

Effects of Trehalase on the Gene Expression of the Reproductive Regulation Pathway Network and Triglyceride Metabolism in Nilaparvata lugens (Stål)

 and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. dsRNA Synthesis and Microinjection
2.3. qRT-PCR
2.4. Observation of Abdomen and Detection of Triglyceride Content
2.5. Data Analysis
3. Results
3.1. Effects of Trehalase on Abdominal Volumes, Vg, and VgR Expression
3.2. Effects of Trehalase on JH and 20E Signaling Pathways
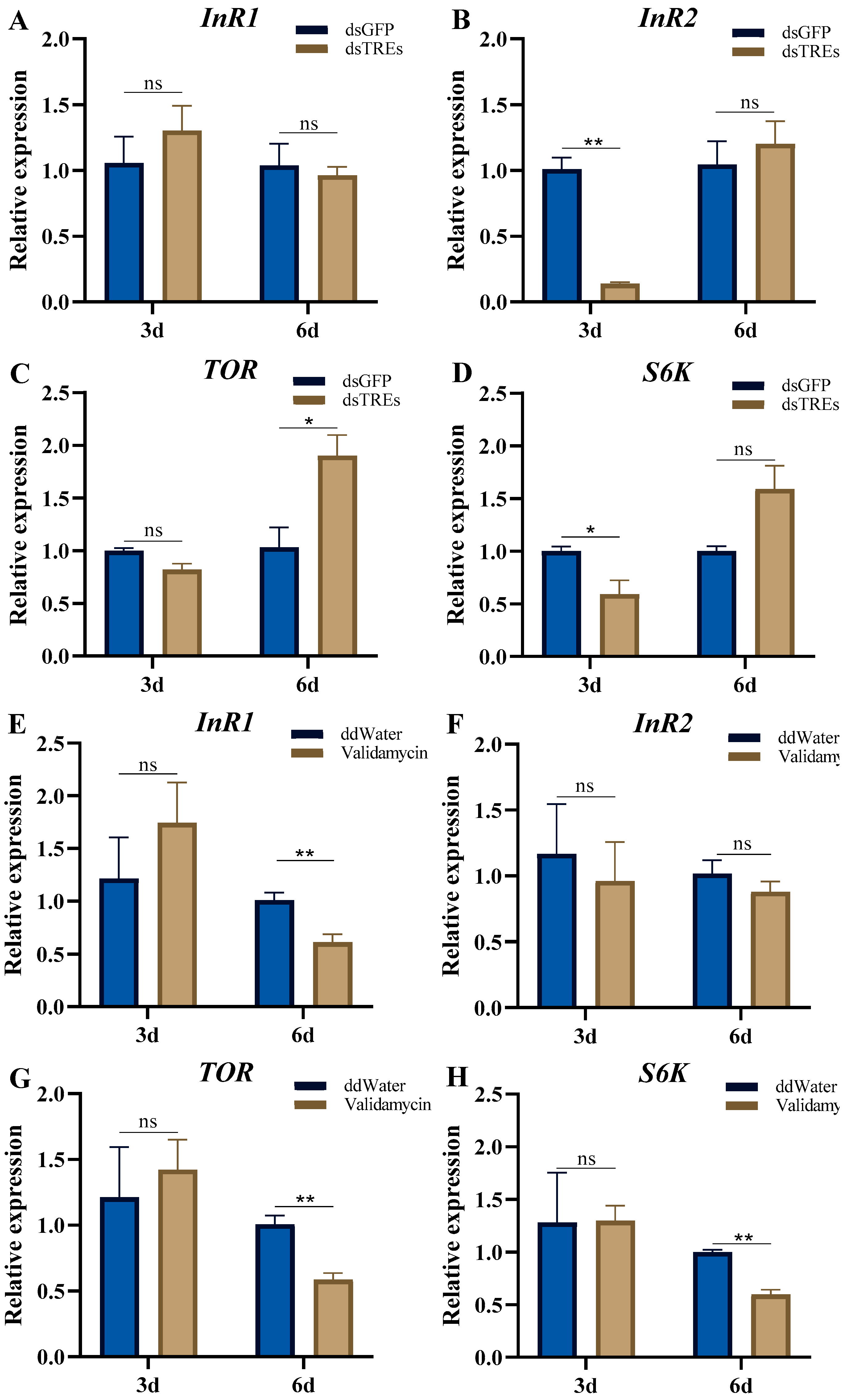
3.3. Effects of Trehalase on Nutrient Signaling Pathways
3.4. Effects of Trehalase on Lipid Metabolism
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Xu, H.J.; Zhang, C.X. Insulin receptors and wing dimorphism in rice planthoppers. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20150489. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.M.; Zhang, H.H.; Gao, H.L.; Yuan, X.W.; Liu, Z.W. The transcription factor CREB3-2 regulated neutral lipase gene expression in ovary of Nilaparvata lugens. Pestic. Biochem. Physiol. 2023, 196, 105632. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.H.; Yang, B.J.; Xu, X.X.; Zou, J.Z.; Liu, Z.W. Roles of spermary-specific carboxylesterases in Nilaparvata lugens reproduction: Opposite between insecticide-induced upregulation and resistance-associated overexpression. Int. J. Biol. Macromol. 2024, 290, 138991. [Google Scholar] [CrossRef]
- Xia, X.; Luo, G.H.; Yu, J.L.; Liu, B.Q.; Zhang, R.; Zhang, G.; Shu, Z.L.; Lou, Y.G.; Hoffmann, A.A.; Fang, J.C. Jasmonic acid changes associated with long-term control of lepidopteran rice pests after insecticide seed treatment. Entomol. Gen. 2024, 44, 573–581. [Google Scholar] [CrossRef]
- Yang, B.; Lin, X.; Yu, N.; Gao, H.; Zhang, Y.; Liu, W.; Liu, Z. Contribution of glutathione s-transferases to imidacloprid resistance in Nilaparvata lugens. J. Agric. Food Chem. 2020, 68, 15403–15408. [Google Scholar] [CrossRef]
- Tang, B.; Xu, K.; Liu, Y.; Zhou, Z.; Karthi, S.; Yang, H.; Li, C. A review of physiological resistance to insecticide stress in Nilaparvata lugens. 3 Biotech 2022, 12, 84. [Google Scholar] [CrossRef]
- Wang, L.X.; Tao, S.; Zhang, Y.C.; Pei, X.G.; Gao, Y.; Song, X.Y.; Yu, Z.T.; Gao, C.F. Overexpression of ATP-binding cassette transporter Mdr49-like confers resistance to imidacloprid in the field populations of brown planthopper, Nilaparvata lugens. Pest Manag. Sci. 2022, 78, 579–590. [Google Scholar] [CrossRef]
- Huang, J.H.; Lee, H.J. RNA interference unveils functions of the hypertrehalosemic hormone on cyclic fluctuation of hemolymph trehalose and oviposition in the virgin female Blattella germanica. J. Insect Physiol. 2011, 57, 858–864. [Google Scholar] [CrossRef]
- Tang, B.; Hu, S.R.; Luo, Y.J.; Shi, D.M.; Liu, X.Y.; Zhong, F.; Jiang, X.Y.; Hu, G.; Li, C.; Duan, H.X.; et al. Impact of Three Thiazolidinone Compounds with Piperine Skeletons on Trehalase Activity and Development of Spodoptera frugiperda Larvae. J. Agric. Food Chem. 2024, 72, 8423–8433. [Google Scholar] [CrossRef]
- Li, Y.; Wang, S.S.; Si, H.R.; Wan, S.J.; Li, G.Y.; Shu, Y.H.; Dai, X.Y.; Wang, R.J.; Wang, S.G.; Zhai, Y.F.; et al. Responses of aphid and ladybird to lead transfer through soil and broad beans. Entomol. Gen. 2024, 44, 621–623. [Google Scholar] [CrossRef]
- Avonce, N.; Mendoza-Vargas, A.; Morett, E.; Morett, E.; Iturriaga, G. Insights on the evolution of trehalose biosynthesis. BMC Evol. Biol. 2006, 6, 109. [Google Scholar] [CrossRef]
- Mitsumasu, K.; Azuma, M.; Niimi, T.; Yamashita, O.; Yaginuma, T. Membrane-penetrating trehalase from Bombyx mori. Molecular cloning and localization in larval midgut. Insect Mol. Biol. 2005, 14, 501–508. [Google Scholar] [PubMed]
- Lee, J.H.; Saito, S.; Mori, H.; Nishimoto, M.; Okuyama, M.; Kim, D.; Wongchawalit, J.; Kimura, A.; Chiba, S. Molecular cloning of cDNA for trehalase from the European honeybee, Apis mellifera L., and its heterologous expression in Pichia pastoris. Biosci. Biotechnol. Biochem. 2007, 71, 2256–2265. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Wei, P.; Zhao, L.N.; Shi, Z.; Shen, Q.; Yang, M.; Xie, G.; Wang, S. Knockdown of five trehalase genes using RNA interference regulates the gene expression of the chitin biosynthesis pathways in Tribolium castaneum. BMC Biotechnol. 2016, 16, 67. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.N.; Yang, M.M.; Shen, Q.D.; Liu, X.J.; Shi, Z.K.; Wang, S.G.; Tang, B. Functional characterization of three trehalase genes regulating the chitin metabolism pathway in rice brown planthopper using RNA interference. Sci. Rep. 2016, 6, 27841. [Google Scholar] [CrossRef]
- Tang, B.; Yang, M.M.; Shen, Q.D.; Xu, Y.X.; Wang, H.J.; Wang, S.G. Suppressing the activity of trehalase with validamycin disrupts the trehalose and chitin biosynthesis pathways in the rice brown planthopper, Nilaparvata lugens. Pestic. Biochem. Physiol. 2017, 137, 81–90. [Google Scholar] [CrossRef]
- Tang, B.; Han, Y.; Mao, Q.X.; Fu, H.Y.; Luo, Y.J.; Hua, L.Y.H.; Liu, B.S.; Hu, G.; Wang, S.G.; Desneux, N.; et al. Regulation of three novel pepper thiothiazolidinones on the fecundity of Spodoptera frugiperda. Pestic. Biochem. Physiol. 2024, 204, 106033. [Google Scholar] [CrossRef]
- Matassini, C.; Parmeggiani, C.; Cardona, F. New Frontiers on human safe insecticides and fungicides: An opinion on trehalase inhibitors. Molecules 2020, 25, 3013. [Google Scholar] [CrossRef]
- Yu, H.Z.; Zhang, Q.; Lu, Z.J.; Deng, M.J. Validamycin treatment significantly inhibits the glycometabolism and chitin synthesis in the common cutworm, Spodoptera litura. Insect Sci. 2022, 29, 840–854. [Google Scholar] [CrossRef]
- Turna Demir, F.; Demir, E. In vivo evaluation of the neurogenotoxic effects of exposure to validamycin A in neuroblasts of Drosophila melanogaster larval brain. J. Appl. Toxicol. 2024, 44, 355–370. [Google Scholar] [CrossRef]
- Tatun, N.; Wangsantitham, O.; Tungitwitayakul, J.; Sakvrai, S. Trehalase activity in fungus-growing termite, Odontotermes feae (Isoptera: Termitideae) and inhibitory effect of validamycin. J. Econ. Entomol. 2014, 107, 1224–1232. [Google Scholar] [CrossRef]
- Tatun, N.; Tungjitwitayakul, J.; Sakurai, S. Developmental and Lethal Effects of Trehalase Inhibitor (Validamycin) on the Tribolium castaneum (Coleoptera: Tenebrionidae). Ann. Entomol. Soc. Am. 2016, 109, 224–231. [Google Scholar] [CrossRef]
- Wangsantitham, O.; Pothong, P.; Tungjitwitayakul, J.; Tatun, N. Suppression of Trehalase Activity by Validamycin Induces Mortality and Developmental Delays in Sitophilus zeamais (Coleoptera: Curculionidae). J. Entomol. Sci. 2024, 59, 311–322. [Google Scholar] [CrossRef]
- Marten, A.D.; Stothard, A.I.; Kalera, K.; Swarts, B.M.; Conway, M.J. Validamycin A Delays Development and Prevents Flight in Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2020, 57, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Zuo, H.; Gao, L.; Yi, X.; Zhong, G. Silencing of rieske iron-sulfur protein impacts upon the development and reproduction of Spodoptera exigua by regulating ATP synthesis. Front. Physiol. 2018, 9, 575. [Google Scholar] [CrossRef]
- Tufail, M.; Takeda, M. Molecular characteristics of insect vitellogenins. J. Insect Physiol. 2008, 54, 1447–1458. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Chen, Y.Z.; Lou, Y.H.; Zhang, C.X. Vitellogenin and vitellogenin-like genes in the brown planthopper. Front. Physiol. 2019, 10, 1181. [Google Scholar] [CrossRef]
- Wu, Z.X.; Yang, L.B.; He, Q.J.; Zhou, S.T. Regulatory mechanisms of vitellogenesis in insects. Front. Cell Dev. Biol. 2021, 8, 593613. [Google Scholar] [CrossRef]
- Zou, Z.; Saha, T.T.; Roy, S.; Shin, S.W.; Backman, T.W.; Girke, T.; White, K.P.; Raikhel, A.S. Juvenile hormone and its receptor, methoprene-tolerant, control the dynamics of mosquito gene expression. Proc. Natl. Acad. Sci. USA 2013, 110, E2173–E2181. [Google Scholar] [CrossRef]
- Hansen, I.A.; Attardo, G.M.; Rodriguez, S.D.; Drake, L.L. Four-way regulation of mosquito yolk protein precursor genes by juvenile hormone-, ecdysone-, nutrient-, and insulin-like peptide signaling pathways. Front. Physiol. 2014, 5, 103. [Google Scholar] [CrossRef]
- Roy, S.; Saha, T.T.; Zou, Z.; Raikhel, A.S. Regulatory pathways controlling female insect reproduction. Annu. Rev. Entomol. 2018, 63, 489–511. [Google Scholar] [CrossRef]
- Wu, Z.; Guo, W.; Yang, L.; He, Q.; Zhou, S. Juvenile hormone promotes locust fat body cell polyploidization and vitellogenesis by activating the transcription of Cdk6 and E2f1. Insect Biochem. Mol. Biol. 2018, 102, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.G.; Humann, F.C.; Hartfelder, K. Juvenile hormone signaling in insect oogenesis. Curr. Opin. Insect Sci. 2019, 31, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.; Raikhel, A.S. Cross-talk of insulin-like peptides, juvenile hormone, and 20-hydroxyecdysone in regulation of metabolism in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2021, 118, e2023470118. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Liu, S.; Zhang, W.; Yang, L.; Huang, J.; Zhou, S.; Feng, Q.; Palli, S.R.; Wang, J.; Roth, S.; et al. Juvenile hormone signaling promotes ovulation and maintains egg shape by inducing expression of extracellular matrix genes. Proc. Natl. Acad. Sci. USA 2021, 118, e2104461118. [Google Scholar] [CrossRef]
- Wu, M.Z.; Fan, S.T.; Zhang, Y.C.; Tan, J.F.; Zhu, G.H. Disrupting shadow in the prothoracic gland induced larval development arrest in the fall armyworm Spodoptera frugiperda. Front. Physiol. 2024, 15, 1502753. [Google Scholar] [CrossRef]
- Ekoka, E.; Maharaj, S.; Nardini, L.; Dahan-Moss, Y.; Koekemoer, L.L. 20-Hydroxyecdysone (20E) signaling as a promising target for the chemical control of malaria vectors. Parasites Vectors 2021, 14, 86. [Google Scholar] [CrossRef]
- Wu, Q.; Brown, M.R. Signaling and function of insulin-like peptides in insects. Annu. Rev. Entomol. 2006, 51, 1–24. [Google Scholar] [CrossRef]
- Kannan, K.; Fridell, Y.W. Functional implications of Drosophila insulin-like peptides in metabolism, aging, and dietary restriction. Front. Physiol. 2013, 4, 288. [Google Scholar] [CrossRef]
- Dong, Y.; Chen, W.; Kang, K.; Pang, R.; Dong, Y.; Liu, K.; Zhang, W. FoxO directly regulates the expression of TOR/S6K and vitellogenin to modulate the fecundity of the brown planthopper. Sci. China Life Sci. 2021, 64, 133–143. [Google Scholar] [CrossRef]
- Huangfu, N.; Zhu, X.; Wang, L.; Zhang, K.; Li, D.; Chen, L.; Gao, X.; Niu, L.; Gao, M.; Ji, J.; et al. Insulin receptor substrate-1 (IRS1) regulates oogenesis and vitellogenesis in Propylea japonica by Mediating the FOXO transcription factor expression, independent of JH and 20E signaling pathways. J. Agric. Food Chem. 2023, 71, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Rauschenbach, I.Y.; Karpova, E.K.; Burdina, E.V.; Adonyeva, N.V.; Bykov, R.A.; Ilinsky, Y.Y.; Menshanov, P.N.; Gruntenko, N.E. Insulin-like peptide DILP6 regulates juvenile hormone and dopamine metabolism in Drosophila females. Gen. Comp. Endocrinol. 2017, 243, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Smykal, V.; Raikhel, A.S. Nutritional control of insect reproduction. Curr. Opin. Insect Sci. 2015, 11, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Katewa, S.D.; Kapahi, P. Role of TOR signaling in aging and related biological processes in Drosophila melanogaster. Exp. Gerontol. 2011, 46, 382–390. [Google Scholar] [CrossRef]
- Lu, K.; Chen, X.; Liu, W.T.; Zhang, X.Y.; Chen, M.X.; Zhou, Q. Nutritional signaling regulates vitellogenin synthesis and egg development through juvenile hormone in Nilaparvata lugens (Stål). Int. J. Mol. Sci. 2016, 17, 269. [Google Scholar] [CrossRef]
- Lu, K.; Chen, X.; Liu, W.T.; Zhou, Q. TOR pathway-mediated juvenile hormone synthesis regulates nutrient-dependent female reproduction in Nilaparvata lugens (Stål). Int. J. Mol. Sci. 2016, 17, 438. [Google Scholar] [CrossRef]
- Lu, K.; Chen, X.; Li, W.; Li, Y.; Zhang, Z.; Zhou, Q. Insulin-like peptides and DNA/tRNA methyltransferases are involved in the nutritional regulation of female reproduction in Nilaparvata lugens (Stål). Gene 2018, 639, 96–105. [Google Scholar] [CrossRef]
- Zhou, X.; Ye, Y.Z.; Ogihara, M.H.; Takeshima, M.; Fujinaga, D.; Liu, C.W.; Zhu, Z.; Kataoka, H.; Bao, Y.Y. Functional analysis of ecdysteroid biosynthetic enzymes of the rice planthopper, Nilaparvata lugens. Insect Biochem. Mol. Biol. 2020, 123, 103428. [Google Scholar] [CrossRef]
- Liu, Y.K.; Zhu, Y.; Wan, S.J.; Wang, X.Z.; Guan, L.W.; Xu, C.D.; Xie, B.H.; Wang, S.G.; Sun, S.S.; Tang, B. Trehalase regulates ovarian maturation and egg hatchability of Nilaparvata lugens (Stål). J. Pest Sci. 2025, 31, 1. [Google Scholar]
- Xu, H.J.; Xue, J.; Lu, B.; Zhang, X.C.; Zhuo, J.C.; He, S.F.; Ma, X.F.; Jiang, Y.Q.; Fan, H.W.; Xu, J.Y.; et al. Two insulin receptors determine alternative wing morphs in planthoppers. Nature 2015, 519, 464–467. [Google Scholar] [CrossRef]
- Li, L.; Jiang, Y.; Liu, Z.; You, L.; Wu, Y.; Xu, B.; Ge, L.; Stanley, D.; Song, Q.; Wu, J. Jinggangmycin increases fecundity of the brown planthopper, Nilaparvata lugens (Stål) via fatty acid synthase gene expression. J. Proteom. 2016, 130, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Chen, X.; Li, Y.; Li, W.; Zhou, Q. Lipophorin receptor regulates Nilaparvata lugens fecundity by promoting lipid accumulation and vitellogenin biosynthesis. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2018, 219–220, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Shu, Y.; Zhou, J.; Zhang, X.; Zhang, X.; Chen, M.; Yao, Q.; Zhou, Q.; Zhang, W. Molecular characterization and RNA interference analysis of vitellogenin receptor from Nilaparvata lugens (Stål). J. Insect Physiol. 2015, 73, 20–29. [Google Scholar] [CrossRef]
- Li, Y.N.; Liu, Y.B.; Xie, X.Q.; Zhang, J.N.; Li, W.L. The modulation of trehalose metabolism by 20-hydroxyecdysone in Antheraea pernyi (Lepidoptera: Saturniidae) during its diapause termination and post-termination period. J. Insect Sci. 2020, 20, 18. [Google Scholar] [CrossRef]
- He, Y.N.; Bu, T.L.; Liu, X.D. Tendency to mate with short-winged partner of the brown planthopper Nilaparvata lugens (Hemiptera: Delphacidae). J. Econ. Entomol. 2024, 117, 2425–2432. [Google Scholar] [CrossRef]
- Xue, W.H.; Xu, N.; Chen, S.J.; Liu, X.Y.; Zhang, J.L.; Xu, H.J. Neofunctionalization of a second insulin receptor gene in the wing-dimorphic planthopper, Nilaparvata lugens. PLoS Genet. 2021, 17, e1009653. [Google Scholar] [CrossRef]
- Xue, W.H.; Liu, Y.L.; Jiang, Y.Q.; He, S.F.; Wang, Q.Q.; Yang, Z.N.; Xu, H.J. Molecular characterization of insulin-like peptides in the brown planthopper, Nilaparvata lugens (Hemiptera: Delphacidae). Insect Mol. Biol. 2020, 29, 309–319. [Google Scholar] [CrossRef]
- Ziegler, R.; Van Antwerpen, R. Lipid uptake by insect oocytes. Insect Biochem. Mol. Biol. 2006, 36, 264–272. [Google Scholar] [CrossRef]
- Athenstaedt, K.; Daum, G. The life cycle of neutral lipids: Synthesis, storage and degradation. Cell. Mol. Life Sci. 2006, 63, 1355–1369. [Google Scholar] [CrossRef]
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef]
- Wang, W.; Yang, R.R.; Peng, L.Y.; Zhang, L.; Yao, Y.L.; Bao, Y.Y. Proteolytic activity of the proteasome is required for female insect reproduction. Open Biol. 2021, 11, 200251. [Google Scholar] [CrossRef]
- Wang, W.; Ma, Y.; Yang, R.R.; Cheng, X.; Huang, H.J.; Zhang, C.X.; Bao, Y.Y. An MD-2-related lipid-recognition protein is required for insect reproduction and integument development. Open Biol. 2021, 11, 210170. [Google Scholar] [CrossRef]
- Lu, K.; Wang, Y.; Chen, X.; Zhang, X.; Li, W.; Cheng, Y.; Li, Y.; Zhou, J.; You, K.; Song, Y.; et al. Adipokinetic hormone receptor mediates trehalose homeostasis to promote vitellogenin uptake by oocytes in Nilaparvata lugens. Front. Physiol. 2019, 9, 1904. [Google Scholar] [CrossRef]
- Arai, C.; Suyama, A.; Arai, S.; Arai, N.; Yoshizane, C.; Koya-Miyata, S.; Mizote, A.; Endo, S.; Ariyasu, T.; Mitsuzumi, H.; et al. Trehalose itself plays a critical role on lipid metabolism: Trehalose increases jejunum cytoplasmic lipid droplets which negatively correlated with mesenteric adipocyte size in both HFD-fed trehalase KO and WT mice. Nutr. Metab. 2020, 17, 22. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Accession Number | Primer Sequences (5′-3′) | Length | |
|---|---|---|---|---|
| NlTRE1-1 | FJ790319.1 | F: GATGCAATCAAGGAGGTGTTATGGC | R: CGTATTCACCTCCACCTCCGT | 451 bp |
| NlTRE1-2 | KU556829.1 | F: AGATGAAGGCATGTGGTTCG | R: CATCGATTCGCCAACTGGTAAGC | 321 bp |
| NlTRE2 | GQ397451.1 | F: CCAACTGCTATGACACCGACAAG | R: GGGTTCAGATCCTGCCGTCGCT | 440 bp |
| GFP | MW987535.2 | F: AAGGGCGAGGAGCTGTTCACCG | R: CAGCAGGACCATGTGATCGCGC | 720 bp |
| Primer Name | Accession Number | Primer Sequences (5′-3′) | |
|---|---|---|---|
| QNlActin | EU179847.1 | F: TGGACTTCGAGCAGGAAATGG | R: ACGTCGCACTTCATGATCGAG |
| QNlVg | JF330416.1 | F: CACTGCCCGTGCTGTGCTCTA | R: TGACTTCCTTGCTTTGCTCCC |
| QNlVgR | JQ040014.1 | F: AGGCAGCCACACAGATAACCGC | R: AGCCGCTCGCTCCAGAACATT |
| QNlJHAMT | KP769805.1 | F: GAACCTGCAGGCCAAACACA | R: ACCACTCGGTTGGGCTGAAT |
| QNlMet | KP797880.1 | F: AGTGGCAGCGAGCGATGATT | R: TGAGGCGCAGCAAAAAGGAG |
| QNlUSP | KX431887.1 | F: GGTGGAGCTGCTGAGGGAGA | R: AGCACTTGAGGCCGATGGAG |
| QNlEcR | FJ263049.1 | F: CGAAGCCTGGAAGGTGGAGA | R: GGCAAAGATTGGCGACGATT |
| QNlInR1 | KF974333.1 | F: GAGTGCAACCCGGAGTATGT | R: TCTTGACGGCACACTTCTTG |
| QNlInR2 | KF974334.1 | F: CTCTTGCCGAACAGCCTTAC | R: GGGTCGTTTAGTGGGTCTGA |
| QNlTOR | JQ793898.1 | F: GGCTACAGGGATGTCAAA | R: GAGATAGATTCAAACGGAAAG |
| QNlS6K | KP769804.1 | F: AATCGGACGACTTGGAGACAGT | R: CAGTTTGGAAAGCGTACATCAGG |
| QNlAKH | AB817235.1 | F: CCCTTCTGATGGCAGTCCTTTG | R: ATGGATGCCTTGCAGCCTTCT |
| QNlFas | XM_022339189.2 | F: CGGAGACTCTGCCCTAA | R: CAGCGACTAATCCAACATC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, B.; Ge, Y.; Liu, Y.; Guan, L.; Han, Y.; Zhu, Y.; Hu, G.; Wu, Y. Effects of Trehalase on the Gene Expression of the Reproductive Regulation Pathway Network and Triglyceride Metabolism in Nilaparvata lugens (Stål). Insects 2025, 16, 725. https://doi.org/10.3390/insects16070725
Tang B, Ge Y, Liu Y, Guan L, Han Y, Zhu Y, Hu G, Wu Y. Effects of Trehalase on the Gene Expression of the Reproductive Regulation Pathway Network and Triglyceride Metabolism in Nilaparvata lugens (Stål). Insects. 2025; 16(7):725. https://doi.org/10.3390/insects16070725
Chicago/Turabian StyleTang, Bin, Yuxin Ge, Yongkang Liu, Liwen Guan, Ye Han, Yang Zhu, Gao Hu, and Yan Wu. 2025. "Effects of Trehalase on the Gene Expression of the Reproductive Regulation Pathway Network and Triglyceride Metabolism in Nilaparvata lugens (Stål)" Insects 16, no. 7: 725. https://doi.org/10.3390/insects16070725
APA StyleTang, B., Ge, Y., Liu, Y., Guan, L., Han, Y., Zhu, Y., Hu, G., & Wu, Y. (2025). Effects of Trehalase on the Gene Expression of the Reproductive Regulation Pathway Network and Triglyceride Metabolism in Nilaparvata lugens (Stål). Insects, 16(7), 725. https://doi.org/10.3390/insects16070725