
Glycosidase Isoforms in Honey and the Honey Bee (Apis mellifera L.): Differentiating Bee- and Yeast-Derived Enzymes and Implications for Honey Authentication

 ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Honey Bee Collection, Dissection, and Sample Preparation
2.3. Isoelectric Focusing and Zymogram Detection of Glycosidase Activity
2.4. Determination of α-Glucosidase pI Values
2.5. Analysis of Sugar Hydrolysis Products
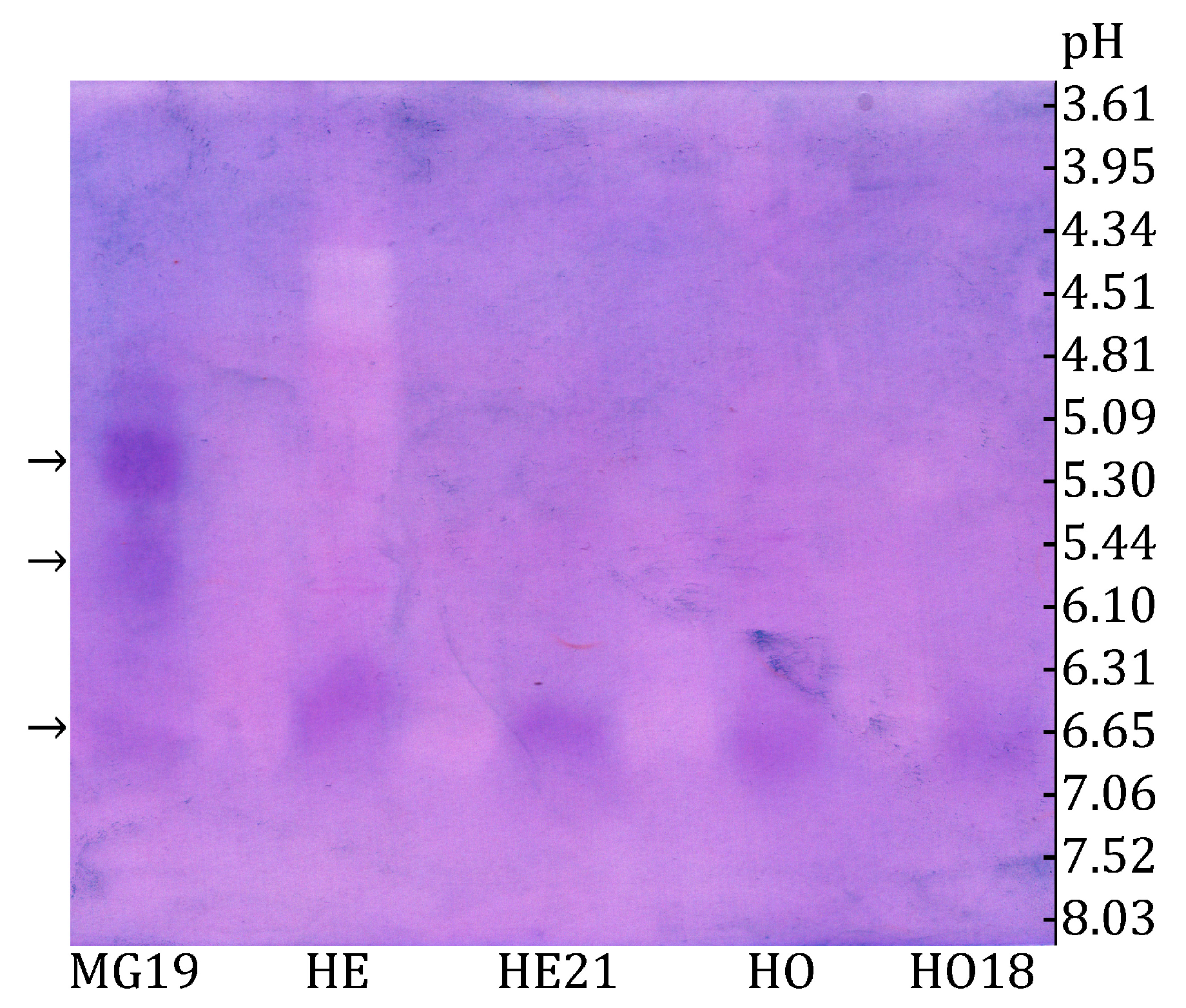
3. Results
3.1. Characterization of Honey Bee α-Glucosidases
3.2. Distinguishing Between α-Glucosidase and β-Fructofuranosidase
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vanengelsdorp, D.; Hayes, J.; Underwood, R.M.; Caron, D.; Pettis, J. A Survey of Managed Honey Bee Colony Losses in the USA, Fall 2009 to Winter 2010. J. Apic. Res. 2011, 50, 1–10. [Google Scholar] [CrossRef]
- Bryś, M.S.; Skowronek, P.; Strachecka, A. Pollen Diet—Properties and Impact on a Bee Colony. Insects 2021, 12, 798. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, E.M.; Dolezal, A.G. Nutrition, Pesticide Exposure, and Virus Infection Interact to Produce Context-Dependent Effects in Honey Bees (Apis mellifera). Sci. Total Environ. 2024, 949, 175125. [Google Scholar] [CrossRef] [PubMed]
- Pavlović, R.; Brodschneider, R.; Goessler, W.; Stanisavljević, L.; Vujčić, Z.; Zarić, N.M. Micronutrient Deficiency May Be Associated with the Onset of Chalkbrood Disease in Honey Bees. Insects 2024, 15, 269. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Salignon, M.; Le Conte, Y.; Belzunces, L.P.; Decourtye, A.; Kretzschmar, A.; Suchail, S.; Brunet, J.L.; Alaux, C. Influence of Pollen Nutrition on Honey Bee Health: Do Pollen Quality and Diversity Matter? PLoS ONE 2013, 8, e72016. [Google Scholar] [CrossRef]
- Brodschneider, R.; Crailsheim, K. Nutrition and Health in Honey Bees. Apidologie 2010, 41, 278–294. [Google Scholar] [CrossRef]
- Scofield, H.N.; Mattila, H.R. Honey Bee Workers That Are Pollen Stressed as Larvae Become Poor Foragers and Waggle Dancers as Adults. PLoS ONE 2015, 10, e0121731. [Google Scholar] [CrossRef]
- Taylor, M.A.; Robertson, A.W.; Biggs, P.J.; Richards, K.K.; Jones, D.F.; Parkar, S.G. The Effect of Carbohydrate Sources: Sucrose, Invert Sugar and Components of Mānuka Honey, on Core Bacteria in the Digestive Tract of Adult Honey Bees (Apis mellifera). PLoS ONE 2019, 14, e0225845. [Google Scholar] [CrossRef]
- Wheeler, M.M.; Robinson, G.E. Diet-Dependent Gene Expression in Honey Bees: Honey vs. Sucrose or High Fructose Corn Syrup. Sci. Rep. 2014, 4, srep05726. [Google Scholar] [CrossRef]
- Přidal, A.; Musila, J.; Svoboda, J. Condition and Honey Productivity of Honeybee Colonies Depending on Type of Supplemental Feed for Overwintering. Animals 2023, 13, 323. [Google Scholar] [CrossRef]
- Standifer, L.N.; Moeller, F.E.; Kauffeld, N.M.; Herbert, E.W., Jr.; Shimanuki, H. Supplemental Feeding of Honey Bee Colonies. Agric. Inf. Bull. 1978, 413, 1–11. [Google Scholar] [CrossRef]
- Camazine, S.; Sneyd, J.; Jenkins, M.J.; Murray, J.D. A Mathematical Model of Self-Organized Pattern Formation on the Combs of Honeybee Colonies. J. Theor. Biol. 1990, 147, 553–571. [Google Scholar] [CrossRef]
- Pavlović, R.; Dojnov, B.; Slavić, M.Š.; Ristović, M.; Vujčić, M.; Stojanović, S.; Vujčić, Z. Differential Processing of Sucrose and Invert Syrup in Honey Bees. Arch. Insect Biochem. Physiol. 2025, 118, e70052. [Google Scholar] [CrossRef]
- Kotwal, S.M.; Shankar, V. Immobilized Invertase. Biotechnol. Adv. 2009, 27, 311–322. [Google Scholar] [CrossRef]
- Margetić, A.; Vujčić, Z. Comparative Study of Stability of Soluble and Cell Wall Invertase from Saccharomyces cerevisiae. Prep. Biochem. Biotechnol. 2017, 47, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Özcan, M.; Arslan, D.; Ali Ceylan, D. Effect of Inverted Saccharose on Some Properties of Honey. Food Chem. 2006, 99, 24–29. [Google Scholar] [CrossRef]
- Basser, A.; Shall, S. Properties of the Internal Invertase of Yeast, Saccharomyces cerevisiae. Biochim. Biophys. Acta (BBA)—Enzymol. 1971, 250, 192–202. [Google Scholar] [CrossRef]
- Kaewmuangmoon, J.; Kilaso, M.; Leartsakulpanich, U.; Kimura, K.; Kimura, A.; Chanchao, C. Expression of a Secretory α-Glucosidase II from Apis Cerana Indica in Pichia Pastoris and Its Characterization. BMC Biotechnol. 2013, 13, 16. [Google Scholar] [CrossRef]
- White, J.W.; Maher, J. Transglucosidation by Honey Invertasel. Arch. Biochem. Biophys. 1952, 42, 360–367. [Google Scholar] [CrossRef]
- Bhalla, T.C.; Thakur, N.; Thakur, N. Invertase of Saccharomyces cerevisiae SAA-612: Production, Characterization and Application in Synthesis of Fructo-Oligosaccharides. LWT 2017, 77, 178–185. [Google Scholar] [CrossRef]
- Kimura, A.; Yoshida-Kitahara, F.; Chiba, S. Characteristics of Transglucosylation of Honeybee α-Glucosidase I. Agric. Biol. Chem. 1987, 51, 1859–1864. [Google Scholar] [CrossRef]
- Silva, S.P.; Moreira, A.S.P.; Domingues, M.D.R.M.; Evtuguin, D.V.; Coelho, E.; Coimbra, M.A. Contribution of Non-Enzymatic Transglycosylation Reactions to the Honey Oligosaccharides Origin and Diversity. Pure Appl. Chem. 2019, 91, 1231–1242. [Google Scholar] [CrossRef]
- LeBlanc, B.W.; Eggleston, G.; Sammataro, D.; Cornett, C.; Dufault, R.; Deeby, T.; Cyr, E.S. Formation of Hydroxymethylfurfural in Domestic High-Fructose Corn Syrup and Its Toxicity to the Honey Bee (Apis mellifera). J. Agric. Food Chem. 2009, 57, 7369–7376. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, R.; Hebbar, H.U.; Rastogi, N.K. Processing of Honey: A Review. Int. J. Food Prop. 2007, 10, 127–143. [Google Scholar] [CrossRef]
- Nishimoto, M.; Kubota, M.; Tsuji, M.; Mori, H.; Kimura, A.; Matsui, H.; Chiba, S. Purification and Substrate Specificity of Honeybee, Apis mellifera L., α-Glucosidase III. Biosci. Biotechnol. Biochem. 2001, 65, 1610–1616. [Google Scholar] [CrossRef]
- Nicolson, S.W. Sweet Solutions: Nectar Chemistry and Quality. Philos. Trans. R. Soc. B Biol. Sci. 2022, 377, 20210163. [Google Scholar] [CrossRef]
- Nicolson, S.W.; Thornburg, R.W. Nectar Chemistry. In Nectaries and Nectar; Nicolson, S.W., Nepi, M., Pacini, E., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 215–264. ISBN 978-1-4020-5936-0. [Google Scholar]
- Nation, J.L. Insect Physiology and Biochemistry, 2nd ed.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2008; ISBN 978-1-4200-6177-2. [Google Scholar]
- Machado De-Melo, A.A.; de Almeida-Muradian, L.B.; Sancho, M.T.; Pascual-Maté, A. Composition and Properties of Apis mellifera Honey: A Review. J. Apic. Res. 2018, 57, 5–37. [Google Scholar] [CrossRef]
- Kubo, T.; Sasaki, M.; Nakamura, J.; Sasagawa, H.; Ohashi, K.; Takeuchi, H.; Natori, S. Change in the Expression of Hypopharyngeal-Gland Proteins of the Worker Honeybees (Apis mellifera L.) with Age and/or Role. J. Biochem. 1996, 119, 291–295. [Google Scholar] [CrossRef]
- Takewaki, S.; Chiba, S.; Kimura, A.; Matsui, H.; Koike, Y. Purification and Properties of A-Glucosidases of the Honey Bee Apis mellifera L. Agric. Biol. Chem. 1980, 44, 731–740. [Google Scholar] [CrossRef]
- Wongchawalit, J.; Yamamoto, T.; Nakai, H.; Kim, Y.M.; Sato, N.; Nishimoto, M.; Okuyama, M.; Mori, H.; Saji, O.; Chanchao, C.; et al. Purification and Characterization of α-Glucosidase I from Japanese Honeybee (Apis cerana japonica) and Molecular Cloning of Its CDNA. Biosci. Biotechnol. Biochem. 2006, 70, 2889–2898. [Google Scholar] [CrossRef]
- Tomczyk, M.; Czerniecka-Kubicka, A.; Miłek, M.; Sidor, E.; Dżugan, M. Tracking of Thermal, Physicochemical, and Biological Parameters of a Long-Term Stored Honey Artificially Adulterated with Sugar Syrups. Molecules 2023, 28, 1736. [Google Scholar] [CrossRef] [PubMed]
- Giebel, W.; Zwilling, R.; Pfleiderer, G. The Evolution of Endopeptidases-XII. The Proteolytic Enzymes of the Honeybee (Apis mellifera L.): Purification and Characterization of Endopeptidases in the Midgut of Adult Workers and Comparative Studies on the Endopeptidase-Pattern of Different Castes and on Different Ontogenetic Stages. Comp. Biochem. Physiol. Part. B Comp. Biochem. 1971, 38, 197–206. [Google Scholar] [CrossRef]
- Crailsheim, K.; Schneider, L.H.W.; Hrassnigg, N.; Buhlmann, G.; Brosch, U.; Gmeinbauer, R.; Schoffmann, B. Pollen Consumption and Utilization in Worker Honeybees (Apis mellifera carnica): Dependence on Individual Age and Function. J. Insect Physiol. 1992, 38, 409–419. [Google Scholar] [CrossRef]
- Pavlović, R.; Šokarda Slavić, M.; Margetić, A.; Božić, N.; Vujčić, M.; Dojnov, B.; Vujčić, Z. Exploring the Connection between Food and Midgut Digestive Enzymes to Improve Honey Bee (Apis mellifera) Nutrition. J. Apic. Res. 2024. [Google Scholar] [CrossRef]
- Bernfeld, P. Amylases, α and β. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1955; Volume 1, pp. 149–158. [Google Scholar] [CrossRef]
- Davis, B.J. Disc Electrophoresis. II. Method and Application to Human Serum Proteins. Ann. N. Y. Acad. Sci. 1964, 121, 404–427. [Google Scholar] [CrossRef] [PubMed]
- Jue, C.K.; Lipke, P.N. Determination of Reducing Sugars in the Nanomole Range with Tetrazolium Blue. J. Biochem. Biophys. Methods 1985, 11, 109–115. [Google Scholar] [CrossRef]
- Kubota, M.; Tsuji, M.; Nishimoto, M.; Wongchawalit, J.; Okuyama, M.; Mori, H.; Matsui, H.; Surarit, R.; Svasti, J.; Kimura, A.; et al. Localization of α-Glucosidases I, II, and III in Organs of European Honeybees, Apis mellifera L., and the Origin of α-Glucosidase in Honey. Biosci. Biotechnol. Biochem. 2004, 68, 2346–2352. [Google Scholar] [CrossRef] [PubMed]
- Shaw, K.L.; Grimsley, G.R.; Yakovlev, G.I.; Makarov, A.A.; Pace, C.N. The Effect of Net Charge on the Solubility, Activity, and Stability of Ribonuclease Sa. Protein Sci. 2001, 10, 1206–1215. [Google Scholar] [CrossRef]
- Takahashi, Y.; Yoshida, I.; Yokozeki, T.; Igarashi, T.; Fujita, K. Investigation of Foreign Amylase Adulteration in Honey Distributed in Japan by Rapid and Improved Native PAGE Activity Staining Method. J. Appl. Glycosci. 1999, 70, 67–73. [Google Scholar] [CrossRef]
- Kimura, A.; Takewaki, S.-I.; Matsui, H.; Kubota, M.; Chiba, S. Allosteric Properties, Substrate Specificity, and Subsite Affinities of Honeybee α-Glucosidase I. J. Biochem. 1990, 107, 762–768. [Google Scholar] [CrossRef]
- Liu, J.; Cheng, J.; Huang, M.; Shen, C.; Xu, K.; Xiao, Y.; Pan, W.; Fang, Z. Identification of an Invertase With High Specific Activity for Raffinose Hydrolysis and Its Application in Soymilk Treatment. Front. Microbiol. 2021, 12, 646801. [Google Scholar] [CrossRef] [PubMed]
- Royer, J.C.; Nakas, J.P. Simple, Sensitive Zymogram Technique for Detection of Xylanase Activity in Polyacrylamide Gels. Appl. Environ. Microbiol. 1990, 56, 1516–1517. [Google Scholar] [CrossRef] [PubMed]
- Ng, A.M.J.; Zhang, H.; Nguyen, G.K.T. Zymography for Picogram Detection of Lipase and Esterase Activities. Molecules 2021, 26, 1542. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavlović, R.; Stojanović, S.; Pavlović, M.; Drulović, N.; Vujčić, M.; Dojnov, B.; Vujčić, Z. Glycosidase Isoforms in Honey and the Honey Bee (Apis mellifera L.): Differentiating Bee- and Yeast-Derived Enzymes and Implications for Honey Authentication. Insects 2025, 16, 622. https://doi.org/10.3390/insects16060622
Pavlović R, Stojanović S, Pavlović M, Drulović N, Vujčić M, Dojnov B, Vujčić Z. Glycosidase Isoforms in Honey and the Honey Bee (Apis mellifera L.): Differentiating Bee- and Yeast-Derived Enzymes and Implications for Honey Authentication. Insects. 2025; 16(6):622. https://doi.org/10.3390/insects16060622
Chicago/Turabian StylePavlović, Ratko, Sanja Stojanović, Marija Pavlović, Nenad Drulović, Miroslava Vujčić, Biljana Dojnov, and Zoran Vujčić. 2025. "Glycosidase Isoforms in Honey and the Honey Bee (Apis mellifera L.): Differentiating Bee- and Yeast-Derived Enzymes and Implications for Honey Authentication" Insects 16, no. 6: 622. https://doi.org/10.3390/insects16060622
APA StylePavlović, R., Stojanović, S., Pavlović, M., Drulović, N., Vujčić, M., Dojnov, B., & Vujčić, Z. (2025). Glycosidase Isoforms in Honey and the Honey Bee (Apis mellifera L.): Differentiating Bee- and Yeast-Derived Enzymes and Implications for Honey Authentication. Insects, 16(6), 622. https://doi.org/10.3390/insects16060622