
Nosemosis in Russian Apis mellifera L. Populations: Distribution and Association with Hybridization

 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Determining the Origin of Bee Colonies
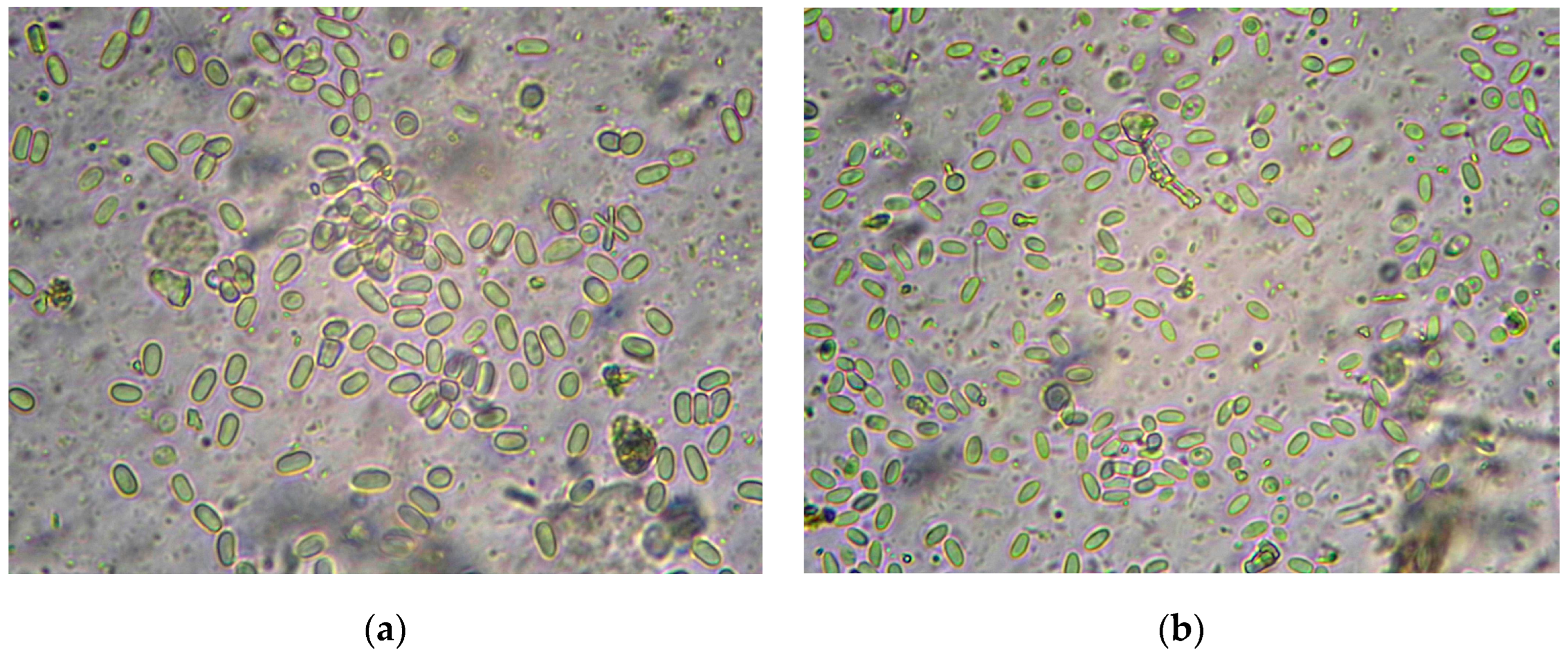
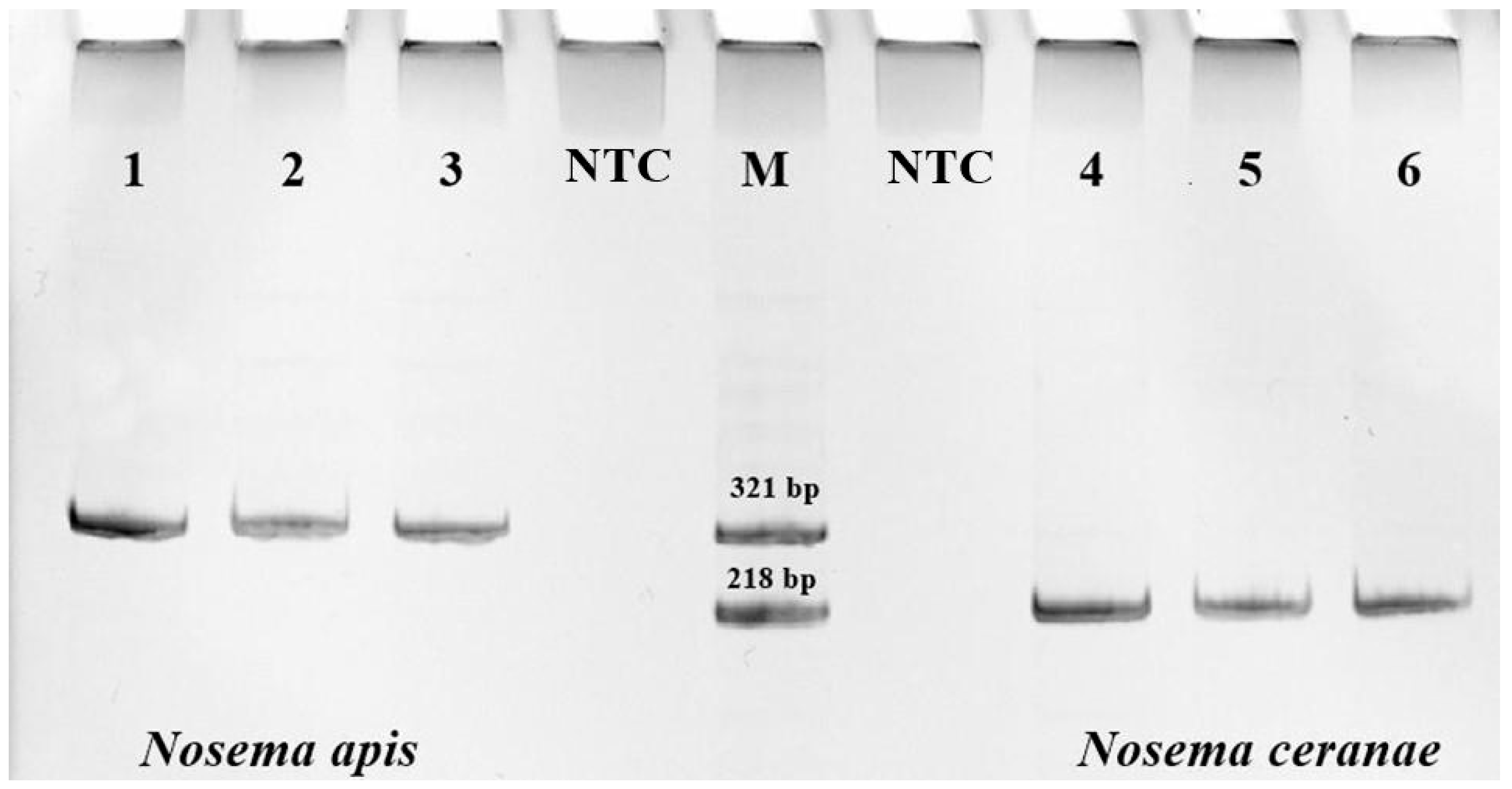
2.3. Microscopy and PCR Diagnostics of Nosemosis
3. Results
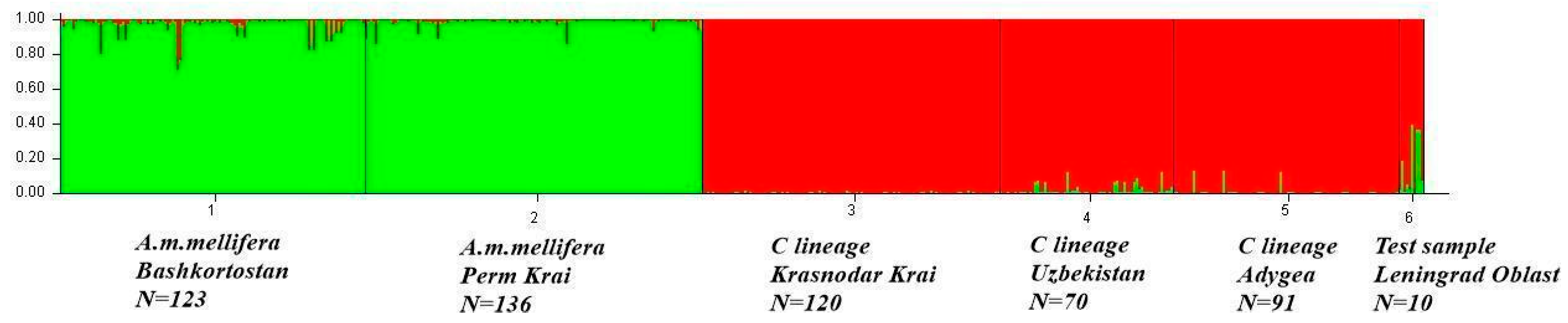
3.1. Genetic Structure of Apis mellifera Populations
3.2. Evaluation of the Relationship Between the Prevalence of Nosemosis and the Origin of Bees
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Giampieri, F.; Quiles, J.L.; Cianciosi, D.; Forbes-Hernández, T.Y.; Orantes Bermejo, F.J.; Alvarez-Suarez, J.M.; Battino, M. Bee products: An emblematic example of underutilized sources of bioactive compounds. J. Agric. Food Chem. 2022, 70, 6833–6848. [Google Scholar] [CrossRef]
- Ruttner, F. Biogeography and Taxonomy of Honeybees; Springer: Berlin, Germany, 1988; p. 291. [Google Scholar]
- Cridland, J.M.; Tsutsui, N.D.; Ramírez, S.R. The complex demographic history and evolutionary origin of the western honey bee, Apis mellifera. Genome Biol. Evol. 2017, 9, 457–472. [Google Scholar] [CrossRef] [PubMed]
- Momeni, J.; Parejo, M.; Nielsen, R.O.; Langa, J.; Montes, I.; Papoutsis, L.; Farajzadeh, L.; Bendixen, C.; Căuia, E.; Charrière, J.D.; et al. Authoritative subspecies diagnosis tool for European honey bees based on ancestry informative SNPs. BMC Genom. 2021, 22, 101. [Google Scholar] [CrossRef] [PubMed]
- Alpatov, V.V. Bee races and red clover pollination. Bee World 1948, 29, 61–63. [Google Scholar] [CrossRef]
- Alburaki, M.; Madella, S.; Lopez, J.; Bouga, M.; Chen, Y.; van Engelsdorp, D. Honey bee populations of the USA display restrictions in their mtDNA haplotype diversity. Front. Genet. 2023, 13, 1092121. [Google Scholar] [CrossRef]
- Kaskinova, M.D.; Gaifullina, L.R.; Saltykova, E.S. Haplotypes of the tRNAleu-COII mtDNA Region in Russian Apis mellifera Populations. Animals 2023, 13, 2394. [Google Scholar] [CrossRef] [PubMed]
- Ilyasov, R.A.; Lee, M.; Takahashi, J.; Kwon, H.W.; Nikolenko, A.G. A revision of subspecies structure of western honey bee Apis mellifera. Saudi J. Biol. Sci. 2020, 27, 3615–3621. [Google Scholar] [CrossRef]
- Frunze, O.; Brandorf, A.; Kang, E.-J.; Choi, Y.-S. Beekeeping Genetic Resources and Retrieval of Honey Bee Apis mellifera L. Stock in the Russian Federation: A Review. Insects 2021, 12, 684. [Google Scholar] [CrossRef]
- Ilyasov, R.A.; Kosarev, M.N.; Neal, A.; Yumaguzhin, F.G. Burzyan Wild-Hive Honeybee A. m. mellifera in South Ural. Bee World 2015, 92, 7–11. [Google Scholar] [CrossRef]
- Ilyasov, R.A.; Poskryakov, A.V.; Petukhov, A.V.; Nikolenko, A.G. Molecular genetic analysis of five extant reserves of black honeybee Apis mellifera mellifera in the Urals and the Volga Region. Russ. J. Genet. 2016, 52, 828–839. [Google Scholar] [CrossRef]
- Oleksa, A.; Tofilski, A. Wing geometric morphometrics and microsatellite analysis provide similar discrimination of honey bee subspecies. Apidologie 2015, 46, 49–60. [Google Scholar] [CrossRef]
- Ellis, J.S.; Soland-Reckeweg, G.; Buswell, V.G.; Huml, J.V.; Brown, A.; Knight, M.E. Introgression in native populations of Apis mellifera mellifera L.: Implications for conservation. J. Insect Conserv. 2018, 22, 377–390. [Google Scholar] [CrossRef]
- Orlovskytė, S.; Budrys, E.; Skrodenytė-Arbačiauskienė, V.; Blažytė-Čereškienė, L. The dark European honey bee Apis mellifera mellifera in Lithuania: Data on mitotype diversity of native bee population. J. Apic. Res. 2024, 64, 959–962. [Google Scholar] [CrossRef]
- Munoz, I.; Cepero, A.; Pinto, M.A.; Martin-Hernandez, R.; Higes, M.; De La Rua, P. Presence of Nosema ceranae associated with honeybee queen introduction. Infect. Genet. Evol. 2014, 23, 161–168. [Google Scholar] [CrossRef]
- Byatt, M.A.; Chapman, N.C.; Latty, T.; Oldroyd, B.P. The genetic consequences of the anthropogenic movement of social bees. Insectes Sociaux 2016, 63, 15–24. [Google Scholar] [CrossRef]
- Shamaev, N.D.; Shuralev, E.A.; Mukminov, M.N. Current status of Nosema spp. infection cases in Apis mellifera in eurasian countries and Ptp3 gene haplotypes in the Republic of Tatarstan, Russia. Vet. Res. Commun. 2024, 48, 2691–2698. [Google Scholar] [CrossRef]
- Ostap-Chec, M.; Cait, J.; Scott, R.W.; Arct, A.; Moroń, D.; Rapacz, M.; Miler, K. Nosemosis negatively affects honeybee survival: Experimental and meta-analytic evidence. Parasitology 2024, 15, 1530–1542. [Google Scholar] [CrossRef]
- Tokarev, Y.S.; Huang, W.F.; Solter, L.F.; Malysh, J.M.; Becnel, J.J.; Vossbrinck, C.R. A formal redefinition of the genera Nosema and Vairimorpha (Microsporidia: Nosematidae) and reassignment of species based on molecular phylogenetics. J. Invertebr. Pathol. 2020, 169, 107279. [Google Scholar] [CrossRef]
- Bartolomé, C.; Higes, M.; Hernández, R.M.; Chen, Y.P.; Evans, J.D.; Huang, Q. The recent revision of the genera Nosema and Vairimorpha (Microsporidia: Nosematidae) was flawed and misleads the bee scientific community. J. Invertebr. Pathol. 2024, 206, 108146. [Google Scholar] [CrossRef]
- Yoshiyama, M.; Kimura, K. Distribution of Nosema ceranae in the European honeybee, Apis mellifera in Japan. J. Invertebr. Pathol. 2011, 106, 263–267. [Google Scholar] [CrossRef]
- Lannutti, L.; Gonzales, F.N.; Dus Santos, M.J.; Florin-Christensen, M.; Schnittger, L. Molecular Detection and Differentiation of Arthropod, Fungal, Protozoan, Bacterial and Viral Pathogens of Honeybees. Vet. Sci. 2022, 9, 221. [Google Scholar] [CrossRef] [PubMed]
- Porrini, M.P.; Porrini, L.P.; Garrido, P.M.; Neto, S.; Porrini, D.P.; Muller, F.; Nuñez, L.A.; Alvarez, L.; Iriarte, P.F.; Eguaras, M.J. Nosema ceranae in South American native stingless bees and social wasps. Microb. Ecol. 2017, 74, 761–764. [Google Scholar] [CrossRef]
- Grupe, A.C., 2nd; Quandt, C.A. A growing pandemic: A review of Nosema parasites in globally distributed domesticated and native bees. PLoS Pathog. 2020, 18, e1008580. [Google Scholar] [CrossRef]
- Martín-Hernández, R.; Bartolome, C.; Chejanovsky, N.; Le Conte, Y.; Dalmon, A.; Dussaubat, C.; Dussaubat, C.; Meana, A.; Pinto, M.; Soroker, V.; et al. Nosema ceranae in Apis mellifera: A 12 years post-detection perspective. Environ. Microbiol. 2018, 20, 1302–1329. [Google Scholar] [CrossRef] [PubMed]
- Chauzat, M.P.; Higes, M.; Martín-Hernández, R.; Meana, A.; Cougoule, N.; Faucon, J.P. Presence of Nosema ceranae in French honey bee colonies. J. Apic. Res. 2007, 46, 127–128. [Google Scholar] [CrossRef]
- Higes, M.; Martın-Hernandez, R.; Meana, A. Nosema ceranae in Europe: An emergent type C nosemosis. Apidologie 2010, 41, 375–392. [Google Scholar] [CrossRef]
- Rangel, J.; Baum, K.; Rubink, W.L.; Coulson, R.N.; Johnston, J.S.; Traver, B.E. Prevalence of Nosema species in a feral honey bee population: A 20-year survey. Apidologie 2016, 47, 561–571. [Google Scholar] [CrossRef]
- Timofeev, S.A.; Ignatieva, A.N.; Dolgikh, V.V. Nosemosis type C of bees caused by Microsporidia Nosema (Vairimorpha) ceranae: Current views, pathogenesis, prevention, diagnosis and treatment. Agric. Biol. 2023, 58, 274–287. [Google Scholar] [CrossRef]
- Martín-Hernández, R.; Botías, C.; Bailón, E.G.; Martínez-Salvador, A.; Prieto, L.; Meana, A.; Higes, M. Microsporidia infecting Apis mellifera: Coexistence or competition. Is Nosema ceranae replacing Nosema apis? Environ. Microbiol. 2012, 14, 2127–2138. [Google Scholar] [CrossRef]
- Traver, B.E.; Fell, R.D. Prevalence and infection intensity of Nosema in honey bee (Apis mellifera L.) colonies in Virginia. J. Invertebr. Pathol. 2011, 107, 43–49. [Google Scholar] [CrossRef]
- Chen, Y.; Evans, J.D.; Smith, I.B.; Pettis, J.S. Nosema ceranae is a long-present and wide-spread microsporidian infection of the European honey bee (Apis mellifera) in the United States. J. Invertebr. Pathol. 2008, 97, 186–188. [Google Scholar] [CrossRef] [PubMed]
- Fries, I. Nosema ceranae in European honey bees (Apis mellifera). J. Invertebr. Pathol. 2010, 103, S73–S79. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.J.; Al-Ghamdi, A.; Nuru, A.; Khan, K.A.; Alattal, Y. Geographical distribution and molecular detection of Nosema ceranae from indigenous honey bees of Saudi Arabia. Saudi J. Biol. Sci. 2017, 24, 983–991. [Google Scholar] [CrossRef]
- Guerrero-Molina, C.; Correa-Benítez, A.; Hamiduzzaman, M.M.; Guzman-Novoa, E. Nosema ceranae is an old resident of honey bee (Apis mellifera) colonies in Mexico, causing infection levels of one million spores per bee or higher during summer and fall. J. Invertebr. Pathol. 2016, 141, 38–40. [Google Scholar] [CrossRef]
- Roberts, K.; Evison, S.; Baer, B.; Hughes, W.O.H. The cost of promiscuity: Sexual transmission of Nosema microsporidian parasites in polyandrous honey bees. Sci. Rep. 2015, 5, 10982. [Google Scholar] [CrossRef]
- Teixeira, É.W.; Guimarães-Cestaro, L.; Alves, M.L.T.M.F.; Message, D.; Martins, M.F.; Pinto da Luz, C.F.; Serrão, J.E. Spores of Paenibacillus larvae, Ascosphaera apis, Nosema ceranae and Nosema apis in bee products supervised by the Brazilian Federal Inspection Service. Rev. Bras. Entomol. 2018, 62, 188–194. [Google Scholar] [CrossRef]
- Galajda, R.; Valenčáková, A.; Sučik, M.; Kandráčová, P. Nosema disease of European honey bees. J. Fungus 2021, 7, 714. [Google Scholar] [CrossRef]
- MacInnis, C.I.; Keddie, B.A.; Pernal, S.F. Honey bees with a drinking problem: Potential routes of Nosema ceranae spore transmission. Parasitology 2022, 149, 573–580. [Google Scholar] [CrossRef]
- Janashia, I.; Westover, L.; Japoshvili, G. A review of Apis mellifera caucasica (Hym., Apidae): History, taxonomy and distribution. J. Insect Biodivers. Syst. 2025, 11, 455–468. [Google Scholar] [CrossRef]
- Haberl, M.; Tautz, D. Tri- and tetranucleotide microsatellite loci in honey bees (Apis mellifera)—A step towards quantitative genotyping. Mol. Ecol. 1999, 8, 1358–1360. [Google Scholar] [CrossRef]
- Solignac, M.; Vautrin, D.; Loiseau, A.; Mougel, F.; Baudry, E.; Estoup, A.; Garnery, L.; Haberl, M.; Cornuet, J.-M. Five hundred and fifty microsatellite markers for the study of the honeybee (Apis mellifera L.) genome. Mol. Ecol. 2003, 3, 307–311. [Google Scholar] [CrossRef]
- Garnery, L.; Solignac, M.; Celebrano, G.; Cornuet, J.-M. A simple test using restricted PCR-amplified mitochondrial DNA to study the genetic structure of Apis mellifera L. Experientia 1993, 49, 1016–1021. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Li, Y.L.; Liu, J.X. StructureSelector: A web-based software to select and visualize the optimal number of clusters using multiple methods. Mol. Ecol. Resour. 2018, 18, 176–177. [Google Scholar] [CrossRef]
- Fries, I.; Chauzat, M.-P.; Chen, Y.-P.; Doublet, V.; Genersch, E.; Gisder, S.; Higes, M.; McMahon, D.P.; Martín-Hernández, R.; Natsopoulou, M.; et al. Standard methods for Nosema research. J. Apic. Res. 2013, 52, 1–28. [Google Scholar] [CrossRef]
- Martín-Hernández, R.; Meana, A.; Prieto, L.; Salvador, A.M.; Garrido-Bailon, E.; Higes, M. Outcome of colonization of Apis mellifera by Nosema ceranae. Appl. Environ. Microbiol. 2007, 73, 6331–6338. [Google Scholar] [CrossRef]
- Tokarev, Y.S.; Zinatullina, Z.Y.; Ignatieva, A.N.; Zhigileva, O.N.; Malysh, J.M.; Sokolova, Y.Y. Detection of two Microsporidia pathogens of the European honey bee Apis mellifera (Insecta: Apidae) in Western Siberia. Acta Parasit. 2018, 63, 728–732. [Google Scholar] [CrossRef]
- Ostroverkhova, N.V.; Konusova, O.L.; Kucher, A.N.; Kireeva, T.N.; Rosseykina, S.A. Prevalence of the Microsporidian Nosema spp. in Honey Bee Populations (Apis mellifera) in Some Ecological Regions of North Asia. Vet. Sci. 2020, 7, 111. [Google Scholar] [CrossRef]
- Tokarev, Y.S.; Ignatieva, A.N.; Zinatullina, Z.Y. Molecular detection of Nosema disease. Pchelovodstvo 2010, 5, 18–19. (In Russian) [Google Scholar]
- Zinatullina, Z.Y.; Dolnikova, T.Y.; Domatskaya, T.F.; Domatsky, A.N. Monitoring diseases of honey bees (Apis mellifera) in Russia. Ukr. J. Ecol. 2018, 8, 106–112. [Google Scholar]
- Rortais, A.; Arnold, G.; Alburaki, M.; Legout, H.; Garnery, L. Review of the DraI COI-COII test for the conservation of the black honeybee (Apis mellifera mellifera). Conserv. Genet. Resour. 2011, 3, 383–391. [Google Scholar] [CrossRef]
- Hassett, J.; Browne, K.A.; McCormack, G.P.; Moore, E.; Society, N.I.H.B.; Soland, G.; Geary, M. A significant pure population of the dark European honey bee (Apis mellifera mellifera) remains in Ireland. J. Apic. Res. 2018, 57, 337–350. [Google Scholar] [CrossRef]
- Oleksa, A.; Kusza, S.; Tofilski, A. Mitochondrial DNA Suggests the Introduction of Honeybees of African Ancestry to East-Central Europe. Insects 2021, 12, 410. [Google Scholar] [CrossRef]
- Chávez-Galarza, J.; López-Montañez, R.; Jiménez, A.; Ferro-Mauricio, R.; Oré, J.; Medina, S.; Rea, R.; Vásquez, H. Mitochondrial DNA Variation in Peruvian Honey Bee (Apis mellifera L.) Populations Using the tRNAleu-cox2 Intergenic Region. Insects 2021, 12, 641. [Google Scholar] [CrossRef]
- Techer, M.A.; Clémencet, J.; Turpin, P.; Volbert, N.; Reynaud, B.; Delatte, H. Genetic characterization of the honeybee (Apis mellifera) population of Rodrigues Island, based on microsatellite and mitochondrial DNA. Apidologie 2015, 46, 445–454. [Google Scholar] [CrossRef]
- Chapman, N.C.; Bourgeois, A.L.; Beaman, L.D.; Lim, J.; Harpur, B.A.; Zayed, A.; Allsopp, M.H.; Rinderer, T.E.; Oldroyd, B.P. An abbreviated SNP panel for ancestry assignment of honeybees (Apis mellifera). Apidologie 2017, 48, 776–783. [Google Scholar] [CrossRef]
- Nikonorov, I.M.; Ben’kovskaya, G.V.; Poskryakov, A.V.; Nikolenko, A.G.; Vakhitov, V.A. The use of the PCR technique for control of pure breeding of honeybee (Apis mellifera mellifera L.) colonies from the Southern Urals. Russ. J. Genet. 1998, 34, 13441347. [Google Scholar]
- Brandorf, A.Z.; Ivoylova, M.M.; Ilyasov, R.A.; Poskryakov, A.V.; Nikolenko, A.G. Population-genetic differentiation of honey bees of the Kirov region. Bee J. 2012, 7, 14–16. (In Russian) [Google Scholar]
- Jensen, A.B.; Palmer, K.A.; Boomsma, J.J.; Pedersen, B.V. Varying degrees of Apis mellifera ligustica introgression in protected populations of the black honeybee, Apis mellifera mellifera, in northwest Europe. Mol. Ecol. 2005, 14, 93–106. [Google Scholar] [CrossRef]
- Soland-Reckeweg, G.; Heckel, G.; Neumann, P.; Fluri, P.; Excoffier, L. Gene flow in admixed populations and implications for the conservation of the Western honeybee, Apis mellifera. J. Insect Conserv. 2009, 13, 317–328. [Google Scholar] [CrossRef]
- Brown, M.J.F. Microsporidia: An Emerging Threat to Bumblebees? Trends Parasitol. 2017, 33, 754–762. [Google Scholar] [CrossRef]
- Hristov, P.; Shumkova, R.; Palova, N.; Neov, B. Factors Associated with Honey Bee Colony Losses: A Mini-Review. Vet. Sci. 2020, 7, 166. [Google Scholar] [CrossRef]
- Marín-García, P.J.; Peyre, Y.; Ahuir-Baraja, A.E.; Garijo, M.M.; Llobat, L. The Role of Nosema ceranae (Microsporidia: Nosematidae) in Honey Bee Colony Losses and Current Insights on Treatment. Vet. Sci. 2022, 9, 130. [Google Scholar] [CrossRef]
- Heo, J.; Yoo, D.S.; Cheon, D.S.; Kim, Y.; Kim, D.Y. Prevalence of pathogens in abnormal honey bees in South Korea, 2020–2023. J. Vet. Diagn. Investig. 2025, 37, 104–113. [Google Scholar] [CrossRef]
- Schüler, V.; Liu, Y.C.; Gisder, S.; Horchler, L.; Groth, D.; Genersch, E. Significant, but not biologically relevant: Nosema ceranae infections and winter losses of honey bee colonies. Commun. Biol. 2023, 6, 229. [Google Scholar] [CrossRef]
- Petric, A.; Guzman-Novoa, E.; Eberl, H.J. A mathematical model for the interplay of Nosema infection and forager losses in honey bee colonies. J. Biol. Dyn. 2016, 11 (Suppl. 2), 348–378. [Google Scholar] [CrossRef]
- D’Alvise, P.; Seeburger, V.; Gihring, K.; Kieboom, M.; Hasselmann, M. Seasonal dynamics and co-occurrence patterns of honey bee pathogens revealed by high-throughput RT-qPCR analysis. Ecol. Evol. 2019, 9, 10241–10252. [Google Scholar] [CrossRef]
- Jabal-Uriel, C.; Barrios, L.; Bonjour-Dalmon, A.; Caspi-Yona, S.; Chejanovsly, N.; Erez, T.; Henriques, D.; Higes, M.; Le Conte, Y.; Lopes, A.R.; et al. Epidemiology of the Microsporidium Nosema ceranae in Four Mediterranean Countries. Insects 2022, 13, 844. [Google Scholar] [CrossRef]
- Sgroi, G.; D’Auria, L.J.; Lucibelli, M.G.; Mancusi, A.; Proroga, Y.T.R.; Esposito, M.; Rea, S.; Signorelli, D.; Gargano, F.; D’Alessio, N.; et al. Bees on the run: Nosema spp. (Microsporidia) in Apis mellifera and related products, Italy. Front. Vet. Sci. 2025, 11, 1530169. [Google Scholar] [CrossRef]
- Baigazanov, A.; Tikhomirova, Y.; Valitova, N.; Nurkenova, M.; Koigeldinova, A.; Abdullina, E.; Zaikovskaya, O.; Ikimbayeva, N.; Zainettinova, D.; Bauzhanova, L. Occurrence of Nosemosis in honey bee, Apis mellifera L. at the apiaries of East Kazakhstan. PeerJ 2022, 10, e14430. [Google Scholar] [CrossRef]
- Lannutti, L.; Mira, A.; Basualdo, M.; Rodriguez, G.; Erler, S.; Silva, V.; Gisder, S.; Genersch, E.; Florin-Christensen, M.; Schnittger, L. Development of a Loop-Mediated Isothermal Amplification (LAMP) and a Direct LAMP for the Specific Detection of Nosema ceranae, a Parasite of Honey Bees. Parasitol. Res. 2020, 119, 3947–3956. [Google Scholar] [CrossRef] [PubMed]
- Mutinelli, F. The spread of pathogens through trade in honey bees and their products (including queen bees and semen): Overview and recent developments. Rev. Sci. Tech. 2011, 30, 257–271. [Google Scholar] [CrossRef] [PubMed]
- Owen, R. Role of Human Action in the Spread of Honey Bee (Hymenoptera: Apidae) Pathogens. J. Econ. Entomol. 2017, 110, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Idrissou, F.O.; Huang, Q.; Yañez, O.; Neumann, P. International beeswax trade facilitates small hive beetle invasions. Sci. Rep. 2019, 9, 10665. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Number of Colonies | ||||||
|---|---|---|---|---|---|---|
| Group | Number of Colonies | N. apis | N. ceranae | Nosemosis | Coinfection | Microscopy |
| c | 151 | 43 | 52 | 81 | 14 | 49 |
| h | 58 | 14 | 17 | 23 | 8 | 16 |
| m | 140 | 30 | 33 | 49 | 14 | 33 |
| Total | 349 | 87 | 102 | 153 | 36 | 98 |
| Region | M mtDNA | C mtDNA | M ≥ 0.88 nDNA | N. apis | N. ceranae | Coinfection |
|---|---|---|---|---|---|---|
| Altai Territory (N = 8) | 8 | 0 | 4 | 0 | 0 | 0 |
| Belgorod Region (N = 29) | 6 | 23 | 6 | 7 | 14 | 2 |
| Krasnodar Territory (N = 29) | 0 | 29 | 0 | 11 | 9 | 4 |
| Leningrad Region (N = 10) | 10 | 0 | 0 | 1 | 3 | 0 |
| Novgorod Region (N = 17) | 0 | 17 | 0 | 3 | 9 | 2 |
| Orenburg Region (N = 5) | 0 | 5 | 0 | 3 | 1 | 0 |
| Republic of Adygeya (N = 26) | 0 | 26 | 0 | 19 | 7 | 6 |
| Ryazan Region (N = 10) | 0 | 10 | 0 | 1 | 3 | 0 |
| Samara Region (N = 3) | 0 | 3 | 0 | 0 | 2 | 0 |
| Sverdlovsk Region (N = 12) | 10 | 2 | 7 | 1 | 3 | 0 |
| Ulyanovsk Region (N = 1) | 0 | 1 | 0 | 1 | 1 | 1 |
| Republic of Bashkortostan (N = 199) | 164 | 35 | 123 | 40 | 50 | 21 |
| Summary (N = 349) | 198 | 151 | 140 | 87 | 102 | 36 |
| Group | N. apis (%) | N. ceranae (%) | Nosemosis (%) |
|---|---|---|---|
| c = 151 | 43 (28%) | 52 (34%) | 81 (54%) |
| h = 58 | 14 (24%) | 17 (29%) | 23 (40%) |
| m = 140 | 30 (21%) | 33 (24%) | 49 (35%) |
| Pearson’s chi-squared test | X-squared = 1.9517, df = 2, p-value = 0.3769 | X-squared = 4.1467, df = 2, p-value = 0.1258 | X-squared = 10.749, df = 2, p-value = 0.004633 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaskinova, M.; Gaifullina, L.; Zaitsev, G.; Davydychev, A.; Saltykova, E. Nosemosis in Russian Apis mellifera L. Populations: Distribution and Association with Hybridization. Insects 2025, 16, 641. https://doi.org/10.3390/insects16060641
Kaskinova M, Gaifullina L, Zaitsev G, Davydychev A, Saltykova E. Nosemosis in Russian Apis mellifera L. Populations: Distribution and Association with Hybridization. Insects. 2025; 16(6):641. https://doi.org/10.3390/insects16060641
Chicago/Turabian StyleKaskinova, Milyausha, Luisa Gaifullina, Gleb Zaitsev, Alexandr Davydychev, and Elena Saltykova. 2025. "Nosemosis in Russian Apis mellifera L. Populations: Distribution and Association with Hybridization" Insects 16, no. 6: 641. https://doi.org/10.3390/insects16060641
APA StyleKaskinova, M., Gaifullina, L., Zaitsev, G., Davydychev, A., & Saltykova, E. (2025). Nosemosis in Russian Apis mellifera L. Populations: Distribution and Association with Hybridization. Insects, 16(6), 641. https://doi.org/10.3390/insects16060641