
Exploration of the Effects of Different Beauveria bassiana Strains on Dioryctria sylvestrella Larvae from the Perspective of Oxidative Stress

 , , and
, , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
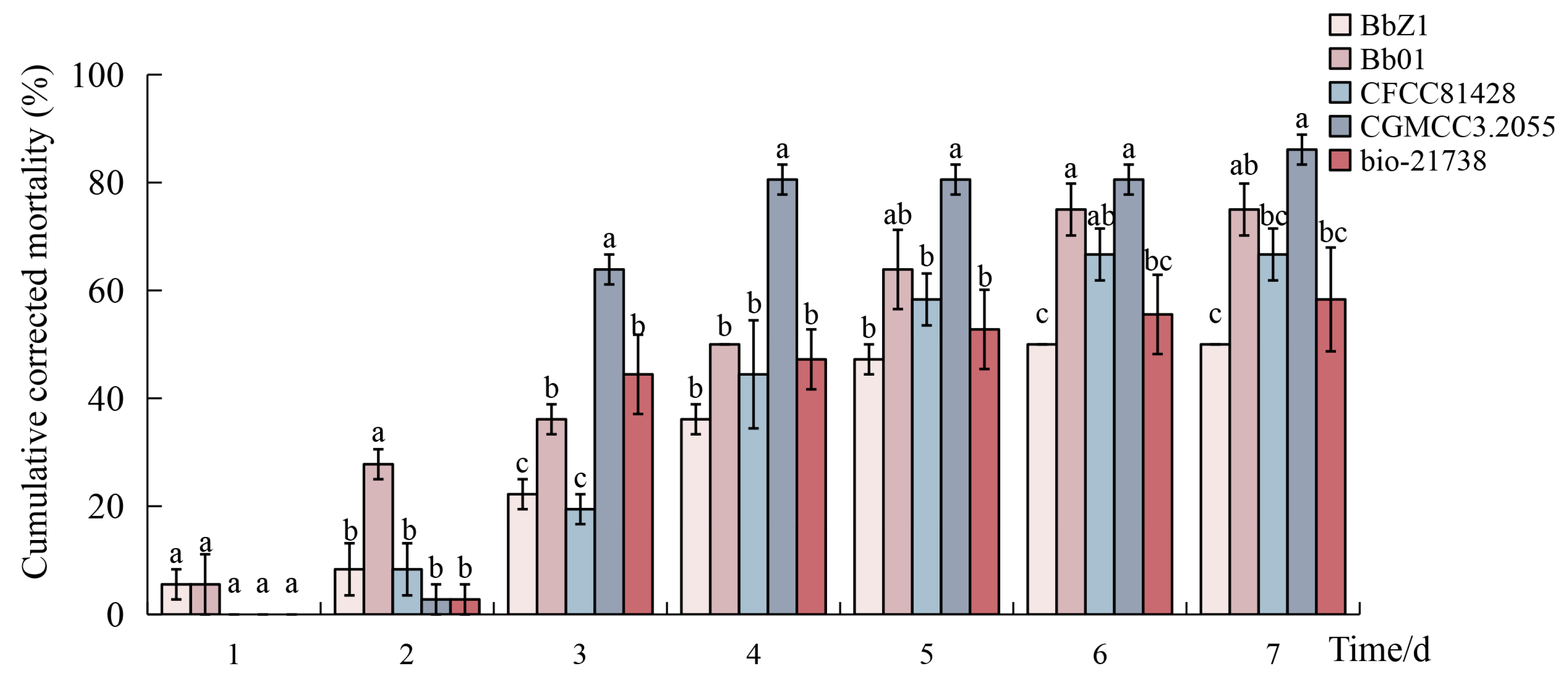
3.1. Screening of Highly Pathogenic Strains
3.2. The Defense Response System of D. sylvestrella Larvae Against B. bassiana
3.2.1. The Influence of B. bassiana on the Antioxidant Capacity of D. sylvestrella Larvae
3.2.2. Influence of B. bassiana on Redox System of D. sylvestrella Larvae
3.2.3. Effects of B. bassiana on the Detoxification Enzymes of D. sylvestrella Larvae
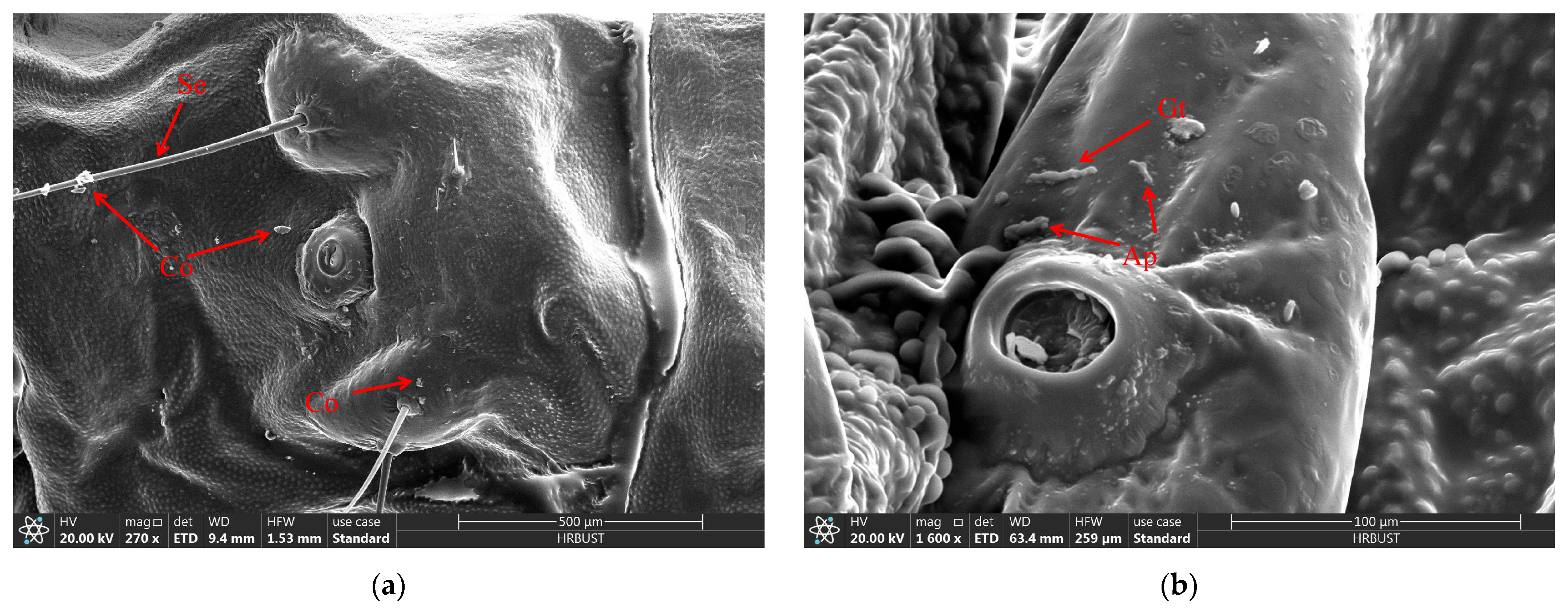
3.3. Microscopic Structure Observation of D. sylvestrella Larvae Infected by B. bassiana
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Y.X.; Zhang, X.X.; Cai, K.W.; Zhang, Q.H.; Jiang, L.P.; Li, H.X.; Lv, Y.Z.; Qu, G.Z.; Zhao, X.Y. Comparative transcriptomic and metabolic analyses reveal the coordinated mechanisms in Pinus koraiensis under different light stress conditions. Int. J. Mol. Sci. 2022, 23, 9556. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Yan, S.C.; Nie, W.L.; Wang, Y.L. A complementary study on biological characteristics of two Species of Dioryctria (Lepidoptera: Pyralidae) and their damage to cones and twigs of Korean Pine. Sci. Silv. Sin. 2010, 46, 188–192. [Google Scholar]
- Jactel, H.; Goulard, M.; Menassieu, P.; Goujon, G. Habitat diversity in forest plantations reduces infestations of the pine stem borer Dioryctria sylvestrella. J. Appl. Ecol. 2002, 39, 618–628. [Google Scholar] [CrossRef]
- Lv, C.H.; Liu, C.; Zhang, M.L.; Bai, H.Y.; Li, X.X.; Wang, L.G. Effect contrast test of spraying three kinds of pollution-free pesticides against Dioryctria sylvestrella by drone. For. Pest Dis. 2021, 40, 40–43. [Google Scholar]
- Jin, M.H.; Peng, Y.; Peng, J.; Zhang, H.H.; Shan, Y.X.; Liu, K.Y.; Xiao, Y.T. Transcriptional regulation and overexpression of GST cluster enhances pesticide resistance in the cotton bollworm, Helicoverpa armigera (Lepidoptera: Noctuidae). Commun. Biol. 2023, 6, 1064. [Google Scholar] [CrossRef]
- Meng, S.K.; Liang, C.P.; Zheng, Q.; Zhu, S.Q.; Wu, J.; Wang, B.T.; Wang, Y.Q.; Zhang, Z.X.; Cheng, D.M. Insecticidal effect and mechanism of Metarhizium anisopliae ZHKUJGZ1 against Solenopsis invicta (Hymenoptera: Formicidae). Pestic. Biochem. Physiol. 2025, 209, 106350. [Google Scholar] [CrossRef]
- Valero-Jiménez, C.A.; Wiegers, H.; Zwaan, B.J.; Koenraadt, C.J.; van Kan, J.A. Genes involved in virulence of the entomopathogenic fungus Beauveria bassiana. J. Invertebr. Pathol. 2016, 133, 41–49. [Google Scholar] [CrossRef]
- Liu, J.; Ling, Z.Q.; Wang, J.J.; Xiang, T.T.; Xu, L.; Gu, C.X.; Liu, R.; Xu, J.; Xu, C.L.; Zhou, W.; et al. In vitro transcriptomes analysis identifies some special genes involved in pathogenicity difference of the Beauveria bassiana against different insect hosts. Microb. Pathog. 2021, 154, 104824. [Google Scholar] [CrossRef]
- Wang, J.J.; Bai, W.W.; Zhou, W.; Liu, J.; Chen, J.; Liu, X.Y.; Xiang, T.T.; Liu, R.H.; Wang, W.H.; Zhang, B.L.; et al. Transcriptomic analysis of two Beauveria bassiana strains grown on cuticle extracts of the silkworm uncovers their different metabolic response at early infection stage. J. Invertebr. Pathol. 2017, 145, 45–54. [Google Scholar] [CrossRef]
- Kim, S.; Kim, J.C.; Lee, S.J.; Lee, M.R.; Park, S.E.; Li, D.; Baek, S.; Shin, T.Y.; Kim, J.S. Beauveria bassiana ERL836 and JEF-007 with similar virulence show different gene expression when interacting with cuticles of western flower thrips, Frankniella occidentalis. BMC Genom. 2020, 21, 836. [Google Scholar] [CrossRef]
- Zhang, C.; Teng, B.; Liu, H.M.; Wu, C.Y.; Wang, L.; Jin, S. Impact of Beauveria bassiana on antioxidant enzyme activities and metabolomic profiles of Spodoptera frugiperda. J. Invertebr. Pathol. 2023, 198, 107929. [Google Scholar] [CrossRef] [PubMed]
- Lalouette, L.; Williams, C.M.; Hervant, F.; Sinclair, B.J.; Renault, D. Metabolic rate and oxidative stress in insects exposed to low temperature thermal fluctuations. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2011, 158, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, V.I. Free radicals, reactive oxygen species, oxidative stress and its classification. Chem. Biol. Interact. 2014, 224, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Kinareikina, A.; Silivanova, E. Impact of insecticides at sublethal concentrations on the enzyme activities in adult Musca domestica L. Toxics 2023, 11, 47. [Google Scholar] [CrossRef]
- Liu, Z.C.; Zhou, L.; Wang, J.L.; Liu, X.S. Expression of a phenoloxidase cascade inhibitor enhances the virulence of the fungus Beauveria bassiana against the insect Helicoverpa armigera. Dev. Comp. Immunol. 2021, 117, 103986. [Google Scholar] [CrossRef]
- Wang, X.M. Study on the Pathogenicity of Biocontrol Strains in Dioryctria abietella. Ph.D. Thesis, Northeast Forestry University, Harbin, China, 2023. [Google Scholar]
- Liu, X.; Nie, Z.Y.; Chi, D.F.; He, L.M. Toxicity and killing effect of Beauveria bassiana and Pyemotes zhonghuajia on Dioryctria abietella and related enzyme activities. J. Northeast. For. Univ. 2023, 51, 139–146. [Google Scholar]
- Zhang, H.W.; Chen, R.T.; Xu, J.J.; Zhou, J.H.; Shi, P.; Yang, Q.S.; Han, J.; Chu, K.; Yu, J.; Chi, D.F. Scanning electron microscopy and pathogenicity of Beauveria bassiana infection with Hylurgus ligniperda. J. Cent. South. Univ. For. Technol. 2024, 44, 62–72. [Google Scholar]
- Qu, S.; Wang, S.B. Interaction of entomopathogenic fungi with the host immune system. Dev. Comp. Immunol. 2018, 83, 96–103. [Google Scholar] [CrossRef]
- Brahma, S.; Chatterjee, S.; Dey, A. Role of eicosanoids in insect immunity: New insights and recent advances. Insect Sci. 2024; advance online publication. [Google Scholar] [CrossRef]
- Forlani, L.; Juárez, M.P.; Lavarías, S.; Pedrini, N. Toxicological and biochemical response of the entomopathogenic fungus Beauveria bassiana after exposure to deltamethrin. Pest. Manag. Sci. 2014, 70, 751–756. [Google Scholar] [CrossRef]
- Feng, A.Q.; Zhao, Z.W.; Liu, C.F.; Du, C.; Gao, P.Y.; Liu, X.G.; Li, D.Q. Study on characterization of Bupleurum chinense polysaccharides with antioxidant mechanisms focus on ROS relative signaling pathways and anti-aging evaluation in vivo model. Int. J. Biol. Macromol. 2024, 266 Pt 2, 131171. [Google Scholar] [CrossRef]
- Gogoi, K.; Gogoi, H.; Borgohain, M.; Saikia, R.; Chikkaputtaiah, C.; Hiremath, S.; Basu, U. The molecular dynamics between reactive oxygen species (ROS), reactive nitrogen species (RNS) and phytohormones in plant’s response to biotic stress. Plant Cell Rep. 2024, 43, 263. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.T.; Wang, Q.; Tan, M.T.; Zhang, A.Y.; Yan, S.C.; Jiang, D. Toxicological assessment of cadmium exposure through Hyphantria cunea larvae on the predation fitness of Arma chinensis. Sci. Total Environ. 2024, 949, 175142. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.T.; Yuan, Y.D.; Cai, M.; Wang, D.S.; Lv, W.G. Toxicity of isoprocarb to earthworms (Eisenia fetida): Oxidative stress, neurotoxicity, biochemical responses and detoxification mechanisms. Environ. Pollut. 2021, 290, 118038. [Google Scholar] [CrossRef] [PubMed]
- Mao, G.L.; Lu, W.; Xu, Y.H.; Liu, H.; Xu, H.H.; Zeng, J.W. Bruceine D inhibits the growth of Spodoptera litura by inducing cell apoptosis in the midgut via an oxidative burst. Pest. Manag. Sci. 2024, 80, 3126–3139. [Google Scholar] [CrossRef]
- Bano, A.; Muqarab, R. Plant defence induced by PGPR against Spodoptera litura in tomato (Solanum lycopersicum L.). Plant Biol. 2017, 19, 406–412. [Google Scholar] [CrossRef]
- Winkiel, M.J.; Chowański, S.; Walkowiak-Nowicka, K.; Lubawy, J.; Słocińska, M. Modulation of the antioxidant system by glycoalkaloids in the beetle Tenebrio molitor L. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2024, 286, 110018. [Google Scholar] [CrossRef]
- Liu, X.; Bian, D.D.; Jiang, Q.; Jiang, J.J.; Jin, Y.; Chen, F.X.; Zhang, D.Z.; Liu, Q.N.; Tang, B.P.; Dai, L.S. Insights into chlorantraniliprole exposure via activating cytochrome P450-mediated xenobiotic metabolism pathway in the Procambarus clarkii: Identification of P450 genes involved in detoxification. Int. J. Biol. Macromol. 2024, 277 Pt 2, 134231. [Google Scholar] [CrossRef]
- Zhang, T.S.; Yuan, Y.D.; Teng, H.Y.; Wang, D.S.; Gu, H.T. Exposure to cyantraniliprole adversely impacts fitness of Harmonia axyridis: Acute toxicity and sublethal effects on development, fecundity and antioxidant responses. Insects 2024, 15, 773. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef]
- Liang, G.B.; Liao, X.Y.; Du, G.C.; Chen, J. A new strategy to enhance glutathione production by multiple H2O2 induced oxidative stresses in Candida utilis. Bioresour. Technol. 2009, 100, 350–355. [Google Scholar] [CrossRef]
- Yang, Y.L.; Tao, J.L.; Liang, F.F.; Sun, B.; Jiang, J.B.; Zhao, M.X.; Gao, D. A fluorescent probe for ultrarapid H2O2 detection during reagent-stimulated oxidative stress in cells and zebrafish. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2024, 313, 124061. [Google Scholar] [CrossRef] [PubMed]
- Karthi, S.; Vaideki, K.; Shivakumar, M.S.; Ponsankar, A.; Thanigaivel, A.; Chellappandian, M.; Vasantha-Srinivasan, P.; Muthu-Pandian, C.K.; Hunter, W.B.; Senthil-Nathan, S. Effect of Aspergillus flavus on the mortality and activity of antioxidant enzymes of Spodoptera litura Fab. (Lepidoptera: Noctuidae) larvae. Pestic. Biochem. Physiol. 2018, 149, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Chaurasia, A.; Lone, Y.; Wani, O.; Gupta, U.S. Effect of certain entomopathogenic fungi on oxidative stress and mortality of Periplaneta americana. Pestic. Biochem. Physiol. 2016, 127, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Yang, H.; Wang, Z.; Long, G.Y.; Jin, D.C. Protective and detoxifying enzyme activity and ABCG subfamily gene expression in Sogatella furcifera under insecticide stress. Front. Physiol. 2019, 9, 1890. [Google Scholar] [CrossRef]
- Seo, Y.J.; Lee, J.W.; Lee, E.H.; Lee, H.K.; Kim, H.W.; Kim, Y.H. Role of glutathione in the adaptive tolerance to H2O2. Free Radic. Biol. Med. 2004, 37, 1272–1281. [Google Scholar] [CrossRef]
- Chen, R.T.; Zhuang, Y.T.; Wang, M.L.; Yu, J.; Chi, D.F. Transcriptomic analysis of the response of the Dioryctria abietella larva midgut to Bacillus thuringiensis 2913 infection. Int. J. Mol. Sci. 2024, 25, 10921. [Google Scholar] [CrossRef]
- Bitencourt, R.O.B.; Salcedo-Porras, N.; Umaña-Diaz, C.; da Costa Angelo, I.; Lowenberger, C. Antifungal immune responses in mosquitoes (Diptera: Culicidae): A review. J. Invertebr. Pathol. 2021, 178, 107505. [Google Scholar] [CrossRef]
- Lu, A.R.; Zhang, Q.L.; Zhang, J.; Yang, B.; Wu, K.; Xie, W.; Luan, Y.X.; Ling, E.J. Insect prophenoloxidase: The view beyond immunity. Front. Physiol. 2014, 5, 252. [Google Scholar] [CrossRef]
- Ghanem, M.M.E.; Abd-Elaziz, A.M.; Mohamed, M.A. Biochemical and toxicological characteristics of polyphenol oxidase from red palm weevil Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2025, 287, 110044. [Google Scholar] [CrossRef]
- Xu, L.; Ma, L.; Wang, W.; Li, L.; Lu, Z. Phenoloxidases are required for the pea aphid’s defence against bacterial and fungal infection. Insect Mol. Biol. 2019, 28, 176–186. [Google Scholar] [CrossRef]
- Quesada-Moraga, E.; Maranhao, E.A.A.; Valverde-García, P.; Santiago-Álvarez, C. Selection of Beauveria bassiana isolates for control of the whiteflies Bemisia tabaci and Trialeurodes vaporariorum on the basis of their virulence, thermal requirements, and toxicogenic activity. Biol. Control. 2006, 36, 274–287. [Google Scholar] [CrossRef]
- Gao, Y.P.; Luo, M.; Wang, X.Y.; He, X.Z.; Lu, W.; Zheng, X.L. Pathogenicity of Beauveria bassiana PfBb and immune responses of a non-target host, Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2022, 13, 914. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.; Noh, M.Y.; Mun, S.; Lee, S.J.; Arakane, Y.; Kim, J.S. Ultrastructural analysis of beetle larva cuticles during infection with the entomopathogenic fungus, Beauveria bassiana. Pest. Manag. Sci. 2022, 78, 3356–3364. [Google Scholar] [CrossRef] [PubMed]
- Charnley, A.K.; St. Leger, R.J. The role of cuticle-degrading enzymes in fungal pathogenesis in insects. In The Fungal Spore and Disease Initiation in Plants and Animals; Cole, G.T., Hoch, H.C., Eds.; Springer: Boston, MA, USA, 1991; pp. 267–286. [Google Scholar]
- Xiong, Q.; Xie, Y.P.; Zhu, Y.M.; Xue, J.L.; Li, J.; Fan, R.J. Morphological and ultrastructural characterization of Carposina sasakii larvae (Lepidoptera: Carposinidae) infected by Beauveria bassiana (Ascomycota: Hypocreales: Clavicipitaceae). Micron 2013, 44, 303–311. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain No. | Strains | Original Hosts |
|---|---|---|
| CGMCC3.2055 | B. bassiana | Dendrolimus spp. |
| CFCC81428 | B. bassiana | Monochamus alternatus |
| bio-21738 | B. bassiana | Dendrolimus punctatus |
| Bb01 | B. bassiana | Xylotrechus rusticus |
| BbZ1 | B. bassiana | Hylurgus ligniperda |
| Strains | Regression Equation | Goodness of Fit Test χ2 | p Value | LT50 (d) | 95% Confidence Interval |
|---|---|---|---|---|---|
| BbZ1 | Y = −1.788 + 0.980x | 5.914 | 0.998 | 6.200 | 5.116–8.410 |
| Bb01 | Y = −1.532 + 1.162x | 8.878 | 0.975 | 3.739 | 3.214–4.347 |
| CGMCC3.2055 | Y = −2.265 + 1.922x | 22.453 | 0.262 | 3.248 | 2.884–3.593 |
| Bio-21738 | Y = −2.045 + 1.285x | 22.222 | 0.273 | 4.911 | 4.290–5.792 |
| CFCC81428 | Y = −2.522 + 1.621x | 10.020 | 0.952 | 4.737 | 4.237–5.364 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, R.; Wang, M.; Zhang, H.; Xu, J.; Wang, X.; Chi, D.; Yu, J. Exploration of the Effects of Different Beauveria bassiana Strains on Dioryctria sylvestrella Larvae from the Perspective of Oxidative Stress. Insects 2025, 16, 640. https://doi.org/10.3390/insects16060640
Chen R, Wang M, Zhang H, Xu J, Wang X, Chi D, Yu J. Exploration of the Effects of Different Beauveria bassiana Strains on Dioryctria sylvestrella Larvae from the Perspective of Oxidative Stress. Insects. 2025; 16(6):640. https://doi.org/10.3390/insects16060640
Chicago/Turabian StyleChen, Ruting, Meiling Wang, Hanwen Zhang, Jianjiao Xu, Xiaomei Wang, Defu Chi, and Jia Yu. 2025. "Exploration of the Effects of Different Beauveria bassiana Strains on Dioryctria sylvestrella Larvae from the Perspective of Oxidative Stress" Insects 16, no. 6: 640. https://doi.org/10.3390/insects16060640
APA StyleChen, R., Wang, M., Zhang, H., Xu, J., Wang, X., Chi, D., & Yu, J. (2025). Exploration of the Effects of Different Beauveria bassiana Strains on Dioryctria sylvestrella Larvae from the Perspective of Oxidative Stress. Insects, 16(6), 640. https://doi.org/10.3390/insects16060640