
Identification of Oral Secretion Proteins in Ostrinia furnacalis by Transcriptome and LC-MS/MS Analyses


Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Biological Materials
2.2. Transcriptome Sequencing
2.3. Oral Secretions Collection, Protein Extraction and Digestion
2.4. Nano-LC-MS/MS Analysis
2.5. Bioinformatics Analysis
2.6. Quantitative PCR (qPCR) Analysis
3. Results
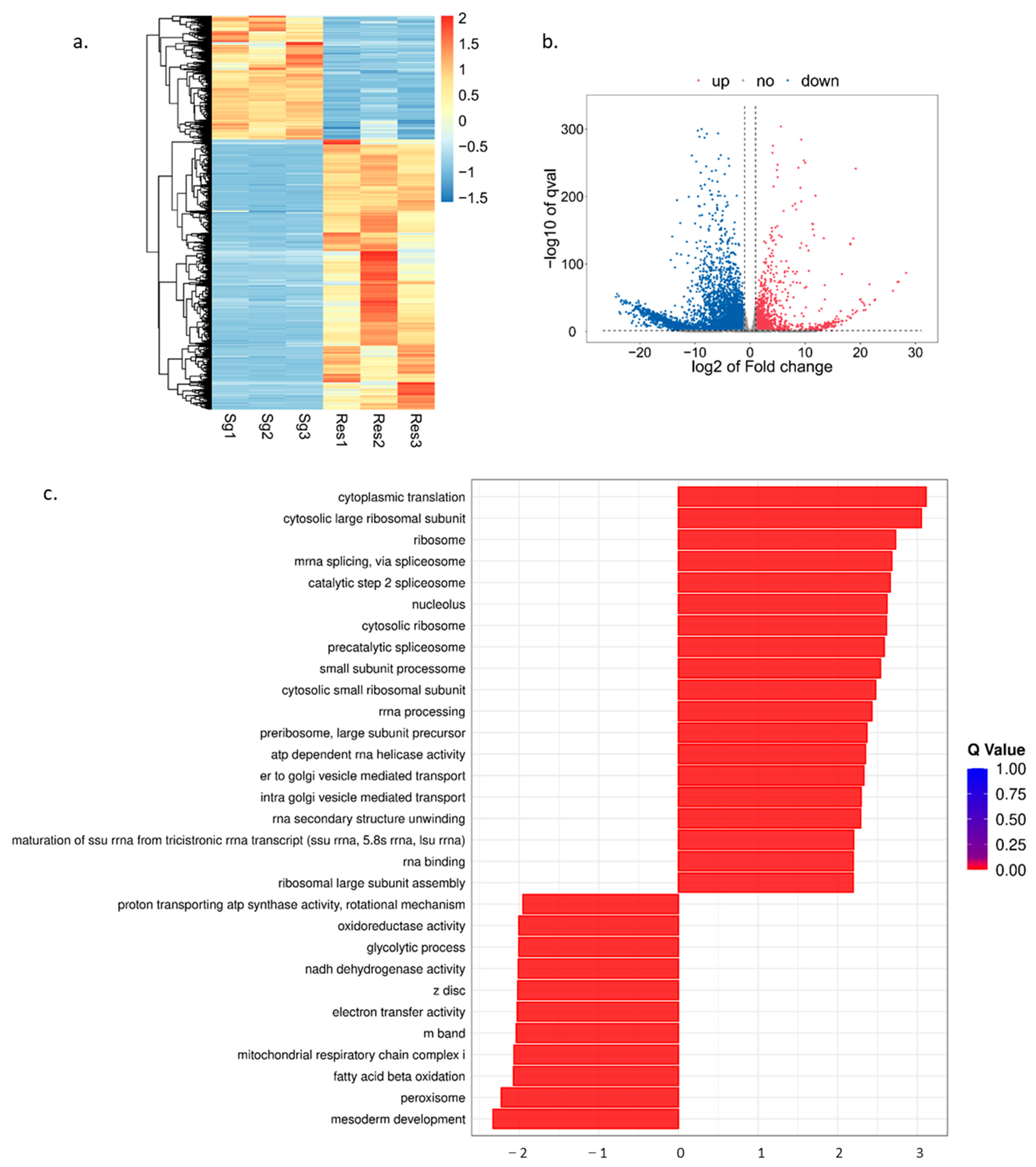
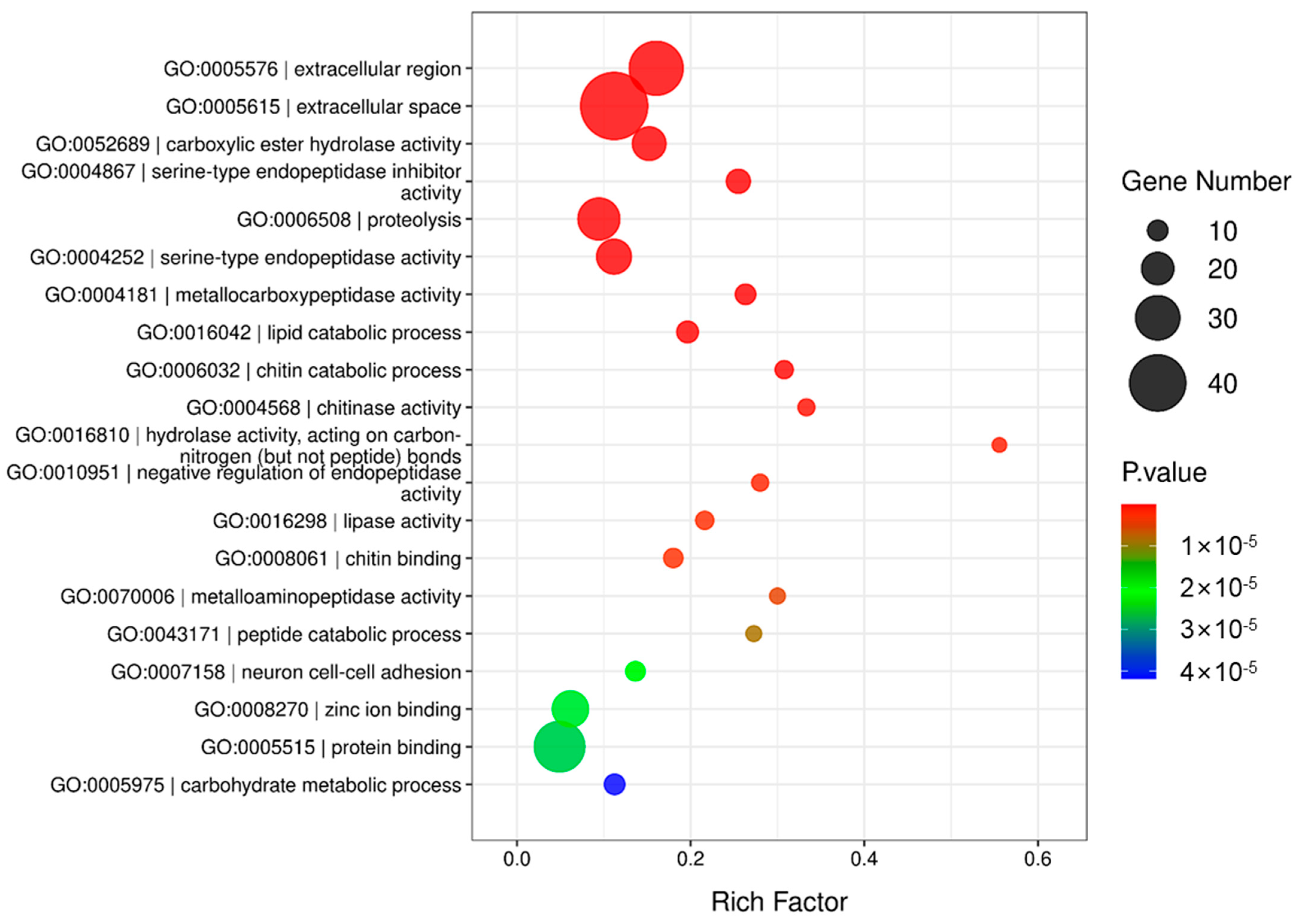
3.1. Transcriptome Analysis of O. furnacalis
3.2. Proteomic Analysis of OS in O. furnacalis
3.3. Combined Analysis of OS Proteins
3.4. Putative Effector Proteins Prediction
3.5. Tissue-Specific Expressions of Putative Effector Protein Gene
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Q.; Shi, J.; Huang, C.; Guo, J.; He, K.; Wang, Z. Asian Corn Borer (Ostrinia furnacalis) Infestation Increases Fusarium verticillioides Infection and Fumonisin Contamination in Maize and Reduces the Yield. Plant Dis. 2023, 107, 1557–1564. [Google Scholar] [CrossRef]
- Dai, X.; Chen, H.; Liu, Y.; Wang, R.; Su, L.; Wang, Y.; Zhao, S.; Wang, C.; Zheng, L.; Zhai, Y. Trichogramma ostriniae as a sustainable biological control agent against Ostrinia furnacalis: An eight-year field and laboratory study in China. J. Pest. Sci. 2026, 99, 40. [Google Scholar] [CrossRef]
- Liu, K.; Zhang, T.; Zhang, Y.; Wang, Z.; He, K. Chromosome-level genome of a multivoltine biotype Ostrinia furnacalis strain. Sci. Data. 2025, 12, 881. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhou, W.; Huang, B.; Gao, M.; Li, Q.; Tao, Y.; Wang, Z. Molecular and Functional Characterization of Peptidoglycan Recognition Proteins OfPGRP-A and OfPGRP-B in Ostrinia furnacalis (Lepidoptera: Crambidae). Insects 2022, 13, 417. [Google Scholar] [CrossRef]
- Abbas, A.; Saddam, B.; Ullah, F.; Hassan, M.A.; Shoukat, K.; Hafeez, F.; Alam, A.; Abbas, S.; Ghramh, H.A.; Khan, K.A.; et al. Global distribution and sustainable management of Asian corn borer (ACB), Ostrinia furnacalis (Lepidoptera: Crambidae): Recent advancement and future prospects. Bull. Entomol. Res. 2025, 115, 105–120. [Google Scholar] [CrossRef]
- Trisyono, Y.A.; Aryuwandari, V.E.F.; Sinulingga, N.G.H.; Andika, I.P. Damage assessment of the Asian corn borer, Ostrinia furnacalis (Lepidoptera: Crambidae), in corn production centers in Indonesia. Heliyon 2025, 11, e42795. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Luan, B.; Wang, H.; Wang, P.; Gao, X.; Liu, B. Analysis of Population Dynamics and Migration Status of Ostrinia furnacalis Under Searchlight Trap and Ground Light-Trap. Sci. Agric. Sin. 2024, 57, 4071–4081. [Google Scholar] [CrossRef]
- Li, Q.; Shi, J.; Huang, C.; Guo, J.; Hellmich, R.L.; He, K.; Wang, Z. Roles of Ear Injury, Infestation, and Vector Activity by Ostrinia furnacalis (Asian Corn Borer) and Conogethes punctiferalis (Yellow Peach Moth) in Fusarium verticillioides Infection and Kernel Fumonisin Level. Phytopathology 2023, 113, 1867–1875. [Google Scholar] [CrossRef]
- Guo, J.; Qi, J.; He, K.; Wu, J.; Bai, S.; Zhang, T.; Zhao, J.; Wang, Z. The Asian corn borer Ostrinia furnacalis feeding increases the direct and indirect defence of mid-whorl stage commercial maize in the field. Plant Biotechnol. J. 2019, 17, 88–102. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef]
- Futuyma, D.J.; Agrawal, A.A. Macroevolution and the biological diversity of plants and herbivores. Proc. Natl. Acad. Sci. USA 2009, 106, 18054–18061. [Google Scholar] [CrossRef]
- Villarroel, C.A.; Jonckheere, W.; Alba, J.M.; Glas, J.J.; Dermauw, W.; Haring, M.A.; Van Leeuwen, T.; Schuurink, R.C.; Kant, M.R. Salivary proteins of spider mites suppress defenses in Nicotiana benthamiana and promote mite reproduction. Plant J. 2016, 86, 119–131. [Google Scholar] [CrossRef]
- Erb, M.; Reymond, P. Molecular Interactions Between Plants and Insect Herbivores. Annu. Rev. Plant Biol. 2019, 70, 527–557. [Google Scholar] [CrossRef]
- Huang, H.; Li, J.; Chen, J.; Zhang, C. Composition, evolution, and functions of herbivorous insect saliva. Sci. Sin. Vitae 2025, 55, 1646–1660. [Google Scholar] [CrossRef]
- Hogenhout, S.A.; Bos, J.I. Effector proteins that modulate plant–insect interactions. Curr. Opin. Plant Biol. 2011, 14, 422–428. [Google Scholar] [CrossRef]
- Acevedo, F.E.; Peiffer, M.; Tan, C.W.; Stanley, B.A.; Stanley, A.; Wang, J.; Jones, A.G.; Hoover, K.; Rosa, C.; Luthe, D.; et al. Fall Armyworm-Associated Gut Bacteria Modulate Plant Defense Responses. Mol. Plant Microbe Interact. 2017, 30, 127–137. [Google Scholar] [CrossRef]
- Basu, S.; Varsani, S.; Louis, J. Altering Plant Defenses: Herbivore-Associated Molecular Patterns and Effector Arsenal of Chewing Herbivores. Mol. Plant Microbe Interact. 2018, 31, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, F.E.; Stanley, B.A.; Stanley, A.; Peiffer, M.; Luthe, D.S.; Felton, G.W. Quantitative proteomic analysis of the fall armyworm saliva. Insect Biochem. Mol. Biol. 2017, 86, 81–92. [Google Scholar] [CrossRef]
- Zhu, Y.; Yao, J.; Luttrell, R. Identification of Genes Potentially Responsible for extra-Oral Digestion and Overcoming Plant Defense from Salivary Glands of the Tarnished Plant Bug (Hemiptera: Miridae) Using cDNA Sequencing. J. Insect Sci. 2016, 16, 60. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Vega, L.J.; Acevedo, F.E.; Felton, G.W. Genomics of Lepidoptera saliva reveals function in herbivory. Curr. Opin. Insect Sci. 2017, 19, 61–69. [Google Scholar] [CrossRef]
- Eberhard, S.H.; Hrassnigg, N.; Crailsheim, K.; Krenn, H.W. Evidence of protease in the saliva of the butterfly Heliconius melpomene (L.) (Nymphalidae, Lepidoptera). J. Insect Physiol. 2007, 53, 126–131. [Google Scholar] [CrossRef]
- Dong, Y.; Jing, M.; Shen, D.; Wang, C.; Zhang, M.; Liang, D.; Nyawira, K.T.; Xia, Q.; Zuo, K.; Wu, S.; et al. The mirid bug Apolygus lucorum deploys a glutathione peroxidase as a candidate effector to enhance plant susceptibility. J. Exp. Bot. 2020, 71, 2701–2712. [Google Scholar] [CrossRef]
- Zhang, H.; Chi, Y.; Chen, S.; Lv, X.; Jia, D.; Chen, Q.; Wei, T. Scavenging H2O2 of plant host by saliva catalase of leafhopper vector benefits viral transmission. New Phytol. 2024, 243, 2368–2384. [Google Scholar] [CrossRef]
- Eichenseer, H.; Mathews, M.C.; Bi, J.L.; Murphy, J.B.; Felton, G.W. Salivary glucose oxidase: Multifunctional roles for helicoverpa zea? Arch. Insect Biochem. Physiol. 1999, 42, 99–109. [Google Scholar]
- Wang, J.; Yang, M.; Song, Y.; Acevedo, F.E.; Hoover, K.; Zeng, R.; Felton, G.W. Gut-Associated Bacteria of Helicoverpa zea Indirectly Trigger Plant Defenses in Maize. J. Chem. Ecol. 2018, 44, 690–699, Correction in J. Chem. Ecol. 2018, 44, 700. [Google Scholar] [CrossRef]
- Hoballah, M.E.; Turlings, T.C. The role of fresh versus old leaf damage in the attraction of parasitic wasps to herbivore-induced maize volatiles. J. Chem. Ecol. 2005, 31, 2003–2018. [Google Scholar] [CrossRef]
- Prajapati, V.K.; Vijayan, V.; Vadassery, J. Secret Weapon of Insects: The Oral Secretion Cocktail and Its Modulation of Host Immunity. Plant Cell Physiol. 2024, 65, 1213–1223. [Google Scholar] [CrossRef]
- Jones, A.C.; Felton, G.W.; Tumlinson, J.H. The dual function of elicitors and effectors from insects: Reviewing the ‘arms race’ against plant defenses. Plant Mol. Biol. 2022, 109, 427–445. [Google Scholar] [CrossRef]
- Chen, C.; Liu, Y.; Song, W.; Chen, D.; Chen, F.; Chen, X.; Chen, Z.; Ge, S.; Wang, C.; Zhan, S. An effector from cotton bollworm oral secretion impairs host plant defense signaling. Proc. Natl. Acad. Sci. USA 2019, 116, 14331–14338. [Google Scholar] [CrossRef]
- Lyu, F.; Han, F.; Ge, C.; Mao, W.; Chen, L.; Hu, H.; Chen, G.; Lang, Q.; Fang, C. OmicStudio: A composable bioinformatics cloud platform with real-time feedback that can generate high-quality graphs for publication. Imeta 2023, 2, e85. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Rao, W.; Zheng, X.; Liu, B.; Guo, Q.; Guo, J.; Wu, Y.; Shangguan, X.; Wang, H.; Wu, D.; Wang, Z. Secretome Analysis and In Planta Expression of Salivary Proteins Identify Candidate Effectors from the Brown Planthopper Nilaparvata lugens. Mol. Plant Microbe Interact. 2019, 32, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Lortzing, T.; Firtzlaff, V.; Nguyen, D.; Rieu, I.; Stelzer, S.; Schad, M.; Kallarackal, J.; Steppuhn, A. Transcriptomic responses of Solanum dulcamara to natural and simulated herbivory. Mol. Ecol. Resour. 2017, 17, e196–e211. [Google Scholar] [CrossRef]
- Parmagnani, A.S.; Maffei, M.E. Calcium Signaling in Plant-Insect Interactions. Plants 2022, 11, 2689. [Google Scholar] [CrossRef]
- Aljbory, Z.; Chen, M. Indirect plant defense against insect herbivores: A review. Insect Sci. 2018, 25, 2–23. [Google Scholar] [CrossRef]
- Simon, J.C.; d’Alençon, E.; Guy, E.; Jacquin-Joly, E.; Jaquiéry, J.; Nouhaud, P.; Peccoud, J.; Sugio, A.; Streiff, R. Genomics of adaptation to host-plants in herbivorous insects. Brief. Funct. Genomics. 2015, 14, 413–423. [Google Scholar] [CrossRef]
- Miao, Y.; Jia, H.; Li, Z.; Liu, Y.; Hou, M. Transcriptomic and Expression Analysis of the Salivary Glands in Brown Planthoppers, Nilaparvata lugens (Hemiptera: Delphacidae). J. Econ. Entomol. 2018, 111, 2884–2893. [Google Scholar] [CrossRef]
- Sharma, A.; Khan, A.N.; Subrahmanyam, S.; Raman, A.; Taylor, G.S.; Fletcher, M.J. Salivary proteins of plant-feeding hemipteroids—Implication in phytophagy. Bull. Entomol. Res. 2014, 104, 117–136. [Google Scholar] [CrossRef]
- Dong, Y.; Zhang, W.; Jin, Y.; Shen, D.; Xia, A. Apolygus lucorum effector Al6 promotes insect feeding performance on soybean plants: RNAi analysis and feeding behaviour study with electrical penetration graph. Insect Mol. Biol. 2023, 32, 12808. [Google Scholar] [CrossRef]
- Shang, Z.; Yang, J.; Zhang, R.; Liu, D. Functional analyses of the salivary protein SaE23 in Sitobion avenae. Int. J. Biol. Macromol. 2025, 307, 142068. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Jia, H.; Cui, X.; Zhai, N.; Wang, H.; Guo, X.; Xu, B. Isolation of carboxylesterase (esterase FE4) from Apis cerana cerana and its role in oxidative resistance during adverse environmental stress. Biochimie 2018, 144, 85–97. [Google Scholar] [CrossRef]
- Cai, X.; Wang, Y.; Li, X.; Zhang, Z.; Ul Haq, I.; Wang, J.; Hou, Y. Detecting the Role of Salivary Protein Genes in Spodoptera frugiperda Adaptation to Different Host Plants. J. Agric. Food Chem. 2025, 73, 22295–22310. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Ji, R.; Fu, J.; Li, J.; Wang, L.; Zhang, H.; Yang, S.; Ye, W.; Fang, J.; Zhu-Salzman, K. A salivary calcium-binding protein from Laodelphax striatellus acts as an effector that suppresses defense in rice. Pest. Manag. Sci. 2021, 77, 2272–2281. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Yi, G.; Lv, X.; Mao, Q.; Wei, T. A leafhopper saliva protein mediates horizontal transmission of viral pathogens from insect vectors into rice phloem. Commun. Biol. 2022, 5, 204. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Peng, Z.; Tong, H.; Xie, W.; Wang, S.; Wu, Q.; Zhang, J.; Li, C.; Zhang, Y. A salivary ferritin in the whitefly suppresses plant defenses and facilitates host exploitation. J. Exp. Bot. 2019, 70, 3343–3355. [Google Scholar] [CrossRef]
- Duchemin, J.B.; Paradkar, P.N. Iron availability affects West Nile virus infection in its mosquito vector. Virol. J. 2017, 14, 103. [Google Scholar] [CrossRef]
- Zheng, X.; Wan, Y.; Tao, M.; Yuan, J.; Zhang, K.; Wang, J.; Zhang, Y.; Liang, P.; Wu, Q. Obstructor, a Frankliniella occidentalis protein, promotes transmission of tomato spotted wilt orthotospovirus. Insect Sci. 2023, 30, 741–757. [Google Scholar] [CrossRef] [PubMed]
- Kallure, G.S.; Kumari, A.; Shinde, B.A.; Giri, A.P. Characterized constituents of insect herbivore oral secretions and their influence on the regulation of plant defenses. Phytochemistry 2022, 193, 113008. [Google Scholar] [CrossRef]
- Terra, W.R.; Ferreira, C. Evolutionary trends of digestion and absorption in the major insect orders. Arthropod Struct. Dev. 2020, 56, 100931. [Google Scholar] [CrossRef]
- Chen, G.; Gao, X.; Zhang, Y.; Ma, C.; Ma, W.; Zhou, Z. The carboxypeptidase B and carbonic anhydrase genes play a reproductive regulatory role during multiple matings in Ophraella communa. Front. Mol. Biosci. 2023, 10, 1095645. [Google Scholar] [CrossRef]
- Cao, H.; Zhang, S.; Zhu, L.; Wang, J.; Liu, Y.; Wang, Y.; Kong, X.; You, L.; Toufeeq, S.; Liu, S. The digestive proteinase trypsin, alkaline A contributes to anti-BmNPV activity in silkworm (Bombyx mori). Dev. Comp. Immunol. 2021, 119, 104035. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.A.; Kim, W.J.; Lee, S.; Yang, H.S.; Lee, B.H.; Lee, S.H. Comparative analyses of the venom components in the salivary gland transcriptomes and saliva proteomes of some heteropteran insects. Insect Sci. 2022, 29, 411–429. [Google Scholar] [CrossRef] [PubMed]
- Winssy, T.D.; Gopal, N.O.; Indiragandhi, P.; Balasubramani, V.; Anandham, R. Chitinase Producing Gut-Associated Bacteria Affected the Survivability of the Insect Spodoptera frugiperda. Front. Biosci. 2024, 16, 15. [Google Scholar] [CrossRef]
- Liu, T.; Guo, X.; Bu, Y.; Zhou, Y.; Duan, Y.; Yang, Q. Structural and biochemical insights into an insect gut-specific chitinase with antifungal activity. Insect Biochem. Mol. Biol. 2020, 119, 103326. [Google Scholar] [CrossRef] [PubMed]
- Da Lage, J.L. The Amylases of Insects. Int. J. Insect Sci. 2018, 10, 1179543318804783. [Google Scholar] [CrossRef]
- Zhang, L.; Cai, Q.; Cui, J.; Zhang, W.; Dong, S.; Xiao, W.; Li, J.; Kou, T.; Zhang, X.; He, H.; et al. A secreted-Cu/Zn superoxide dismutase from Microplitis bicoloratus reduces reactive oxygen species triggered by symbiotic bracovirus. Dev. Comp. Immunol. 2019, 92, 129–139. [Google Scholar] [CrossRef]
- Zdybicka-Barabas, A.; Stączek, S.; Kunat-Budzyńska, M.; Cytryńska, M. Innate Immunity in Insects: The Lights and Shadows of Phenoloxidase System Activation. Int. J. Mol. Sci. 2025, 26, 1320. [Google Scholar] [CrossRef]
- Shakeel, M.; Xu, X.; De Mandal, S.; Jin, F. Role of serine protease inhibitors in insect-host-pathogen interactions. Arch. Insect Biochem. Physiol. 2019, 102, e21556. [Google Scholar] [CrossRef]
- Wang, Q.; Ren, M.; Liu, X.; Xia, H.; Chen, K. Peptidoglycan recognition proteins in insect immunity. Mol. Immunol. 2019, 106, 69–76. [Google Scholar] [CrossRef]
- Prajapati, V.K.; Varma, M.; Vadassery, J. In silico identification of effector proteins from generalist herbivore Spodoptera litura. BMC Genom. 2020, 21, 819. [Google Scholar] [CrossRef]
- Acevedo, F.E.; Rivera-Vega, L.J.; Chung, S.H.; Ray, S.; Felton, G.W. Cues from chewing insects—The intersection of DAMPs, HAMPs, MAMPs and effectors. Curr. Opin. Plant Biol. 2015, 26, 80–86. [Google Scholar] [CrossRef]
- Feng, X.; Liu, N. Functional Characterization of Carboxylesterases in Insecticide Resistant House Flies, Musca Domestica. J. Vis. Exp. 2018, 138, 58106. [Google Scholar] [CrossRef]
- Ye, W.; Yu, H.; Jian, Y.; Zeng, J.; Ji, R.; Chen, H.; Lou, Y. A salivary EF-hand calcium-binding protein of the brown planthopper Nilaparvata lugens functions as an effector for defense responses in rice. Sci. Rep. 2017, 7, 40498. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Shi, Y.; Wang, L.; Tian, T.; Li, J.; Gong, L.; Zheng, Z.; Jing, M.; Fang, J.; Ji, R. Planthopper-Secreted Salivary Calmodulin Acts as an Effector for Defense Responses in Rice. Front. Plant Sci. 2022, 13, 841378. [Google Scholar] [CrossRef] [PubMed]
- Bendtsen, J.D.; Jensen, L.J.; Blom, N.; Von Heijne, G.; Brunak, S. Feature-based prediction of non-classical and leaderless protein secretion. PEDS 2004, 17, 349–356. [Google Scholar] [CrossRef]
- Rabouille, C. Pathways of Unconventional Protein Secretion. Trends Cell Biol. 2017, 27, 230–240. [Google Scholar] [CrossRef]
- Gee, H.Y.; Kim, J.; Lee, M.G. Unconventional secretion of transmembrane proteins. Semin. Cell Dev. Biol. 2018, 83, 59–66. [Google Scholar] [CrossRef]
- Filaquier, A.; Marin, P.; Parmentier, M.L.; Villeneuve, J. Roads and hubs of unconventional protein secretion. Curr. Opin. Cell Biol. 2022, 75, 102072. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Category of Genes | Putative Effector Proteins Proteome | No. of Sequences in Proteome | Gene | References |
|---|---|---|---|---|
| enzymes | serine protease | 2 | LOC114361239 | [27,36,37,38] |
| LOC114366020 | ||||
| metalloprotease | 1 | LOC114359768 | ||
| glucose dehydrogenase | 3 | LOC114355043 | ||
| LOC114355055 | ||||
| LOC114356009 | ||||
| aldehyde dehydrogenase | 1 | LOC114357983 | ||
| detoxifying proteins | peroxidase like | 3 | LOC114363884 | [22,39,40] |
| LOC114363885 | ||||
| LOC114358750 | ||||
| esterase FE4-like | 2 | LOC114361121 | [41,42] | |
| LOC114364938 | ||||
| calcium-binding proteins | regucalcin-like | 1 | LOC114356474 | [43,44] |
| calcium-binding protein | 1 | LOC114361112 | ||
| other proteins | ferritin | 1 | LOC114358414 | [45,46] |
| obstructor-E-like | 1 | LOC114359168 | [47] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Sun, X.; Hu, W.; Wang, D.; Zhu, M.; Xin, C.; Yang, W.; Li, H.; Cao, Y. Identification of Oral Secretion Proteins in Ostrinia furnacalis by Transcriptome and LC-MS/MS Analyses. Insects 2026, 17, 357. https://doi.org/10.3390/insects17040357
Sun X, Hu W, Wang D, Zhu M, Xin C, Yang W, Li H, Cao Y. Identification of Oral Secretion Proteins in Ostrinia furnacalis by Transcriptome and LC-MS/MS Analyses. Insects. 2026; 17(4):357. https://doi.org/10.3390/insects17040357
Chicago/Turabian StyleSun, Xinyan, Wei Hu, Dan Wang, Meichen Zhu, Cuiping Xin, Wenbo Yang, Huimin Li, and Yanyong Cao. 2026. "Identification of Oral Secretion Proteins in Ostrinia furnacalis by Transcriptome and LC-MS/MS Analyses" Insects 17, no. 4: 357. https://doi.org/10.3390/insects17040357
APA StyleSun, X., Hu, W., Wang, D., Zhu, M., Xin, C., Yang, W., Li, H., & Cao, Y. (2026). Identification of Oral Secretion Proteins in Ostrinia furnacalis by Transcriptome and LC-MS/MS Analyses. Insects, 17(4), 357. https://doi.org/10.3390/insects17040357