Simple Summary
Tetranychus evansi Baker et Pritchard and Tetranychus urticae Koch (Acari: Tetranychidae) are significant agricultural pests. Phytoseiulus persimilis (Acari: Phytoseiidae) is one of the most effective natural enemies for controlling Tetranychus pests. This study compared the influence of host plants on the biological performance of P. persimilis when preying on these two Tetranychus species raised on either bean or potato plants. A key finding of this study was that T. evansi does not serve as suitable prey for P. persimilis. Furthermore, potato plants were shown to indirectly reduce the effectiveness of P. persimilis by adversely affecting the pest population itself. This research contributes to a better understanding of the complex interactions among crops, pests, and their natural enemies—insights that are important for developing more effective pest management strategies.
Abstract
In this study, P. persimilis was provided with T. evansi and T. urticae that had been reared on either bean or potato plants to investigate the effects of both prey and host plant species on the predator’s growth, development, and fitness. The results indicate that the reproductive potential of P. persimilis populations fed T. evansi was significantly lower than that of populations fed T. urticae from the same host plant (p < 0.01). Phytoseiulus persimilis fed T. evansi that had been reared on potatoes showed poorer performance in oviposition period, post-oviposition period, daily egg production, and total egg production compared to those fed T. evansi reared on beans (p < 0.01). The intrinsic rate of increase (rm) of P. persimilis fed on T. evansi reared on potato was 0.08, which was 55.56% lower than that of populations fed on T. evansi reared on beans. This study sheds light on the complex interactions among host plants, pests, and their natural enemies, thereby providing a theoretical basis for developing more effective and sustainable management strategies against T. evansi that take these intricate ecological relationships into account.
1. Introduction
The spider mites Tetranychus urticae Koch and Tetranychus evansi Baker et Pritchard (Acari: Tetranychidae) are both important pest species within the genus Tetranychus Dufor. Tetranychus evansi is a highly destructive pest of numerous economically important crops, particularly those in the Family Solanaceae [1,2]. First reported in 1960 from specimens collected on tomato in Mauritius [3], this pest has rapidly spread and demonstrated strong adaptability, becoming a significant invasive agricultural pest worldwide [4,5]. Tetranychus evansi poses a considerable threat to the production of solanaceous crops, as well as to a range of other vegetables and ornamental plants in affected regions. Tetranychus urticae is a major pest in both open field and protected cultivation systems worldwide [6,7]. It is notorious for its polyphagous nature, having been documented on hosts from more than 140 botanical families [8].
Research interest in the natural enemies of T. evansi emerged even before the species was formally described [9]. Early records in the late 1970s noted the co-occurrence of a phytoseiid mite, as well as several predatory insects from the families Miridae, Coccinellidae, and Thripidae, with T. evansi populations [10]. This observation prompted wider international efforts to identify effective biological control agents against this pest. In 1982, due to the significant damage caused by T. evansi to potatoes and other solanaceous crops in Southern California, the University of California, Riverside launched a research program to evaluate potential natural enemies [11]. In laboratory assays, eight phytoseiid mite species—including Galendromus annectens (De Leon), Galendromus occidentalis (Nesbitt), Galendromus porresi (McMurtry), Neoseiulus californicus (McGregor), Phytoseiulus longipes Evans, Phytoseiulus macropilis (Banks), Phytoseiulus persimilis Athias-Henriot, and Phytoseius hawaiiensis Prasad—were tested for their predatory efficacy against T. evansi. The results showed very low oviposition and survival rates among these predators when feeding on T. evansi, indicating their limited suitability as effective biocontrol agents [12].
Phytoseiid mites are among the most widely used commercial biological control agents globally, particularly for managing small arthropod pests such as spider mites [13,14]. Among them, P. persimilis is a specialist predator known for its efficacy primarily against T. urticae [15,16]. Interestingly, despite the generally poor performance of phytoseiids against T. evansi in earlier screenings, de Moraes (1986) found that P. persimilis could initially locate and feed on T. evansi as readily as on T. urticae [17]. A feeding deterrent appeared to prolong the handling time for T. evansi eggs, suggesting possible physiological or chemical defenses in this prey species. Despite these findings, the sublethal effects of T. evansi on P. persimilis—particularly regarding reproduction, development, and population growth—remain poorly understood.
Predator efficacy is shaped by both abiotic (e.g., temperature and humidity) and biotic factors such as prey species, host plant identity, and predator sex [18,19,20,21]. Prey type directly influences predator development, survival, and fecundity [22,23,24,25,26], while host plant traits can mediate tri-trophic interactions, altering pest distribution and natural enemy performance [27,28,29,30,31]. Given these dynamics, this study uses T. urticae—a suitable prey for P. persimilis—as a control to systematically evaluate the impact of T. evansi on the development and reproduction of P. persimilis. Additionally, we examine how T. evansi reared on different host plants affect the predator’s performance. Our findings help clarify why P. persimilis is an inadequate biocontrol agent for T. evansi and contribute to the development of integrated management strategies that account for these ecological complexities.
2. Materials and Methods
2.1. Plants Material and Mite Colonies
Bean (‘Hong Hua’, Shenyang Best Agricultural Technology Co., Ltd., Shenyang, China) and potato (‘Ma ErKe’, Mianyang Green Control Technology Co., Ltd., Mianyang, China) were selected as host plants for rearing T. urticae and T. evansi. Bean seeds and potato tubers were planted separately in plastic pots measuring 12 cm in diameter. The seedlings were grown using conventional methods until they reached the two-true leaf stage.
The populations of T. urticae and T. evansi originated from individuals collected in farmland around Mianyang City, Sichuan Province, in 2019. These populations were reared on potted bean seedlings maintained by the College of Life Sciences and Technology, Mianyang Normal University. Prior to this experiment, equal numbers of adult T. urticae and T. evansi were inoculated onto the two host plant species, and then reared separately for five generations in two chambers located 200 m apart. Fresh host plants were replaced at regular intervals to maintain a stable population of the mites.
Additionally, the colony of P. persimilis was supplied by Shoubonong Biotech in 2025. Phytoseiulus persimilis populations was maintained in circular plastic containers, each containing a sponge with a height of 5 cm. A layer of filter paper and a sheet of black plastic film, both cut to decreasing diameters, were placed on top of the sponge. Leaves infested with T. urticae were then placed on the black plastic film.
2.2. Experiment Set-Up
For individual rearing, P. persimilis was maintained in custom acrylic chambers. Each chamber was constructed by sealing a layer of host plant leaf, a piece of filter paper, and a glass slide beneath a transparent acrylic board (30 × 20 × 3 mm) with a central opening 10 mm in diameter. The entire assembly was secured at both ends with clips to prevent mites from escaping (Figure 1) [22].
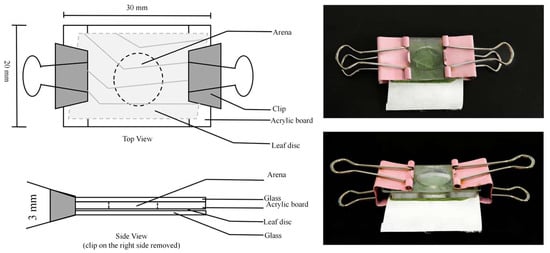
Figure 1.
Schematic views and photographs of the experimental unit.
To test the combined effects of prey species and host plant, four treatments were established in which P. persimilis were fed: (i) T. urticae reared on bean, (ii) T. urticae reared on potato, (iii) T. evansi reared on bean, and (iv) T. evansi reared on potato (see Figure 2 for experimental design). In each treatment, 50 gravid female P. persimilis were assigned (five individuals per container, for a total of 10 chambers). For each treatment, eggs were collected from each treatment after a 12 h oviposition period. From the 120 eggs obtained per treatment, three biological replicates of 40 eggs each were prepared. Subsequently, from each of these replicate chambers, the adult female and all but one egg were removed, leaving a single egg per chamber. The developmental stages of each individual P. persimilis were recorded twice daily (at 8:00 and 20:00) until they reached adulthood. The duration of each developmental stage was then calculated. During the initial phase (prior to the larval stage), the same leaf disc was kept for each individual. Thereafter, the leaf disc was renewed daily until the experiment concluded. Upon reaching adulthood, each female P. persimilis was paired with a male from the same cohort in the rearing chambers. After 24 h, the male was removed, and the female was reared individually until death. Daily, ten adult females each of T. urticae and T. evansi were provided as prey to P. persimilis [29]. The daily egg production of each female was recorded at 24 h intervals until it died, Parameters including the pre-oviposition period, oviposition period, post-oviposition period, longevity, daily fecundity, and cumulative fecundity were then calculated. For each treatment, a minimum of 30 individuals that completed the entire life cycle was required. If any individual escaped or died due to non-experimental causes, the experiment was repeated from the egg stage.
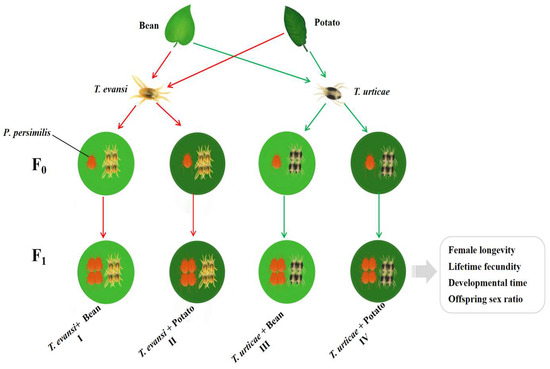
Figure 2.
The experimental design (Red arrows indicate T. evansi; Green arrows indicate T. urticae).
All experiments were conducted under controlled conditions, maintained at: 25 ± 1 °C, 60% relative humidity, and a 16:8 h (light–dark) photoperiod.
2.3. Life History Parameters and Statistical Analysis
The life table parameters of the experimental populations were calculated according to the following formulas:
where x represents the age interval in days; lx denotes the age-specific survival rate of female P. persimilis; mx indicates the age-specific fecundity (average number of female offspring produced per surviving female at age x); R0 is the net reproductive rate (average number of female offspring produced per female over its lifetime); T is the mean generation time (the average time interval between the birth of a parent and the birth of its offspring); λ is the finite rate of increase (population multiplication factor unit time); t is the population doubling time and e is the base of natural logarithms [32].
R0 = ∑lx mx;
T = ∑x lx mx/R0
rm = ln (R0)/T
λ = erm
t = ln (2)/rm
Two-way ANOVA was used to analyze the effects of plant and prey on the life table parameters of P. persimilis. Separately, the Mann–Whitney U test was used to compare the effects of host plant and prey species on specific life history traits: developmental time, longevity, fecundity and oviposition time. All Statistical analyses were performed using SPSS (version 22.0) software. Figures were generated using GraphPad Prism (version 8.0.2).
3. Results
3.1. Influence of Host Plants on Immature Stages of P. persimilis Fed with T. urticae and T. evansi
Phytoseiulus persimilis developed into adults after feeding on T. urticae reared on beans, T. urticae reared on potatoes, T. evansi reared on beans, and T. evansi reared on potatoes, with 110, 112, 107, and 98 individuals, starting from 120 eggs per treatment (Figure 3). When P. persimilis fed on T. urticae reared on potatoes, its egg stage was significantly longer than when it fed on T. urticae reared on beans (Z = −7.571, p < 0.001). The deutonymph development period after feeding on T. urticae reared on beans was significantly longer than after feeding on T. urticae reared on potatoes (Z = −9.077, p < 0.001). When P. persimilis fed on T. evansi reared on beans, its egg stage was significantly longer than that when fed on T. evansi reared on potatoes (Z = −4.872, p < 0.001). In contrast, the deutonymphal development period of P. persimilis was significantly longer after feeding on T. evansi reared on potatoes than reared on beans (Z = −4.317, p < 0.001). No significant differences were detected in the larval stage, protonymphal stage durations, or total development time of P. persimilis across the four prey-host plant combinations (p > 0.05). When preying on T. urticae (reared on bean or potato plants), P. persimilis had significantly longer egg, nymphal, and adult stages than when preying on T. evansi (p < 0.001). However, the protonymphal stage of P. persimilis was significantly longer when feeding on T. evansi compared to T. urticae, regardless of the host plant (p < 0.001) (Figure 3).
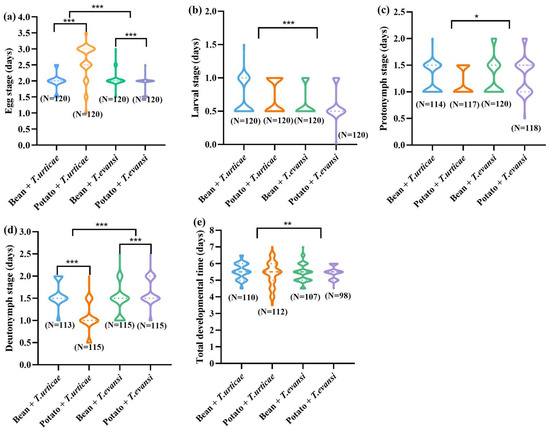
Figure 3.
The effect of the host plants on the duration of the immature stages of P. persimilis fed with T. urticae and T. evansi ((a) is egg; (b) is larva; (c) is protonymph; (d) is deutonymph; (e) is egg to adult). The asterisk stands for significant difference. * p < 0.05; ** p < 0.01; *** p < 0.001.
In summary, neither the plant nor the pest mite species had a significant effect on the immature development of P. persimilis (p > 0.05) (Table 1). In contrast, the plant × mite species interaction was significant for the egg, larval, and nymphal stages (p < 0.001) (Table 1), but not for the total developmental time (F(1, 387) = 3.478, p = 0.063).

Table 1.
Effects of Plants and species on the developmental duration, fecundity, and longevity of P. persimilis.
3.2. Influence of Host Plants on Life Span of P. persimilis Fed with T. urticae and T. evansi
After feeding on T. evansi reared on potatoes, the pre-oviposition period of P. persimilis was extended by 79.17%, 62.30%, and 55.48% compared to feeding on T. urticae reared on beans, T. urticae reared on potatoes, and T. evansi reared on beans, respectively (Figure 4). The pre-oviposition period of P. persimilis was significantly longer when feeding on T. urticae reared on potatoes than on beans (Z = −7.492, p < 0.001). In contrast, its oviposition period was significantly longer on beans compared to potatoes (Z = −7.652, p < 0.001). No significant difference was observed in the post-oviposition period between the two host plant treatments (Z = −1.157, p = 0.247). The pre-oviposition period of P. persimilis was significantly longer when feeding on potato-reared T. urticae than on bean-reared T. urticae (Z = −4.995, p < 0.001). In contrast, both the oviposition period (Z = −2.919, p = 0.004) and post-ovipositions period (Z = −4.024, p < 0.001) were shorter on bean-reared prey. Feeding on T. urticae (from both beans and potatoes) resulted in a significantly shorter pre-oviposition period for P. persimilis compared to feeding on T. evansi from the same host plants (Z = −8.059, p < 0.001). In contrast, the oviposition period (Z = −9.860, p < 0.001) and post-oviposition period (Z = −3.094, p = 0.002) were significantly longer when P. persimilis preyed on T. urticae (Figure 4).
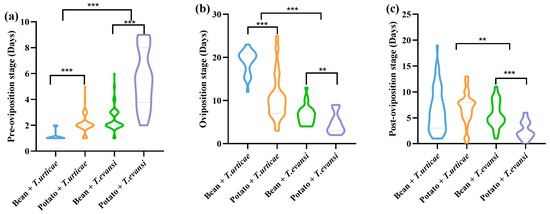
Figure 4.
The effect of the host plants on the life span of P. persimilis fed with T. urticae and T. evansi ((a) Pre-oviposition. (b) Oviposition. (c) Post-oviposition). The asterisk stands for significant difference. ** p < 0.01; *** p < 0.001.
For the adult stage of P. persimilis, both host plant species and prey mite species had significant effects (p < 0.05). Meanwhile, the host plant × prey mite species interaction was also significant for the pre-oviposition, oviposition, and post-oviposition periods (p < 0.05) (Table 1).
3.3. Influence of Host Plants on Fecundity of P. persimilis Fed with T. urticae and T. evansi
For both prey species, potato plants were associated with significantly lower fecundity of P. persimilis than with bean plants. When preying on T. urticae, daily and total fecundity were lower on potatoes (Z = −9.108, p < 0.001; Z = −9.059, p < 0.001). Similarly, when preying on T. evansi, both fecundity measures were also reduced on potatoes (Z = −4.792, p < 0.001; Z = −4.527, p < 0.001). Compared to T. urticae, consumption of T. evansi (from both bean and potato plants) led to significantly lower daily fecundity (Z = −7.923, p < 0.001) and total fecundity (Z = −9.266, p < 0.001) in P. persimilis (Figure 5).
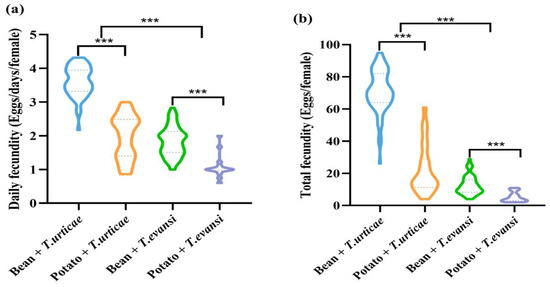
Figure 5.
The effect of the host plants on the fecundity of P. persimilis fed with T. urticae and T. evansi ((a) Daily fecundity. (b) Total fecundity). The asterisk stands for significant difference. *** p < 0.001.
Host plant species, prey pest mite species, and their interaction had significant effects on the daily and total fecundity of P. persimilis (p < 0.001) (Table 1).
3.4. Survival Rate and Daily Female Fecundity of P. persimilis Fed with T. urticae and T. evansi
As shown in Figure 6, the age-specific survival and fecundity of P. persimilis differed by prey and host plant. During development, mortality varied across treatments, with the highest immature mortality observed in the deutonymph stages for P. persimilis feeding on T. evansi reared on potatoes. In the adult stage, despite most individuals living over 20 days, the onset of mortality differed: it began on day 18 (T. urticae/beans), day 6 (T. urticae/potatoes), and day 7 for both T. evansi treatments, with complete mortality occurring by days 43, 29, 22, and 20, respectively.
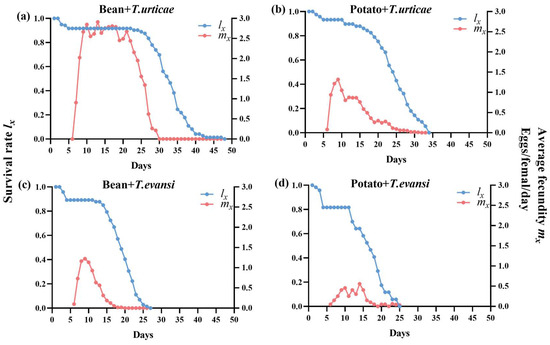
Figure 6.
Survival rate (lx) and age-specific females per day (mx) of P. persimilis fed on T. urticae and T. evansi reared on two different host plants ((a) is P. persimilis fed T. urticae reared on bean; (b) is P. persimilis fed T. urticae reared on potato; (c) is P. persimilis fed T. evansi reared on bean; (d) is P. persimilis fed T. evansi reared on potato).
In all four treatments, daily fecundity peaked shortly after reproduction began, with maximum age-specific rates reaching 2.91, 1.32, 1.22, and 0.56 eggs per female per day for P. persimilis feeding on T. urticae reared on beans, T. urticae reared on potatoes, T. evansi reared on beans, and T. evansi reared on potatoes, respectively.
3.5. Life Table Parameters of P. persimilis Fed on T. urticae and T. evansi Reared on Two Different Host Plants
The net reproductive rates (R0) of P. persimilis fed on T. urticae reared on beans, T. urticae reared on potatoes, T. evansi reared on beans, and T. evansi reared on potatoes were 44.16, 10.30, 6.32 and 2.53, respectively. The intrinsic rates of increase (rm) for P. persimilis across the four treatments were 0.23, 0.19, 0.18, 0.08, respectively. When P. persimilis was fed on T. evansi reared on potatoes, its intrinsic rate of increase (rm) was 65.22%, 57.89%, and 55.56% lower than that when preying on T. urticae reared on beans, T. urticae reared on potatoes, and T. evansi reared on beans, respectively (Table 2).
Table 2.
Life table parameters of P. persimilis fed on T. urticae and T. evansi reared on two different host plants.
4. Discussion
The results of this study indicate that the developmental period from egg to adult for P. persimilis when feeding on T. evansi reared on potatoes did not significantly differ from that time taken when feeding on T. urticae reared on potatoes or beans. However, during the adult stage, parameters including the oviposition period, post-oviposition period, daily egg production, and total egg production all exhibited reduced performance. de Moraes and McMurtry (1986) also found that P. persimilis detected and initiated feeding on T. evansi as effectively as on T. urticae [17]. However, the predation rate of P. persimilis was significantly lower on T. evansi compared to T. urticae. This result suggests that P. persimilis does not initially reject feeding on T. evansi, but over time, certain compounds in T. evansi may gradually impair the reproduction of P. persimilis, thereby reducing its effectiveness as a biological control agent against T. evansi.
Since P. persimilis cannot serve as an effective natural enemy of T. evansi, researchers have attempted to identify other viable natural enemies [33]. Sarmento (2004) observed that the amount of fat body in the generalist coccinellid predator Eriopis connexa (Germar) (Coleoptera) was significantly lower when it fed on T. evansi compared to when it was fed on an aphid species, even though the predator would choose to feed on T. evansi when given the option [34]. Another generalist coccinellid predator, Cycloneda sanguinea, was found by Oliveira (2005) to be unable to complete its developmental cycle when fed T. evansi [35]. These findings collectively indicate that the unsuitability of T. evansi as effective prey for natural enemies is not limited to a single predator species.
Several hypotheses have been proposed to explain the unsuitability of T. evansi as effective prey for natural enemies. Some researchers have suggested that the dense webbing produced by T. evansi may interfere with predation [36], while others have hypothesized that its association with solanaceous host plants could reduce its quality as prey for P. persimilis [37]. However, some studies have shown that the main factor leading to reduced fecundity of natural enemy populations is the prey species rather than the host plant. Similar studies have also demonstrated that the development of natural enemies is affected by the prey species they feed on, but not by the plants [38]. However, this study showed that the intrinsic rates of increase (rm) of P. persimilis fed with T. urticae reared on beans, T. urticae reared on potatoes, T. evansi reared on beans, and T. evansi reared on potatoes were 0.23, 0.19, 0.18, 0.08, respectively. This suggests that after multiple generations reared on bean-reared T. evansi, the reproductive performance of P. persimilis approached that of potato-reared T. urticae. These results confirm that the host plant of T. evansi is a major factor limiting its suitability as prey for P. persimilis, consistent with earlier scholarly speculation [39,40].
In tri-trophic interactions, the primary nutrients and the induced secondary metabolites of the host plants influence the physiology and behavior of pests, thereby affecting the growth, development, survival, and reproduction of their natural enemies [41,42]. In this study, T. evansi is generally recognized as an oligophagous pest of solanaceous crops. Tetranychus evansi has adapted to the secondary metabolites within plants during a long process of co-evolution, while these metabolites cannot be effectively utilized by its natural enemies. It has also been observed in other insects, for instance, Holzinger (1996) found that Danaus plexippus (L.) (Lepidoptera: Nymphalidae) can sequester toxic cardenolides from Digitalis purpurea L. (Lamiales: Plantaginaceae) through a process of chelation, thereby avoiding predation by its enemies [43]. Fordyce (2001) discovered that Battus philenor (L.) (Lepidoptera: Papilionidae) feeds exclusively on Aristolochia californica Torr. (Piperales: Aristolochiaceae; presently placed in the genus Isotrema Raf.), a plant containing alkaloids, and that the mortality rate and foraging efficiency of Chrysoperla Steinmann. (Neuroptera: Chrysopidae) larvae preying on B. philenor larvae were significantly lower than those of the control group [44]. Studies have shown that solanaceous crops contain abundant secondary metabolites such as flavonoids, terpenoids, solanine, methyl ketones, and 2-tridecanone [45]. Further investigation is required to determine which specific compounds accumulate in T. evansi, thereby inhibiting P. persimilis from acting as an effective natural enemy.
Finding effective natural enemies for T. evansi is one of the main strategies for controlling this mite. This study demonstrated that P. persimilis can effectively control T. evansi on beans. However, it remains uncertain whether extending the rearing time of the predatory mite on beans after five generations of T. evansi feeding will be more beneficial for the predator’s effectiveness. Athough P. persimilis effectively preys on T. evansi reared on beans, T. evansi itself poses little threat to bean in the field, which presents certain challenges for utilizing predatory mites to control this pest. Therefore, understanding the fundamental reasons behind how the accumulation of solanaceous crop metabolites affects predatory mites—such as their impact on enzyme activity, metabolites, and genes within the natural enemies—can provide insights for identifying effective natural enemies against T. evansi.
5. Conclusions
This study focused on the effects of prey species and host plants on the growth, development, and fitness of P. persimilis. By rearing T. evansi and T. urticae on beans and potatoes, respectively, and conducting predation experiments, the following core conclusions were drawn: under the same host plant condition, the reproductive potential of P. persimilis populations feeding on T. evansi was significantly lower than that of populations feeding on T. urticae (p < 0.01); Phytoseiulus persimilis feeding on T. evansi reared on potatoes showed the poorest performance in key reproductive indicators such as oviposition period, post-oviposition period, daily egg production, and total egg production (p < 0.01). Compared to those feeding on T. evansi reared on potatoes, P. persimilis feeding on T. evansi reared on beans exhibited a 55.56% higher intrinsic rate of increase (rm). In conclusion, T. evansi cannot serve as an effective prey for P. persimilis, and this result may be related to the solanaceous crop potato, which indirectly reduces the reproductive potential of P. persimilis by affecting T. evansi. This study revealed the complex ecological interaction mechanism among host plants, pests, and their natural enemies, thereby providing a theoretical basis for formulating more effective and sustainable management strategies for T. evansi that take into account these ecological relationships.
Author Contributions
Investigation, C.H., L.R., L.Y., L.X. and Q.T.; Data curation, S.B.; Writing—original draft, Y.Z.; Writing—review & editing, Y.Z.; Project administration, E.W.; Funding acquisition, E.W. All authors have read and agreed to the published version of the manuscript.
Funding
We thank the National Key R&D Program of China (2023YFD1401200) and Beijing Innovation Consortium of Agriculture Research System, BAIC01-2024 (2025) for supporting the publication of this article; Sichuan Science and Technology Program (No. 2024NSFSC1322); Mianyang Science and Technology Program (2023ZYDF076).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request (If needed, please contact Yannan Zhang at the following email address: zyn_082@163.com).
Acknowledgments
The author sincerely thanks Hao Xu of Mianyang Normal University for his assistance in correcting and revising the scientific names in Latin within the text. The author would also like to thank the three anonymous reviewers for their constructive criticism of this manuscript. The author has reviewed and edited the final version and assumes full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ferragut, F.; Luque, E.G.; Pekas, A. The Invasive Spider Mite Tetranychus evansi (Acari: Tetranychidae) Alters Community Composition and Host-Plant Use of Native Relatives. Exp. Appl. Acarol. 2013, 60, 321–341. [Google Scholar] [CrossRef] [PubMed]
- Migeon, A.; Nouguier, E.; Dorkeld, F. Spider Mites Web: A Comprehensive Database for the Tetranychidae; Springer: Dordrecht, The Netherlands, 2006; pp. 557–560. [Google Scholar]
- Baker, E.W.; Pritchard, A.E. The Tetranychoid Mites of Africa. Hilgardia 1960, 29, 455–574. [Google Scholar] [CrossRef]
- Boubou, A.; Migeon, A.; Roderick, G.K.; Navajas, M. Recent Emergence and Worldwide Spread of the Red Tomato Spider Mite, Tetranychus evansi: Genetic Variation and Multiple Cryptic Invasions. Biol. Invasions. 2011, 13, 81–92. [Google Scholar] [CrossRef]
- Tian, L.; Jin, P.Y.; Sun, C.P.; Hong, X.Y. First Distribution Record of the Tomato Red Spider Mite Tetranychus evansi (Acari: Tetranychidae) in Mainland China. Syst. Appl. Acarol. 2019, 24, 965–970. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Vontas, J.; Tsagkarakou, A.; Dermauw, W.; Tirry, L. Acaricide Resistance Mechanisms in the Two-spotted Spider Mite Tetranychus urticae and other Important Acari: A review. Insect Biochem. Mol. Biol. 2010, 40, 563–572. [Google Scholar] [CrossRef]
- Iida, J.; Desaki, Y.; Hata, K.; Uemura, T.; Yasuno, A.; Islam, M.; Maffei, M.E.; Ozawa, R.; Nakajima, T.; Galis, I.; et al. Tetranins: New Putative Spider Mite Elicitors of Host Plant Defense. New Phytol. 2019, 224, 875–885. [Google Scholar] [CrossRef]
- Sedaratian, A.; Fathipour, Y.; Moharramipour, S. Comparative Life Table Analysis of Tetranychus urticae (Acari: Tetranychidae) on 14 Soybean Genotypes. Insect. Sci. 2011, 18, 541–553. [Google Scholar] [CrossRef]
- Silva, P. Um novo acaro nocivo ao tomateiro na Bahia. Bol. Inst. Biol. Bahia 1954, 1, 1–20. [Google Scholar]
- Moutia, L.A. Contribution to the study of some phytophagous Acarina and their predators in Mauritius. Bull. Entomol. Res. 1958, 49, 59–75. [Google Scholar] [CrossRef]
- Blair, B.W. Tetranychus evansi Baker & Pritchard (Acari: Tetranychidae); a New Pest of Tobacco in Zimbabwe; Coresta Phytopathology and Agronomy Study Group: Bergerac, France, 1983; pp. 1–6. [Google Scholar]
- de Moraes, G.J.; McMurtry, J.A. Comparison of Tetranychus evansi and T. urticae (Acari: Tetranychidae) as Prey for Eight Species of Phytoseiid Mites. Entomophaga 1985, 30, 393–397. [Google Scholar] [CrossRef]
- Chen, J.; Zheng, L.; Ye, Z.; Wang, J.; Zhang, F.; Fu, Y.; Zhang, C. Evaluation of the Predatory Mite Neoseiulus barkeri against Spider Mites Damaging Rubber Trees. Insects 2023, 14, 648. [Google Scholar] [CrossRef] [PubMed]
- Van Lenteren, J.C. The State of Commercial Augmentative Biological Control: Plenty of Natural Enemies, but a Frustrating Lack of Uptake. BioControl 2012, 57, 1–20. [Google Scholar] [CrossRef]
- Opit, G.; Nechols, J.; Margolies, D. Biological Control of Twospotted Spider mites, Tetranychus urticae Koch (Acari: Tetranychidae), using Phytoseiulus persimilis Athias-Henriot (Acari: Phytoseidae) on Ivy Geranium: Assessment of Predator Release Ratios. Biol. Control 2004, 29, 445–452. [Google Scholar] [CrossRef]
- Opit, G.P. Comparing Chemical and Biological Control Strategies for Twospotted Spider Mites (Acari: Tetranychidae) in Commercial Greenhouse Production of Bedding Plants. J. Econ. Entomol. 2009, 102, 336–346. [Google Scholar] [CrossRef]
- de Moraes, G.J.; McMurtry, J.A. Suitability of the Spider Mite Tetranychus evansi as Prey for Phytoseiulus persimilis. Entomol. Exp. Appl. 1986, 40, 109–115. [Google Scholar] [CrossRef]
- Grostal, R.; O’Dowd, D.J. Plants, Mites and Mutualism: Leaf Domatia and the Abundance and Reproduction of Mites on Viburnum tinus (Caprifoliaceae). Oecologia 1994, 97, 308–315. [Google Scholar] [CrossRef]
- Allahyari, H.; Fard, P.A.; Nozari, J. Effects of Host on Functional Response of Offspring in Two Populations of Trissolcus grandis, on the Sunn Pest. J. Appl. Entomol. 2004, 128, 39–43. [Google Scholar] [CrossRef]
- Collier, K.F.S.; Albuquerque, G.S.; Lima, J.O.G.D.; Pallini, A.; Molina-Rugama, A.J. Neoseiulus idaeus (Acari: Phytoseiidae) as a Potential Biocontrol Agent of the Two-spotted Spider Mite, Tetranychus urticae (Acari: Tetranychidae) in Papaya: Performance on Different Prey Stage-host Plant Combinations. Exp. Appl. Acarol. 2007, 41, 27–36. [Google Scholar] [CrossRef]
- Buitenhuis, R.; Shipp, L.; Scott-Dupree, C.; Brommit, A.; Lee, W. Host Plant Effects on the Behaviour and Performance of Amblyseius swirskii (Acari: Phytoseiidae). Exp. Appl. Acarol. 2014, 62, 171–180. [Google Scholar] [CrossRef]
- Zhang, X.X.; Lv, J.L.; Hu, Y.; Wang, B.M.; Chen, X.; Xu, X.N.; Wang, E.D. Prey Preference and Life Table of Amblyseius orientalis on Bemisia tabaci and Tetranychus cinnabarinus. PLoS ONE 2015, 10, e0138820. [Google Scholar] [CrossRef]
- Elliott, J.M. Prey Switching in Rhyacophila dorsalis (Trichoptera) Alters with Larval Instar. Freshw. Biol. 2006, 51, 913–924. [Google Scholar] [CrossRef]
- Thao, N.; Thuy, N.T.; Quyen, H.L. Effects of Different Diets on Biological Characteristics of Predatory Mite Amblyseius eharai (Acari: Phytoseiidae). Insects 2023, 14, 519. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Zhang, C.H.; Yi, Z.j.; Ran, X.C.; Zhang, L.S.; Liu, C.X.; Wang, M.Q.; Chen, H.Y. Effects of Three Prey Species on Development and Fecundity of the Predaceous Stinkbug Arma chinensis (Hemiptera: Pentatomidae). Chin. J. Bio Control 2016, 32, 553–561. [Google Scholar] [CrossRef]
- Wang, J.J.; Zhang, K.S.; Li, L.; Zhang, Z.Q. Development and Reproduction of Four Predatory Mites (Parasitiformes: Phytoseiidae) Feeding on the Spider Mites Tetranychus evansi and T. urticae (Trombidiformes: Tetranychidae) and the Dried. Fruit Mite Carpoglyphus lactis (Sarcoptiformes: Carpoglyphidae). Syst. Appl. Acarol. 2024, 29, 269–284. [Google Scholar] [CrossRef]
- Schmidt, R.A. Leaf Structures Affect Predatory Mites (Acari: Phytoseiidae) and Biological Control: A Review. Exp. Appl. Acarol. 2014, 62, 1–17. [Google Scholar] [CrossRef]
- Buitenhuis, R.; Murphy, G.; Shipp, L.; Scott-Dupree, C. Amblyseius swirskii in Greenhouse Production Systems: A Floricultural Perspective. Exp. Appl. Acarol. 2015, 65, 451–464. [Google Scholar] [CrossRef]
- Zhang, Y.N.; Guo, D.D.; Jiang, J.Y.Q.; Zhang, Y.J.; Zhang, J.P. Effects of Host Plant Species on the Development and Reproduction of Neoseiulus bicaudus (Phytoseiidae) Feeding on Tetranychus turkestani (Tetranychidae). Syst. Appl. Acarol. 2016, 21, 647. [Google Scholar] [CrossRef]
- Paspati, A.; Rambla, J.L.; López Gresa, M.P.; Arbona, V.; Gómez-Cadenas, A.; Granell, A.; González-Cabrera, J.; Urbaneja, A. Tomato Trichomes are Deadly Hurdles Limiting the Establishment of Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae). Biol. Control 2021, 157, 104572. [Google Scholar] [CrossRef]
- Buonomo, B.; Giannino, F.; Stéphanie, S.; Venturino, E. Effects of Limited Volatiles Release by Plants in Tritrophic Interactions. Math. Biosci. Eng. 2019, 16, 3331–3344. [Google Scholar] [CrossRef]
- Birch, L. The Intrinsic Rate of Natural Increase of an Insect Population. J. Anim. Ecol. 1948, 17, 15–26. [Google Scholar] [CrossRef]
- Furtado, I.P.; Toledo, S.; Moraes, G.J.; Kreiter, S.; Knapp, M. Search for Effective Natural Enemies of Tetranychus evansi (Acari: Tetranychidae) in Northwest Argentina. Exp. Appl. Acarol. 2007, 43, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Sarmento, R.A.; Oliveira, H.G.; Holtz, A.M.; Da Silva, S.M.; Serrao, J.E.; Pallini, A. Fat Body Morphology of Eriopis connexa (Coleoptera, Coccinelidae) in Function of Two Alimentary Sources. Braz. Arch. Biol. Technol. 2004, 47, 407–411. [Google Scholar] [CrossRef]
- Oliveira, E.E.; Oliveira, C.L.; Sarmento, R.A.; Fadini, M.A.M.; Moreira, L.R. Biological Aspects of the Predator Cycloneda sanguinea (Linnaeus, 1763) (Coleoptera: Coccinellidae) Fed with Tetranychus evansi (Baker and Pritchard, 1960) (Acari: Tetranychidae) and Macrosiphum euphorbiae (Thomas, 1878) (Homoptera: Aphididae) [Aspectos biologicos do predador Cycloneda sanguinea (Linnaeus, 1763) (Coleoptera: Coccinellidae) Alimentado com Tetranychus evansi (Baker e Pritchard, 1960) (Acari: Tetranychidae) e Macrosiphum euphorbiae (Thomas, 1878) (Homoptera: Aphididae)]. Biosci. J. 2005, 21, 33–39. [Google Scholar]
- Navajas, M.; Moraes, G.J.; Auger, P.; Migeon, A. Review of the invasion of Tetranychus evansi: Biology, Colonization Pathways, Potential Expansion and Prospects for Biological Control. Exp. Appl. Acarol. 2013, 59, 43–65. [Google Scholar] [CrossRef]
- de Moraes, G.J.; McMurtry, J.A. Chemically Mediated Arrestment of the Predaceous Mite Phytoseiulus persimilis by Extracts of Tetranychus evansi and Tetranychus urticae. Exp. Appl. Acarol. 1985, 1, 127–138. [Google Scholar] [CrossRef]
- Gotoh, T.; Tsuchiya, A.; Kitashima, Y. Influence of Prey on Developmental Performance, Reproduction and Prey. Consumption of Neoseiulus californicus (Acari: Phytoseiidae). Exp. Appl. Acarol. 2006, 40, 189–204. [Google Scholar] [CrossRef]
- de Moraes, G.J.; McMurtry, J.A. Physiological Effect of the Host Plant on the Suitability of Tetranychus urticae as Prey for Phytoseiulus persimilis. Entomphaga 1987, 32, 35–38. [Google Scholar] [CrossRef]
- Escudero, L.A.; Ferragut, F. Life-history of Predatory Mites Neoseiulus californicus and Phytoseiulus persimilis (Acari: Phytoseiidae) on Four Spider Mite Species as Prey, with Special Reference to Tetranychus evansi (Acari: Tetranychidae). Biol. Control 2005, 32, 378–384. [Google Scholar] [CrossRef]
- Van Geem, M.; Harvey, J.A.; Gols, R. Development of a Generalist Predator, Podisus maculiventris, on Glucosinolate Sequestering and Nonsequestering Prey. Naturwissenschaften 2014, 101, 707–714. [Google Scholar] [CrossRef]
- Wiklund, C.; Friberg, M. Enemy-free Space and Habitat-specific Host Specialization in a Butterfly. Occologia 2008, 157, 287–294. [Google Scholar] [CrossRef]
- Holzinger, F.; Wink, M. Mediation of Cardiac Glycoside Insensitivity in the Monarch butterfly (Danaus plexippus): Role of an Amino Acid Substitution in the Ouabain Binding Site of Na+K+-ATPase. J. Chem. Ecol. 1996, 22, 1921–1937. [Google Scholar] [CrossRef]
- Fordyce, J.A. The Lethal Plant Defence Paradox Remains: Inducible Host-plant Aristolochic Acids and The Growth and Defence of the Pipevine swallowtail. Exp. Appl. Acarol. 2001, 100, 339–346. [Google Scholar] [CrossRef]
- Patra, B.; Schluttenhofer, C.; Wu, Y.M.; Pattanaik, S.; Yuan, L. Transcriptional regulation of secondary metabolite biosynthesis in plants. Biochim. Biophys. Acta 2013, 1829, 1236–1247. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.