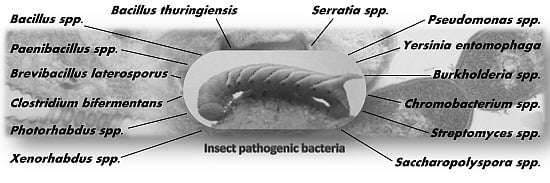
:
{kind=link}
1. Introduction
2. Insect Pathogenic Bacteria
2.1. Bacillaceae
2.1.1. Bacillus thuringiensis
2.1.2. Lysinibacillus sphaericus
2.1.3. Paenibacillus spp.
2.1.4. Brevibacillus laterosporus
2.2. Clostridiaceae
Clostridium bifermentans
2.3. Gammaproteobacteria
2.3.1. Photorhabdus spp. and Xenorhabdus spp.
2.3.2. Serratia spp.
2.3.3. Yersinia entomophaga
2.3.4. Pseudomonas entomophila
2.4. Betaproteobacteria
2.4.1. Burkholderia spp.
2.4.2. Chromobacterium spp.
2.5. Actinobacteria
2.5.1. Streptomyces spp.
2.5.2. Saccharopolyspora spinosa
3. Use of Entomopathogenic Bacteria in Integrated Pest Management
4. Market Overview
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Feldhaar, H. Bacterial symbionts as mediators of ecologically important traits of insect hosts. Ecol. Entomol. 2011, 36, 533–543. [Google Scholar] [CrossRef]
- Vilcinskas, A. Coevolution between pathogen-derived proteinases and proteinase inhibitors of host insects. Virulence 2010, 1, 206–214. [Google Scholar] [CrossRef] [PubMed]
- De Maagd, R.A.; Bravo, A.; Berry, C.; Crickmore, N.; Schnepf, H.E. Structure, diversity, and evolution of protein toxins from spore forming entomopathogenic bacteria. Annu. Rev. Genet. 2003, 37, 409–433. [Google Scholar] [CrossRef] [PubMed]
- Ruiu, L.; Satta, A.; Floris, I. Emerging entomopathogenic bacteria for insect pest management. Bull. Insectol. 2013, 66, 181–186. [Google Scholar]
- Lacey, L.A.; Frutos, R.; Kaya, H.K.; Vail, P. Insect pathogens as biological control agents: Do they have a future? Biol. Control 2001, 21, 230–248. [Google Scholar] [CrossRef]
- Glare, T.; Caradus, J.; Gelernter, W.; Jackson, T.; Keyhani, N.; Kohl, J.; Marrone, P.; Morin, L.; Stewart, A. Have biopesticides come of age? Trends Biotechnol. 2012, 30, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Vega, F.E.; Kaya, H.K. Insect Pathology, 2nd ed.; Elsevier: London, UK, 2012; p. 504. [Google Scholar]
- Pigott, C.R.; Ellar, D.J. Role of receptors in Bacillus thuringiensis crystal toxin activity. Microbiol. Mol. Biol. Rev. 2007, 71, 255–281. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A.; Gill, S.S.; Soberon, M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 2007, 49, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Glare, T.R.; O’Callaghan, M. Bacillus thuringiensis: Biology, Ecology and Safety; Wiley: Chichester, UK, 2000. [Google Scholar]
- Ruiu, L.; Falchi, G.; Floris, I.; Marche, M.G.; Mura, M.E.; Satta, A. Pathogenicity and characterization of a novel Bacillus cereus sensu lato isolate toxic to the Mediterranean fruit fly Ceratitis capitata Wied. J. Invertebr. Pathol. 2015. [Google Scholar] [CrossRef]
- Baumann, P.; Clark, M.A.; Baumann, L.; Broadwell, A.H. Bacillus sphaericus as a mosquito pathogen: Properties of the organism and its toxins. Microbiol. Rev. 1991, 55, 425–436. [Google Scholar] [PubMed]
- Charles, J.F.; Silva-Filha, M.H.; Nielsen-LeRoux, C. Mode of action of Bacillus sphaericus on mosquito larvae: Incidence on resistance. In Entomopathogenic Bacteria: From Laboratory to Field Application; Charles, J.F., Delecluse, A., Nielsen-LeRoux, C., Eds.; Kluwer Academic Publishers: London, UK, 2000; pp. 237–252. [Google Scholar]
- Davidson, E.A. Ultrastructures of American foulbrood disease pathogenesis in larvae of the worker honey bee Apis mellifera. J. Invertebr. Pathol. 1973, 21, 53–61. [Google Scholar] [CrossRef]
- Zhang, J.; Hodgman, T.C.; Krieger, L.; Schnetter, W.; Schairer, H.U. Cloning and analysis of the cry gene from Bacillus popilliae. J. Bacteriol. 1997, 179, 4336–4341. [Google Scholar]
- Ruiu, L. Brevibacillus laterosporus, a pathogen of invertebrates and a broad-spectrum antimicrobial species. Insects 2013, 4, 476–492. [Google Scholar] [CrossRef]
- Djukic, M.; Poehlein, A.; Thürmer, A.; Daniel, R. Genome sequence of Brevibacillus laterosporus LMG 15441, a pathogen of invertebrates. J. Bacteriol. 2011, 193, 5535–5536. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Singh, P.K.; Midha, S.; Ranjan, M.; Korpole, S.; Patil, P.B. Genome sequence of Brevibacillus laterosporus strain GI-9. J. Bacteriol. 2012. [Google Scholar] [CrossRef]
- Warren, G.W. Vegetative insecticidal proteins: Novel proteins for control of corn pests. In Advances in Insect Control: The Role of Transgenic Plants; Carozzi, N.B., Koziel, M.G., Eds.; Taylor & Francis: London, UK, 1997; pp. 109–121. [Google Scholar]
- Zubasheva, M.V.; Ganushkina, L.A.; Smirnova, T.A.; Azizbekyan, R.R. Larvicidal activity of crystal-forming strains of Brevibacillus laterosporus. Appl. Biochem. Microbiol. 2010, 46, 755–762. [Google Scholar] [CrossRef]
- Ruiu, L.; Floris, I.; Satta, A.; Ellar, D.J. Toxicity of a Brevibacillus laterosporus strain lacking parasporal crystals against Musca domestica and Aedes aegypti. Biol. Control 2007, 43, 136–143. [Google Scholar] [CrossRef]
- Ruiu, L.; Satta, A.; Floris, I. Observations on house fly larvae midgut ultrastructure after Brevibacillus laterosporus ingestion. J. Invertebr. Pathol. 2012, 111, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, L.; Hamon, S.; Frachon, E.; Sebald, M.; de Barjac, H. Partial inactivation of the mosquitocidal activity of Clostridium bifermentans serovar malaysia by extracellular proteinases. Appl. Microbiol. Biotechnol. 1990, 34, 36–41. [Google Scholar]
- Barloy, F.; Delècluse, A.; Nicolas, L.; Lecadet, M.M. Cloning and Expression of the first anaerobic toxin gene from Clostridium bifermentans subsp. malaysia encoding a new mosquitocidal protein with homologies to Bacillus thuringiensis delta-endotoxins. J. Bacteriol. 1996, 178, 3099–3105. [Google Scholar]
- Waterfield, N.R.; Bowen, D.J.; Fetherston, J.D.; Perry, R.D.; Ffrench-Constant, R.H. The TC genes of Photorhabdus: A growing family. Trends Microbiol. 2001, 9, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Ffrench-Constant, R.; Waterfield, N. An ABC guide to the bacterial toxin complexes. Adv. Appl. Microbiol. 2006, 58, 169–183. [Google Scholar] [PubMed]
- Ffrench-Constant, R.H.; Dowling, A.; Waterfield, N.R. Insecticidal toxins from Photorhabdus bacteria and their potential use in agriculture. Toxicon 2007, 49, 436–451. [Google Scholar] [CrossRef] [PubMed]
- Waterfield, N.; Kamita, S.G.; Hammock, B.D.; Ffrench-Constant, R. The Photorhabdus Pir toxins are similar to a developmentally regulated insect protein but show no juvenile hormone esterase activity. FEMS Microbiol. Lett. 2005, 245, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Eleftherianos, I.; Millichap, P.J.; Ffrench-Constant, R.H.; Reynolds, S.E. RNAi suppression of recognition protein mediated immune responses in the tobacco hornworm Manduca sexta causes increased susceptibility to the insect pathogen Photorhabdus. Dev. Comparat. Immunol. 2006, 30, 1099–1107. [Google Scholar] [CrossRef]
- Ji, D.; Kim, Y. An entomopathogenic bacterium, Xenorhabdus nematophila, inhibits the expression of an antimicrobial peptide, cecropin, of the beet armyworm, Spodoptera exigua. J. Insect Physiol. 2004, 50, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Tailliez, P.; Pagès, S.; Ginibre, N.; Boemare, N. New insight into diversity in the genus Xenorhabdus, including the description of ten novel species. Int. J. Syst. Evolut. Microbiol. 2006, 56, 2805–2818. [Google Scholar]
- Zhang, C.-X.; Yang, S.-Y.; Xu, M.-X.; Sun, J.; Liu, H.; Liu, J.R.; Liu, H.; Kan, F.; Sun, J.; Lai, R.; et al. Serratia nematodiphila sp. nov., associated symbiotically with the entomopathogenic nematode Heterorhabditidoides chongmingensis (Rhabditida: Rhabditidae). Int. J. Syst. Evolut. Microbiol. 2000, 59, 1603–1608. [Google Scholar]
- Abebe, E.; Abebe-Akele, F.; Morrison, J. An insect pathogenic symbiosis between a Caenorhabditis and Serratia. Virulence 2011, 2, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Torres-Barragan, A.; Suazo, A.; Buhler, W.; Cardoza, Y. Studies on the entomopathogenicity and bacterial associates of the nematode Oscheius carolinensis. Biol. Control 2011, 59, 123–129. [Google Scholar] [CrossRef]
- Castagnola, A.; Stock, S.P. Common virulence factors and tissue targets of entomopathogenic bacterial for biological control of Lepidopteran pests. Insects 2014, 5, 139–166. [Google Scholar] [CrossRef] [PubMed]
- Jackson, T.A.; Pearson, J.F.; O’Callaghan, M.; Mahanty, H.K.; Willocks, M. Pathogen to product development of Serratia entomophila Enterobacteriaceae as a commercial biological control agent for the New Zealand grass grub Costelytra zealandica. In Use of Pathogens in Scarab Pest Management; Jackson, T.A., Glare, T.R., Eds.; Intercept Ltd.: Andover, UK, 1992; pp. 191–198. [Google Scholar]
- Hurst, M.R.; Glare, T.R.; Jackson, T.A.; Ronson, C.W. Plasmid-located pathogenicity determinants of Serratia entomophila, the causal agent of amber disease of grass grub, show similarity to the insecticidal toxins of Photorhabdus luminescens. J. Bacteriol. 2000, 182, 5127–5138. [Google Scholar] [CrossRef] [PubMed]
- Kwak, Y.; Khan, A.R.; Shin, J.H. Genome sequence of Serratia nematodiphila DSM 21420(T), a symbiotic bacterium from entomopathogenic nematode. J. Biotechnol. 2015, 193, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Ishii, K.; Adachi, T.; Hamamoto, H.; Sekimizu, K. Serratia marcescens suppresses host cellular immunity via the production of an adhesion-inhibitory factor against immunosurveillance cells. J. Biol. Chem. 2014, 289, 5876–5888. [Google Scholar] [CrossRef] [PubMed]
- Hurst, M.R.H.; Jones, S.A.; Binglin, T.; Harper, L.A.; Jackson, T.A.; Glare, T.R. The main virulence determinant of Yersinia entomophaga MH96 is a broad-host-range toxin complex active against insects. J. Bacteriol. 2011, 193, 1966–1980. [Google Scholar] [CrossRef] [PubMed]
- Landsberg, M.J.; Jones, S.A.; Rothnagel, R.; Busby, J.N.; Marshall, S.D.G.; Simpson, R.M.; Lott, J.S.; Hankamer, M.; Hurst, M.R.H. 3D structure of the Yersinia entomophaga toxin complex and implications for insecticidal activity. Proc. Natl. Acad. Sci. USA 2011, 108, 20544–20549. [Google Scholar]
- Marshall, S.D.G.; Hares, M.C.; Jones, S.A.; Harper, L.A.; Vernon, J.R.; Harland, D.P.; Jackson, T.A.; Hurst, M.R.H. Histopathological effects of the Yen-Tc toxin complex from Yersinia entomophaga MH96 (Enterobacteriaceae) on the Costelytra zealandica (Coleoptera: Scarabaeidae) larval midgut. Appl. Environ. Microbiol. 2012, 78, 4835–4847. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, C.M.; Barton, D.M.; Harpe, L.A.; Swaminathan, J.; van Koten, C.; Hurst, M.R.H. Survival of Yersinia entomophaga MH96 in a pasture ecosystem and effects on pest and non-target invertebrate populations. New Zealand Plant Prot. 2012, 65, 166–173. [Google Scholar]
- Vodovar, N.; Vinals, M.; Liehl, P.; Basset, A.; Degrouard, J.; Spellman, P.; Boccard, F.; Lemaitre, B. Drosophila host defense after oral infection by and entomopathogenic Pseudomonas species. Proc. Natl. Acad. Sci. USA 2005, 102, 11414–11419. [Google Scholar] [CrossRef] [PubMed]
- Vodovar, N.; Vallenet, D.; Cruveiller, S.; Rouy, Z.; Barbe, V.; Acosta, C.; Cattolico, L.; Jubin, C.; Lajus, A.; Segurens, B.; et al. Complete genome sequence of the entomopathogenic and metabolically versatile soil bacterium Pseudomonas entomophila. Nat. Biotechnol. 2006, 24, 673–679. [Google Scholar]
- Kim, J.K.; Kim, N.H.; Jang, H.A.; Kikuchi, Y.; Kim, C.H.; Fukatsu, T.; Lee, B.L. Specific midgut region controlling the symbiont population in an insect-microbe gut symbiotic association. Appl. Environ. Microbiol. 2013, 79, 7229–7233. [Google Scholar] [CrossRef] [PubMed]
- Martinson, V.G.; Danforth, B.N.; Minckley, R.L.; Rueppell, O.; Tingek, S.; Moran, N.A. A simple and distinctive microbiota associated with honey bees and bumble bees. Mol. Ecol. 2011, 20, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Kil, Y.J.; Seo, M.J.; Kang, D.K.; Oh, S.N.; Cho, H.S.; Youn, Y.N.; Yasunaga-Aoki, C.; Yu, Y.M. Effects of Enterobacteria (Burkholderia sp.) on development of Riptortus pedestris. J. Fac. Agric. Kyushu Univ. 2014, 59, 77–84. [Google Scholar]
- Burkhead, K.D.; Schisler, D.A.; Slininger, P.J. Pyrrolnitrin production by biological-control agent Pseudomonas cepacia B37W in culture and in colonized wounds of potatoes. Appl. Environ. Microbiol. 1994, 60, 2031–2039. [Google Scholar] [PubMed]
- Janisiewicz, W.J.; Roitman, J. Biological control of blue mold and gray mold on apple and pear with Pseudomonas cepacia. Phytopathology 1988, 78, 1697–1700. [Google Scholar] [CrossRef]
- Cordova-Kreylos, A.L.; Fernandez, L.E.; Koivunen, M.; Yang, A.; Flor-Weiler, L.; Marrone, P.G. Isolation and characterization of Burkholderia rinojensis sp. nov., a non-Burkholderia cepacia complex soil bacterium with insecticidal and miticidal activities. Appl. Environ. Microbiol. 2013, 79, 7669–7678. [Google Scholar]
- He, H.; Ratnayake, A.S.; Janso, J.E.; He, M.; Yang, H.Y.; Loganzo, F.; Shor, B.; O’Donnell, C.J.; Koehn, F.E. Cytotoxic spliceostatins from Burkholderia sp. and their semisynthetic analogues. J. Nat. Prod. 2014, 77, 1864–1870. [Google Scholar] [CrossRef]
- Martin, P.A.W.; Hirose, E.; Aldrich, J.R. Toxicity of Chromobacterium subtsugae to Southern stink bug (Heteroptera: Pentatomidae) and corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2007, 100, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.A.W.; Gundersen-Rindal, D.; Blackburn, M.; Buyer, J. Chromobacterium subtsugae sp. nov., a betaproteobacterium toxic to Colorado potato beetle and other insect pests. Int. J. Syst. Evolut. Microbiol. 2007, 57, 993–999. [Google Scholar]
- Asolkar, R.; Huang, H.; Koivunen, M.; Marrone, P. Chromobacterium Bioactive Compositions and Metabolites. US Patent Application Publication, US 14/293,728, 9 October 2014. [Google Scholar]
- Koivunen, M.; Chanbusarakum, L.; Fernandez, L.; Asolkar, R.; Tan, E.; Wallner, D.; Marrone, P. Development of a new microbial insecticide based on Chromobacterium subtsugae. IOBC/wprs Bull. 2009, 45, 183–186. [Google Scholar]
- Martin, P.A.W.; Shropshire, A.D.S.; Gundersen-Rindal, D.E.; Blackburn, M.B. Chromobacterium subtsugae sp. nov. and Use for Control of Insect Pests. U.S. Patent Application Pubblication, 2007/0172463 A1, 26 July 2007. [Google Scholar]
- Ramirez, J.L.; Short, S.M.; Bahia, A.C.; Saraiva, R.G.; Dong, Y.; Kang, S.; Tripathi, A.; Mlambo, G.; Dimopoulos, G. Chromobacterium Csp_P reduces malaria and dengue infection in vector mosquitoes and has entomopathogenic and in vitro anti-pathogen activities. PLOS Pathog. 2014, 10, e1004398. [Google Scholar] [CrossRef] [PubMed]
- Book, A.J.; Lewin, G.R.; McDonald, B.R.; Takasuka, T.E.; Doering, D.T.; Adams, A.S.; Blodgett, J.A.V.; Clardy, J.; Raffa, K.F.; Fox, B.G.; et al. Cellulolytic Streptomyces strains associated with herbivorous insects share a phylogenetically linked capacity to degrade lignocelluloses. Appl. Environ. Microbiol. 2014, 80, 4692–4701. [Google Scholar]
- Copping, G.L.; Menn, J.J. Biopesticides: A review of their action, applications and efficacy. Pest Manag. Sci. 2000, 56, 651–676. [Google Scholar] [CrossRef]
- Craveri, R.; Giolitti, G. An antibiotic with fungicidal and insecticidal activity produced by Streptomyces. Nature 1957, 179, 1307. [Google Scholar] [CrossRef] [PubMed]
- Kido, G.S.; Spyhalski, E. Antimycin A, an antibiotic with insecticidal and miticidal properties. Science 1950, 112, 172–173. [Google Scholar] [CrossRef] [PubMed]
- Takahaski, N.; Suzuki, A.; Kimura, Y.; Miyamoto, S.; Tamura, S.; Mitsui, T.; Fukami, J. Isolation, structure and physiological activities of piericidin B, natural insecticide produced by a Streptomyces. Agric. Biol. Chem. 1968, 32, 1115–1122. [Google Scholar] [CrossRef]
- Oishi, H.; Sugawa, T.; Okutomi, T.; Suzuki, K.; Hayashi, T.; Sawada, M.; Ando, K. Insecticidal activity of macrotetrolide antibiotics. J. Antibiot. 1970, 23, 105–106. [Google Scholar] [CrossRef] [PubMed]
- Box, S.J.; Cole, M.; Yeoman, G.H. Prasinons A and B: Potent insecticides from Streptomyces prasinus. Appl. Microbiol. 1973, 29, 699–704. [Google Scholar]
- Turner, M.J.; Schaeffer, J.M. Mode of action of ivermectin. In Ivermectin and Abamectin; Cambell, W.C., Ed.; Springer-Verlag: New York, NY, USA, 1989; pp. 73–88. [Google Scholar]
- Bloomquist, J.R. Ion Channels as Targets for Insecticides. Annu. Rev. Entomol. 1996, 41, 163–190. [Google Scholar] [CrossRef] [PubMed]
- Mertz, F.P.; Yao, R.C. Saccharopolyspora spinosa sp. nov. isolated from soil collected in a sugar mill rum still. Int. J. Syst. Bacteriol. 1990, 40, 34–39. [Google Scholar]
- Kirst, H.A.; Michel, K.H.; Martin, J.W.; Creemer, L.C.; Chio, E.H.; Yao, R.C.; Nakatsukasa, W.M.; Boeck, L.D.; Occolowitz, J.L.; Paschal, J.W.; et al. A83543A-D, unique fermentation-derived tetracyclic macrolides. Tetrahedron Lett. 1991, 32, 4839–4842. [Google Scholar]
- Waldron, C.; Madduri, K.; Crawford, K.; Merlo, D.J.; Treadway, P.; Broughton, M.C.; Baltz, R.H. A cluster of genes for the biosynthesis of spinosyns, novel macrolide insect control agents produced by Saccharopolyspora spinosa. Antonie Leeuwenhoek 2000, 78, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Kirst, H.A. The spinosyn family of insecticides: Realizing the potential of natural products research. J. Antibiot. 2010, 63, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Salgado, V.L.; Sparks, T.C. The spinosyns: Chemistry, biochemistry, mode of action, and resistance. In Comprehensive Molecular Insect Science; Gilbert, L.J., Iatrou, K., Gill, S.S., Eds.; Elsevier: Oxford, UK, 2005; Volume 6, pp. 137–173. [Google Scholar]
- Perry, T.; McKenzie, J.A.; Batterham, P.A. Da6 knockout strain of Drosophila melanogaster confers a high level of resistance to spinosad. Insect Biochem. Mol. Biol. 2007, 37, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Sparks, T.C.; Crouse, G.D.; Durst, G. Natural products as insecticides: The biology, biochemistry and quantitative structure-activity relationships of spinosyns and spinosoids. Pest Manag. Sci. 2001, 57, 896–905. [Google Scholar] [CrossRef] [PubMed]
- Lewer, P.; Hahn, D.R.; Karr, L.L.; Duebelbeis, D.O.; Gilbert, J.R.; Crouse, G.D.; Worden, T.; Sparks, T.C.; McKamey, P.; Edwards, R.; et al. Discovery of the butenyl-spinosyn insecticides: Novel macrolides from the new bacterial strain Saccharopolyspora pogona. Bioorg. Med. Chem. 2009, 17, 4185–4196. [Google Scholar]
- Morris, O.N. Susceptibility of some forest insects to mixtures of commercial Bacillus thuringiensis and chemical insecticides, and sensitivity of the pathogen to the insecticides. Can. Entomol. 1972, 104, 1419–1425. [Google Scholar] [CrossRef]
- Seleena, P.; Lee, H.L.; Chiang, Y.F. Compatibility of Bacillus thuringiensis serovar israelensis and chemical insecticides for the control of Aedes mosquitoes. J. Vector Ecol. 1999, 24, 216–223. [Google Scholar] [PubMed]
- Musser, F.R.; Nyrop, J.P.; Shelton, A.M. Integrating biological and chemical controls in decision making: European corn borer (Lepidoptera: Crambidae) control in sweet corn as an example. J. Econ. Entomol. 2006, 99, 1538–1549. [Google Scholar] [CrossRef] [PubMed]
- Satinder, K.B.; Verma, M.; Tyagi, R.D.; Valéro, J.R. Recent advances in downstream processing and formulations of Bacillus thuringiensis based biopesticides. Process Biochem. 2006, 41, 323–342. [Google Scholar] [CrossRef]
- Oerke, E.C.; Dehne, H.W. Safeguarding production losses in major crops and the role of crop protection. Crop Prot. 2004, 23, 275–285. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiu, L. Insect Pathogenic Bacteria in Integrated Pest Management. Insects 2015, 6, 352-367. https://doi.org/10.3390/insects6020352
Ruiu L. Insect Pathogenic Bacteria in Integrated Pest Management. Insects. 2015; 6(2):352-367. https://doi.org/10.3390/insects6020352
Chicago/Turabian StyleRuiu, Luca. 2015. "Insect Pathogenic Bacteria in Integrated Pest Management" Insects 6, no. 2: 352-367. https://doi.org/10.3390/insects6020352
APA StyleRuiu, L. (2015). Insect Pathogenic Bacteria in Integrated Pest Management. Insects, 6(2), 352-367. https://doi.org/10.3390/insects6020352