


Effects of Short-Term Heat Stress on the Development, Reproduction, and Demographic Parameters of Phytoseiulus persimilis (Acari: Phytoseiidae)



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Rearing of Prey and Predator
2.2. Test Arena
2.3. Experimental Design
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| APOP | Adult pre-oviposition period |
| TPOP | Total pre-oviposition period |
| L:D | Light–dark |
| RH | Relative humidity |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter and Equation | Definition |
|---|---|
| Adult pre-ovipositional period (APOP) | The period between the adult emergence and first oviposition. |
| Total pre-ovipositional period (TPOP) | The period from birth to first oviposition. |
| Age-stage-specific survival rate (sxj) | The probability that a newly laid egg will survive to age x and stage j. nxj is the number of individuals to survive to age x and stage j, and n01 is the number of newborn offspring used at the beginning of the life table study [67]. |
| Age-stage-specific fecundity (fxj) | The mean number of offspring produced by individuals at age x and stage j [39]. |
| Age-specific fecundity (mx) | The mean number of offspring produced by individuals at age x [39]. |
| Net reproductive rate (R0) | The total number of offspring that an average individual (including females, males, and those that died in the immature stage) can produce during its lifetime [68]. |
| The intrinsic rate of increase (r) | The population growth rate as time approaches infinity and the population reaches a stable age-stage distribution (SASD). The population size will increase at the rate of er per time unit. It is calculated using the Euler–Lotka equation with age indexed from 0 [68]. |
| The finite rate of increase (λ) | The population growth rate as time approaches infinity and the population reaches a stable age-stage distribution. The population size will increase at the rate of λ per time unit [68]. |
| Mean generation time (T) | The period that a population requires to increase to R0-fold of its size as the time approaches infinity and the population settles down to a stable age-stage distribution [68]. |
| Age-stage-specific life expectancy (exj) | The duration for which an individual of age x and stage j is expected to survive. s’iy is the probability that an individual of age x and stage j will survive to age i and stage y and it is calculated by assuming sxj = 1 [69]. |
| Age-stage-specific reproductive value (vxj) | The reproductive value is described as the contribution of an individual of age x and stage j to the future population. For immature stages such as the larva stage, the reproductive value is not an actual reproductive output, but rather a projection of their expected future reproductive contribution if they survive to adulthood [40]. |
References
- Jerbi-Elayed, M.; Lebdi-Grissa, K.; Foray, V.; Muratori, F.; Hance, T. Using Multiple Traits to Estimate the Effects of Heat Shock on the Fitness of Aphidius colemani. Entomol. Exp. Appl. 2015, 155, 18–27. [Google Scholar] [CrossRef]
- Jin, H.-F.; Fu, B.-L.; Qiu, H.-Y.; Yang, S.-Y.; Zhou, S.-H.; Ma, X.-T.; Li, S.-G.; Zhang, F.-P.; Tang, L.-D.; Liu, K. Effects of High Temperature Stress on the Biology and Predatory Behavior of Menochilus sexmaculates. Chin. J. Appl. Entomol. 2020, 57, 700–707. [Google Scholar] [CrossRef]
- Huey, R.B.; Kingsolver, J.G. Climate Warming, Resource Availability, and the Metabolic Meltdown of Ectotherms. Am. Nat. 2019, 194, E140–E150. [Google Scholar] [CrossRef] [PubMed]
- Seneviratne, S.I.; Donat, M.G.; Mueller, B.; Alexander, L.V. No Pause in the Increase of Hot Temperature Extremes. Nat. Clim. Change 2014, 4, 161–163. [Google Scholar] [CrossRef]
- Basarin, B.; Lukić, T.; Matzarakis, A. Review of Biometeorology of Heatwaves and Warm Extremes in Europe. Atmosphere 2020, 11, 1276. [Google Scholar] [CrossRef]
- Barriopedro, D.; García-Herrera, R.; Ordóñez, C.; Miralles, D.G.; Salcedo-Sanz, S. Heat Waves: Physical Understanding and Scientific Challenges. Rev. Geophys. 2023, 61, e2022RG000780. [Google Scholar] [CrossRef]
- Singh, D.; Yadav, A.; Singh, S.; Yadav, R.K.; Sen, M.; Yadav, A.K.; Gehlot, T.; Mishra, A.; Pandey, V.; Singh, A.K. Heat Waves and Its Impact on Crop Production and Mitigation Techniques: A Review. Int. J. Enviorn. Clim. Change 2023, 13, 377–382. [Google Scholar] [CrossRef]
- Robinson, P.J. On the Definition of a Heat Wave. J. Appl. Meteor. 2001, 40, 762–775. [Google Scholar] [CrossRef]
- Hao, X.; Wang, E.; Yan, H.; Zhao, P.; Sheng, F.; Ren, Q.; Liu, M.; Zhang, B.; Xu, X. Effects of Heat Stresses on Fitness of Three Commercial Predatory Mites. Syst. Appl. Acarol. 2024, 29, 1673–1684. [Google Scholar] [CrossRef]
- Sentis, A.; Morisson, J.; Boukal, D.S. Thermal Acclimation Modulates the Impacts of Temperature and Enrichment on Trophic Interaction Strengths and Population Dynamics. Glob. Change Biol. 2015, 21, 3290–3298. [Google Scholar] [CrossRef]
- Tscholl, T.; Nachman, G.; Spangl, B.; Walzer, A. Heat Waves Affect Prey and Predators Differently via Developmental Plasticity: Who May Benefit Most from Global Warming? Pest Manag. Sci. 2022, 78, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Acosta, N.C.; Zehr, L.N.; Snook, J.S.; Szendrei, Z.; Kalwajtys, M.; Wetzel, W.C. Heat Wave Impacts on Crop-pest Dynamics Are Dependent upon Insect Ontogeny and Plant Resistance. Ecosphere 2024, 15, e70028. [Google Scholar] [CrossRef]
- Harvey, J.A.; Heinen, R.; Gols, R.; Thakur, M.P. Climate Change-mediated Temperature Extremes and Insects: From Outbreaks to Breakdowns. Glob. Change Biol. 2020, 26, 6685–6701. [Google Scholar] [CrossRef]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of Extreme Weather Disasters on Global Crop Production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef]
- Hart, A.J.; Bale, J.S.; Tullett, A.G.; Worland, M.R.; Walters, K.F.A. Effects of Temperature on the Establishment Potential of the Predatory Mite Amblyseius californicus McGregor (Acari: Phytoseiidae) in the UK. J. Insect Physiol. 2002, 48, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Ghazy, N.A.; Suzuki, T.; Amano, H.; Ohyama, K. Effects of Air Temperature and Water Vapor Pressure Deficit on Storage of the Predatory Mite Neoseiulus californicus (Acari: Phytoseiidae). Exp. Appl. Acarol. 2012, 58, 111–120. [Google Scholar] [CrossRef]
- Skirvin, D.J.; Fenlon, J.S. The Effect of Temperature on the Functional Response of Phytoseiulus persimilis (Acari: Phytoseiidae). Exp. Appl. Acarol. 2003, 31, 37–49. [Google Scholar] [CrossRef]
- Sabelis, M.W. Modelling the Predator-Prey Interaction at the Individual Level; Biological control of two-spotted spider mites using phytoseiid predators; PUDOC: Wageningen, The Netherlands, 1982; ISBN 978-90-220-0776-1. [Google Scholar]
- Zhao, Y.; Ducharne, A.; Sultan, B.; Braconnot, P.; Vautard, R. Estimating Heat Stress from Climate-Based Indicators: Present-Day Biases and Future Spreads in the CMIP5 Global Climate Model Ensemble. Environ. Res. Lett. 2015, 10, 084013. [Google Scholar] [CrossRef]
- Zhang, G.H.; Li, Y.Y.; Zhang, K.J.; Wang, J.J.; Liu, Y.Q.; Liu, H. Effects of Heat Stress on Copulation, Fecunditiy and Longevity in Newly-Emerged Adults of the Predatory Mite Neoseiulus barkeri (Acari: Phytoseiidae). Syst. Appl. Acarol. 2016, 21, 295–306. [Google Scholar] [CrossRef]
- Yao, F.; Zheng, Y.; Ding, X.; Zhao, J.; Lu, X.; Desneux, N.; He, Y.; Weng, Q. Effects of Heat Shock on Survival and Predation of an Important Whitefly Predator, Serangium japonicum. Entomol. Exp. Appl. 2019, 167, 476–489. [Google Scholar] [CrossRef]
- Zhao, C.; Chen, H.; Guo, J.; Zhou, Z. Effects of Fluctuating Thermal Regimes on Life History Parameters and Body Size of Ophraella communa. Insects 2022, 13, 821. [Google Scholar] [CrossRef]
- Li, W.Z.; Li, H.L.; Guo, Z.K.; Shang, S.Q. Effects of Short-Term Heat Stress on the Development and Reproduction of Predatory Mite Neoseiulus barkeri (Acari, Phytoseiidae). Syst. Appl. Acarol. 2021, 26, 713–723. [Google Scholar] [CrossRef]
- Li, W.; Zhu, T.; Li, H.; Shang, S. The Effects of Short-term Heat Stress on Functional Response of Neoseiulus barkeri to Tetranychus urticae. J. Appl. Entomol. 2022, 146, 310–318. [Google Scholar] [CrossRef]
- Walzer, A.; Formayer, H.; Tixier, M.-S. Evidence of Trans-Generational Developmental Modifications Induced by Simulated Heat Waves in an Arthropod. Sci. Rep. 2020, 10, 4098. [Google Scholar] [CrossRef] [PubMed]
- Mcmurtry, J.A.; Moraes, G.J.D.; Sourassou, N.F. Revision of the Lifestyles of Phytoseiid Mites (Acari: Phytoseiidae) and Implications for Biological Control Strategies. Syst. Appl. Acarol. 2013, 18, 297–320. [Google Scholar] [CrossRef]
- Pakyari, H.; Zemek, R. The Effect of Light Cycles on the Predation Characteristics of Phytoseiulus persimilis (Acari: Phytoseiidae) Feeding on Tetranychus urticae (Acari: Tetranychidae). Plants 2025, 14, 687. [Google Scholar] [CrossRef]
- Helle, W.; Sabelis, M.W. (Eds.) Spider Mites: Their Biology, Natural Enemies, and Control; World crop pests; Elsevier Science Pub. Co.: Amsterdam, The Netherlands; New York, NY, USA, 1985; ISBN 978-0-444-42372-6. [Google Scholar]
- Tixier, M.-S. Predatory Mites (Acari: Phytoseiidae) in Agro-Ecosystems and Conservation Biological Control: A Review and Explorative Approach for Forecasting Plant-Predatory Mite Interactions and Mite Dispersal. Front. Ecol. Evol. 2018, 6, 192. [Google Scholar] [CrossRef]
- Vangansbeke, D.; Audenaert, J.; Nguyen, D.T.; Verhoeven, R.; Gobin, B.; Tirry, L.; De Clercq, P. Diurnal Temperature Variations Affect Development of a Herbivorous Arthropod Pest and Its Predators. PLoS ONE 2015, 10, e0124898. [Google Scholar] [CrossRef] [PubMed]
- Coombs, M.R.; Bale, J.S. Comparison of Thermal Activity Thresholds of the Spider Mite Predators Phytoseiulus macropilis and Phytoseiulus persimilis (Acari: Phytoseiidae). Exp. Appl. Acarol. 2013, 59, 435–445. [Google Scholar] [CrossRef]
- Voroshilov, N.V. Breeding of Thermo-Resistant Lines of Phytoseiulus (Phytoseiulus persimilis A-H). Genetika 1979, 15, 70–76. [Google Scholar]
- Walzer, A.; Nachman, G.; Spangl, B.; Stijak, M.; Tscholl, T. Trans- and within-Generational Developmental Plasticity May Benefit the Prey but Not Its Predator during Heat Waves. Biology 2022, 11, 1123. [Google Scholar] [CrossRef] [PubMed]
- Tscholl, T.; Nachman, G.; Spangl, B.; Serve, H.C.; Walzer, A. Reproducing during Heat Waves: Influence of Juvenile and Adult Environment on Fecundity of a Pest Mite and Its Predator. Biology 2023, 12, 554. [Google Scholar] [CrossRef] [PubMed]
- Tscholl, T.; Nachman, G.; Spangl, B.; Scalmani, I.; Walzer, A. Parental Exposure to Heat Waves Improves Offspring Reproductive Investment in Tetranychus urticae (Acari: Tetranychidae), but Not in Its Predator, Phytoseiulus persimilis (Acari: Phytoseiidae). Ecol. Evol. 2023, 13, e10748. [Google Scholar] [CrossRef]
- Zhu, G.; Xue, M.; Luo, Y.; Ji, G.; Liu, F.; Zhao, H.; Sun, X. Effects of Short-Term Heat Shock and Physiological Responses to Heat Stress in Two Bradysia Adults, Bradysia odoriphaga and Bradysia difformis. Sci. Rep. 2017, 7, 13381. [Google Scholar] [CrossRef]
- Sun, Y.; Yang, M.; Ye, Z.; Zhu, J.; Fu, Y.; Chen, J.; Zhang, F. Effects of High-Temperature Stress on Biological Characteristics of Coccophagus japonicus Compere. Insects 2024, 15, 801. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shen, X.-X.; Yu, X.-F.; Gou, J.-Y.; Huang, C.-Y.; Yang, M.-F. Effects of Short-Term Heat Stress on the Performance of the Predatory Gall Midge Aphidoletes aphidimyza (Diptera: Cecidomyiidae). Biocontrol Sci. Technol. 2023, 33, 190–203. [Google Scholar] [CrossRef]
- Chi, H.; Liu, H. Two New Methods for the Study of Insect Population Ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Huang, Y.; Chi, H. Age-stage, Two-sex Life Tables of Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae) with a Discussion on the Problem of Applying Female Age-specific Life Tables to Insect Populations. Insect Sci. 2012, 19, 263–273. [Google Scholar] [CrossRef]
- Lehmann, P.; Ammunét, T.; Barton, M.; Battisti, A.; Eigenbrode, S.D.; Jepsen, J.U.; Kalinkat, G.; Neuvonen, S.; Niemelä, P.; Terblanche, J.S.; et al. Complex Responses of Global Insect Pests to Climate Warming. Front. Ecol. Environ. 2020, 18, 141–150. [Google Scholar] [CrossRef]
- Aguilar-Fenollosa, E.; Jacas, J.A. Can We Forecast the Effects of Climate Change on Entomophagous Biological Control Agents? Pest. Manag. Sci. 2014, 70, 853–859. [Google Scholar] [CrossRef]
- Yao, F.-L.; You, M.-S. Impacts of Global Warming on the Interaction between Host Plants, Insect Pests and Their Natural Enemies. Chin. J. Appl. Entomol. 2012, 49, 563–572. [Google Scholar]
- Sharma, H.C.; Dhillon, M.K. Climate Change Effects on Arthropod Diversity and Its Implications for Pest Management and Sustainable Crop Production. In Agronomy Monographs; Hatfield, J.L., Sivakumar, M.V.K., Prueger, J.H., Eds.; American Society of Agronomy, Crop Science Society of America, and Soil Science Society of America, Inc.: Madison, WI, USA, 2018; pp. 595–619. ISBN 978-0-89118-358-7. [Google Scholar]
- Peñalver-Cruz, A.; Escudero-Colomar, L.A.; Alins, G.; Franco, M.E.E.; Bosch-Serra, D. Consequences of Global Warming on Apple Orchards in the Mediterranean Basin: Challenges of Insect Pest Management. Entomol. Gen. 2025, 45, 53–68. [Google Scholar] [CrossRef]
- Colinet, H.; Sinclair, B.J.; Vernon, P.; Renault, D. Insects in Fluctuating Thermal Environments. Annu. Rev. Entomol. 2015, 60, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Jafari, S.; Fathipour, Y.; Faraji, F. Temperature-dependent Development of Neoseiulus barkeri (Acari: Phytoseiidae) on Tetranychus urticae (Acari: Tetranychidae) at Seven Constant Temperatures. Insect Sci. 2012, 19, 220–228. [Google Scholar] [CrossRef]
- Yuan, X.; Wang, X.; Wang, J.; Zhao, Y. Effects of brief exposure to high temperature on Neoseiulus californicus. Ying Yong Sheng Tai Xue Bao 2015, 26, 853–858. [Google Scholar]
- Xu, Y.; Zhang, K.; Zhang, Z.-Q. Development, Survival, and Reproduction of Phytoseiulus persimilis Athias-Henriot (Acari: Phytoseiidae) Feeding on Fresh versus Frozen Eggs of Tetranychus urticae Koch (Acari: Tetranychidae). Acarologia 2023, 63, 24–30. [Google Scholar] [CrossRef]
- Lu, F.; Chen, Q.; Chen, Z.; Lu, H.; Xu, X.; Jing, F. Effects of Heat Stress on Development, Reproduction and Activities of Protective Enzymes in Mononychellus mcgregori. Exp. Appl. Acarol. 2014, 63, 267–284. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Rudolf, V.H.W.; Ma, C. Extreme Temperature Events Alter Demographic Rates, Relative Fitness, and Community Structure. Global Change Biol. 2015, 21, 1794–1808. [Google Scholar] [CrossRef]
- McCalla, K.A.; Keçeci, M.; Milosavljević, I.; Ratkowsky, D.A.; Hoddle, M.S. The Influence of Temperature Variation on Life History Parameters and Thermal Performance Curves of Tamarixia radiata (Hymenoptera: Eulophidae), a Parasitoid of the Asian Citrus Psyllid (Hemiptera: Liviidae). J. Econ. Entomol. 2019, 112, 1560–1574. [Google Scholar] [CrossRef]
- Bayu, M.S.Y.I.; Ullah, M.S.; Takano, Y.; Gotoh, T. Impact of Constant versus Fluctuating Temperatures on the Development and Life History Parameters of Tetranychus urticae (Acari: Tetranychidae). Exp. Appl. Acarol. 2017, 72, 205–227. [Google Scholar] [CrossRef]
- Vangansbeke, D.; Nguyen, D.T.; Audenaert, J.; Verhoeven, R.; Gobin, B.; Tirry, L.; De Clercq, P. Prey Consumption by Phytoseiid Spider Mite Predators as Affected by Diurnal Temperature Variations. BioControl 2015, 60, 595–603. [Google Scholar] [CrossRef]
- Doker, I.; Kazak, C.; Karut, K. Functional Response and Fecundity of a Native Neoseiulus californicus Population to Tetranychus urticae (Acari: Phytoseiidae, Tetranychidae) at Extreme Humidity Conditions. Syst. Appl. Acarol. 2016, 21, 1463. [Google Scholar] [CrossRef]
- Ferrero, M.; Gigot, C.; Tixier, M.-S.; Van Houten, Y.M.; Kreiter, S. Egg Hatching Response to a Range of Air Humidities for Six Species of Predatory Mites. Entomol. Exp. Appl. 2010, 135, 237–244. [Google Scholar] [CrossRef]
- Walzer, A.; Castagnoli, M.; Simoni, S.; Liguori, M.; Palevsky, E.; Schausberger, P. Intraspecific Variation in Humidity Susceptibility of the Predatory Mite Neoseiulus californicus: Survival, Development and Reproduction. Biol. Control 2007, 41, 42–52. [Google Scholar] [CrossRef]
- Zundel, C.; Hanna, R.; Scheidegger, U.; Nagel, P. Living at the Threshold: Where Does the Neotropical Phytoseiid Mite Typhlodromalus aripo Survive the Dry Season? Exp. Appl. Acarol. 2007, 41, 11–26. [Google Scholar] [CrossRef]
- De Courcy Williams, M.E.; Kravar-garde, L.; Fenlon, J.S.; Sunderland, K.D. Phytoseiid Mites in Protected Crops: The Effect of Humidity and Food Availability on Egg Hatch and Adult Life Span of Iphiseius degenerans, Neoseiulus cucumeris, N. californicus and Phytoseiulus persimilis (Acari: Phytoseiidae). Exp. Appl. Acarol. 2004, 32, 1–13. [Google Scholar] [CrossRef]
- Le Hesran, S.; Groot, T.; Knapp, M.; Bukovinszky, T.; Forestier, T.; Dicke, M. Phenotypic Variation in Egg Survival in the Predatory Mite Phytoseiulus persimilis under Dry Conditions. Biol. Control 2019, 130, 88–94. [Google Scholar] [CrossRef]
- Kjærsgaard, A.; Blanckenhorn, W.U.; Pertoldi, C.; Loeschcke, V.; Kaufmann, C.; Hald, B.; Pagès, N.; Bahrndorff, S. Plasticity in Behavioural Responses and Resistance to Temperature Stress in Musca domestica. Animal Beh. 2015, 99, 123–130. [Google Scholar] [CrossRef]
- Dingha, B.N.; Appel, A.G.; Vogt, J.T. Effects of Temperature on the Metabolic Rates of Insecticide Resistant and Susceptible German Cockroaches, Blattella germanica (L.) (Dictyoptera: Blattellidae). Midsouth Entomol. 2009, 2, 17–27. [Google Scholar]
- Pakyari, H.; Fathipour, Y.; Enkegaard, A. Effect of Temperature on Life Table Parameters of Predatory Thrips Scolothrips longicornis (Thysanoptera: Thripidae) Fed on Twospotted Spider Mites (Acari: Tetranychidae). J. Econ. Entom. 2011, 104, 799–805. [Google Scholar] [CrossRef]
- Chen, W.; Li, D.; Zhang, M.; Zhao, Y.; Wu, W.; Zhang, G. Cloning and Differential Expression of Five Heat Shock Protein Genes Associated with Thermal Stress and Development in the Polyphagous Predatory Mite Neoseiulus cucumeris (Acari: Phytoseiidae). Exp. Appl. Acarol. 2015, 67, 65–85. [Google Scholar] [CrossRef] [PubMed]
- Colinet, H.; Overgaard, J.; Com, E.; Sørensen, J.G. Proteomic Profiling of Thermal Acclimation in Drosophila melanogaster. Insect Biochem. Mol. Biol. 2013, 43, 352–365. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Zou, Z.; Li, P.; Lin, P. Effect of Temperature on Development and Reproduction of Neoseiulus barkeri (Acari: Phytoseiidae) Fed on Aleuroglyphus ovatus. Exp. Appl. Acarol. 2012, 56, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Chi, H. Life-Table Analysis Incorporating Both Sexes and Variable Development Rates among Individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Goodman, D. Optimal Life Histories, Optimal Notation, and the Value of Reproductive Value. Am. Nat. 1982, 119, 803–823. [Google Scholar] [CrossRef]
- Chi, H.; Su, H.-Y. Age-Stage, Two-Sex Life Tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and Its Host Myzus persicae (Sulzer) (Homoptera: Aphididae) with Mathematical Proof of the Relationship between Female Fecundity and the Net Reproductive Rate. Environ. Entomol. 2006, 35, 10–21. [Google Scholar] [CrossRef]
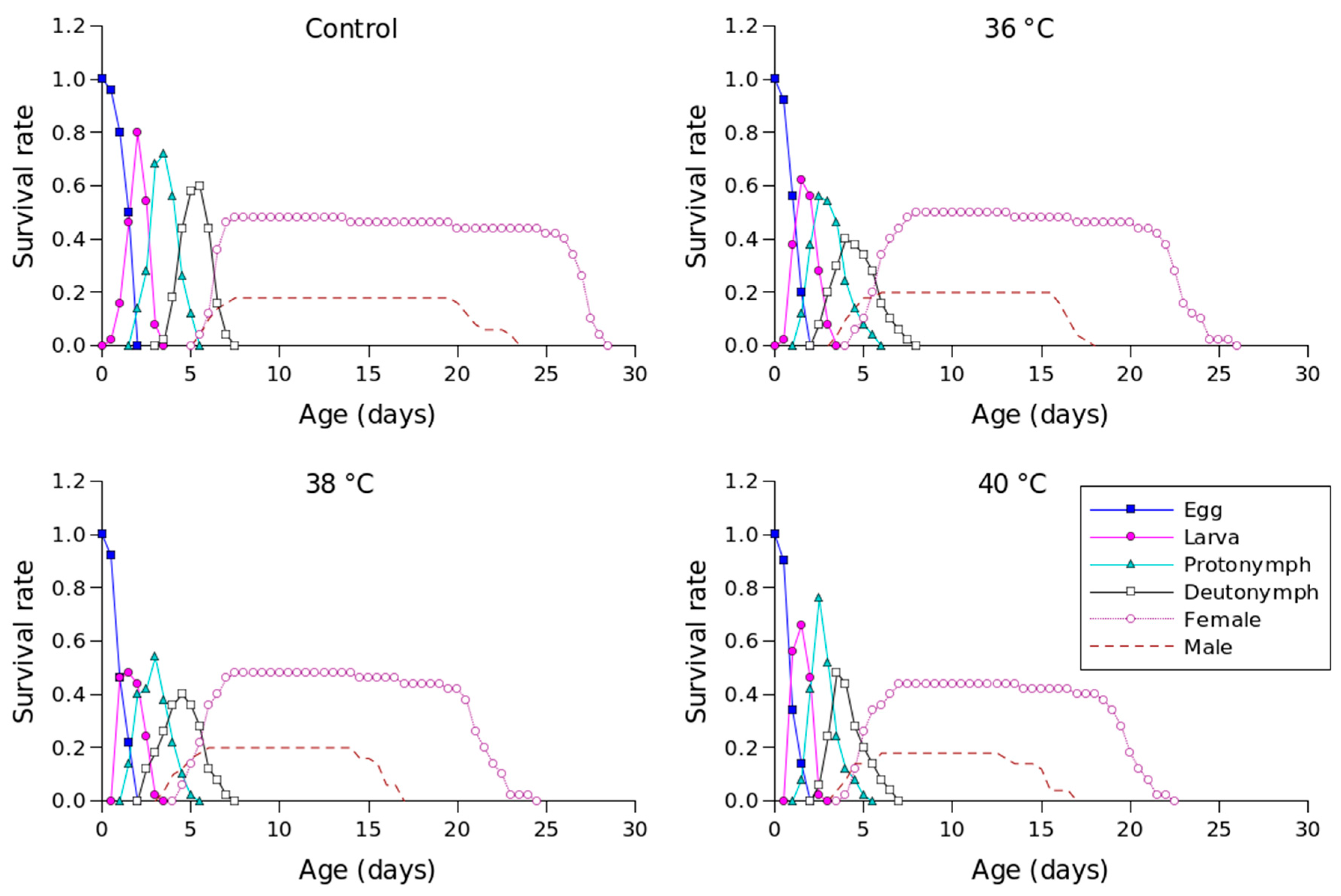
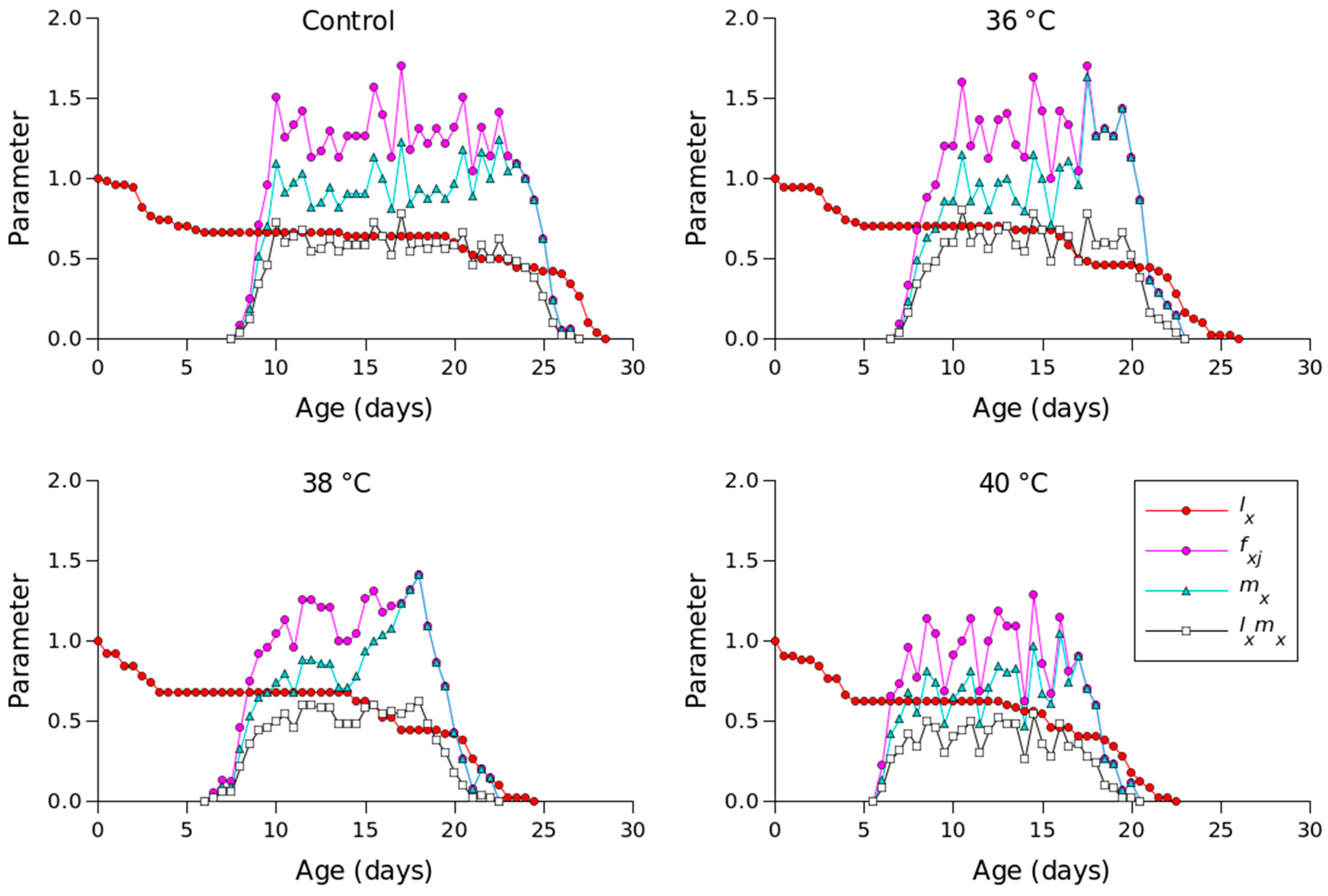
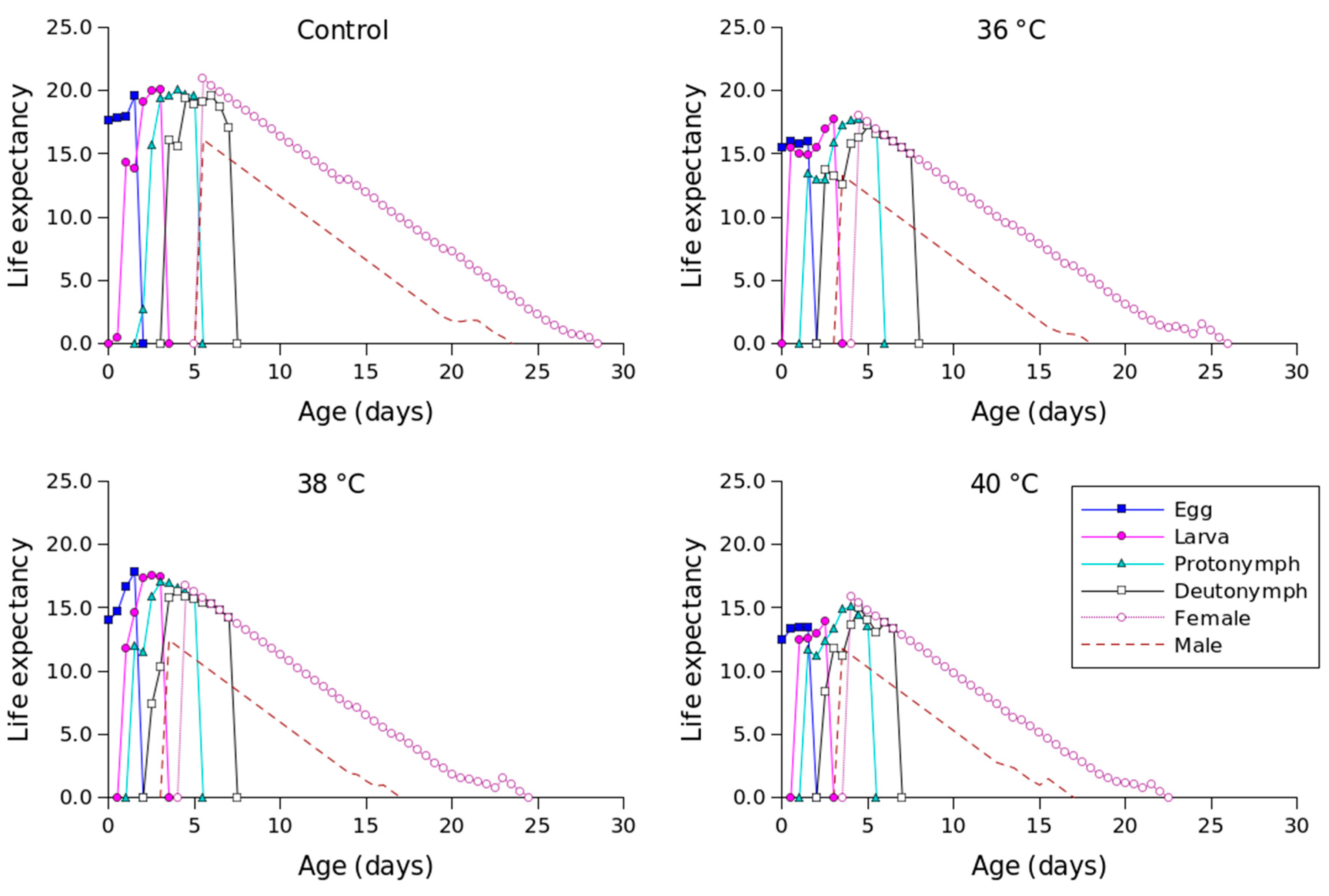
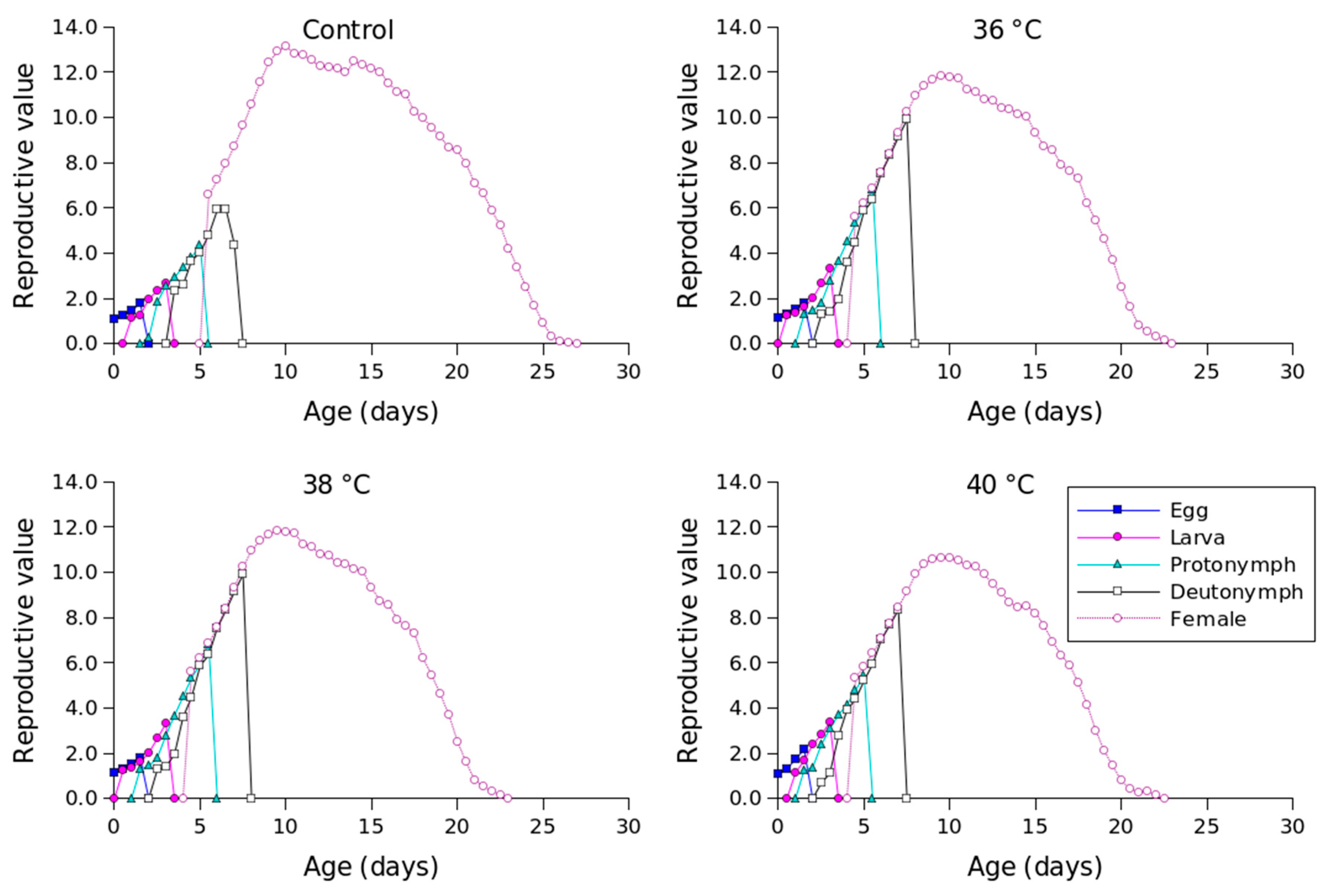
| Stage | Control | 36 °C | 38 °C | 40 °C |
|---|---|---|---|---|
| Egg (days) | 1.83 ± 0.05 a (49) | 1.52 ± 0.09 bc (47) | 1.71 ± 0.06 ab (46) | 1.39 ± 0.10 c (45) |
| Larva (days) | 1.1 ± 0.05 a (47) | 1.08 ± 0.07 a (47) | 0.93 ± 0.05 a (42) | 0.86 ± 0.07 a (44) |
| Protonymph (days) | 1.69 ± 0.09 a (37) | 1.64 ± 0.10 a (40) | 1.38 ± 0.08 a (39) | 1.45 ± 0.13 a (39) |
| Deutonymph (days) | 1.85 ± 0.05 a (33) | 1.74 ± 0.08 ab (35) | 1.56 ± 0.07 b (34) | 1.59 ± 0.08 ab (31) |
| Pre-adult (days) | 6.48 ± 0.10 a (33) | 5.98 ± 0.19 ab (35) | 5.79 ± 0.17 bc (34) | 5.30 ± 0.18 c (31) |
| Pre-adult survival (proportion) | 0.66 ± 0.06 b (33) | 0.70 ± 0.06 a (35) | 0.68 ± 0.07 ab (34) | 0.62 ± 0.07 c (31) |
| Female adult (days) | 19.92 ± 0.64 a (24) | 16.50 ± 0.48 b (25) | 15.46 ± 0.39 bc (24) | 14.55 ± 0.38 c (22) |
| Male adult (days) | 15.33 ± 0.29 a (9) | 12.40 ± 0.16 b (10) | 11.45 ± 0.23 c (10) | 10.78 ± 0.28 c (9) |
| Biological Parameter | Control | 36 °C | 38 °C | 40 °C |
|---|---|---|---|---|
| Female longevity (days) | 26.40 ± 0.64 a (24) | 22.48 ± 0.50 b (25) | 21.25 ± 0.42 bc (24) | 19.84 ± 0.38 c (22) |
| Male longevity (days) | 21.61 ± 0.45 a (9) | 16.80 ± 0.20 b (10) | 15.95 ± 0.28 c (10) | 15.28 ± 0.45 c (9) |
| Fecundity | 39 ± 1.39 a (24) | 32.08 ± 1.23 b (25) | 25.96 ± 0.77 c (24) | 21.82 ± 0.75 d (22) |
| APOP 1 (days) | 2.65 ± 0.09 a (24) | 2.70 ± 0.09 a (25) | 2.75 ± 0.06 a (24) | 2 ± 0.05 b (22) |
| TPOP 2 (days) | 9.12 ± 0.12 a (24) | 8.68 ± 0.19 b (25) | 8.54 ± 0.18 b (24) | 7.30 ± 0.19 c (22) |
| Oviposition days (days) | 14.50 ± 0.55 a (24) | 9.28 ± 0.33 b (25) | 9.31 ± 0.25 b (24) | 7 ± 0.22 c (22) |
| Sex ratio (proportion of females) | 0.48 ± 0.07 a (33) | 0.50 ± 0.07 a (35) | 0.48 ± 0.07 a (34) | 0.44 ± 0.07 a (31) |
| Population Parameter 1 | Control | 36 °C | 38 °C | 40 °C |
|---|---|---|---|---|
| r (day−1) | 0.191 ± 0.011 b | 0.205 ± 0.012 a | 0.188 ± 0.013 b | 0.194 ± 0.016 b |
| λ (day−1) | 1.211 ± 0.013 b | 1.228 ± 0.015 a | 1.206 ± 0.015 b | 1.214 ± 0.019 b |
| R0 (offspring/individual) | 18.72 ± 2.84 a | 16.04 ± 2.33 b | 12.46 ± 1.89 c | 9.60 ± 1.55 d |
| T (days) | 15.32 ± 0.20 a | 13.54 ± 0.23 b | 13.54 ± 0.23 c | 11.68 ± 0.22 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pakyari, H.; Zemek, R. Effects of Short-Term Heat Stress on the Development, Reproduction, and Demographic Parameters of Phytoseiulus persimilis (Acari: Phytoseiidae). Insects 2025, 16, 596. https://doi.org/10.3390/insects16060596
Pakyari H, Zemek R. Effects of Short-Term Heat Stress on the Development, Reproduction, and Demographic Parameters of Phytoseiulus persimilis (Acari: Phytoseiidae). Insects. 2025; 16(6):596. https://doi.org/10.3390/insects16060596
Chicago/Turabian StylePakyari, Hajar, and Rostislav Zemek. 2025. "Effects of Short-Term Heat Stress on the Development, Reproduction, and Demographic Parameters of Phytoseiulus persimilis (Acari: Phytoseiidae)" Insects 16, no. 6: 596. https://doi.org/10.3390/insects16060596
APA StylePakyari, H., & Zemek, R. (2025). Effects of Short-Term Heat Stress on the Development, Reproduction, and Demographic Parameters of Phytoseiulus persimilis (Acari: Phytoseiidae). Insects, 16(6), 596. https://doi.org/10.3390/insects16060596