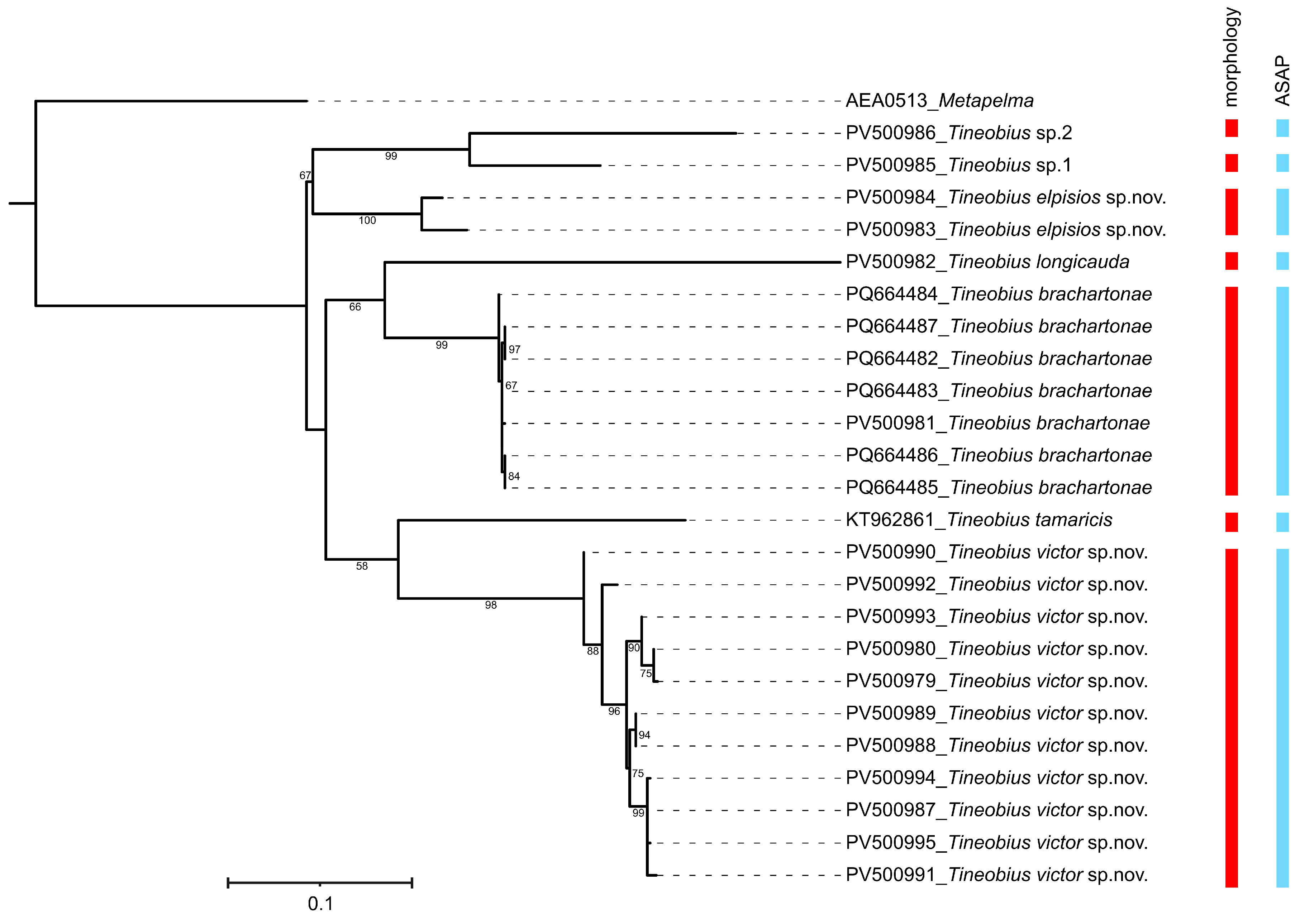
3.3.1. Tineobius (Tineobius) brachartonae (Gahan, 1927) n. rec. (Figure 2, Figure 3 and Figure 4)
Anastatoidea brachartonae Gahan, 1927: 13–15 [
25].
Tineobius brachartonae; Bouček, 1988: 564 [
4].
Tineobius (Tineobius) brachartonae; Fusu & Ribes, 2017: 12 [
2].
Figure 2.
Tineobius brachartonae (Gahan, 1927) n. rec.: (A) body, lateral; (B) body, dorsal, (C) head, dorsal; (D) head, frontal; (E) antenna; (F) mesosoma, lateral; (G) mesosoma, dorsal; (H) forewing; (I) propodeum; (J) gaster, dorsal; (K) apex of mesotibia, ventral ((A) from FAFU-DNA1108; (B–H), (J–K) from FAFU-DNA1098; (I) from FAFU-DNA 481).
Figure 2.
Tineobius brachartonae (Gahan, 1927) n. rec.: (A) body, lateral; (B) body, dorsal, (C) head, dorsal; (D) head, frontal; (E) antenna; (F) mesosoma, lateral; (G) mesosoma, dorsal; (H) forewing; (I) propodeum; (J) gaster, dorsal; (K) apex of mesotibia, ventral ((A) from FAFU-DNA1108; (B–H), (J–K) from FAFU-DNA1098; (I) from FAFU-DNA 481).
Material examined. 6♀1♂ (FAFU), Zhangjiangkou Mangrove National Nature Reserve, Yunxiao, Zhangzhou, Fujian province, China|13 January to 17 October 2022|Malaise trap/DNA1081, 1098, 1103, 1106, 1108, 1109 (♀), 1104 (♂); 1♀ (FAFU), Daiyunshan National Nature Reserve, Dehua, Quanzhou, Fujian province, China 5|October 2015|Malaise trap/DNA481.
Figure 3.
Holotype specimen of
Tineobius brachartonae: (
A) body, lateral; (
B) label; (
C) body, dorsal; (
D) head, frontal; (
E) fore wing; (
F) mesosoma, dorsal (collection number: USNMENT 01570826; Orrell T, Informatics and Data Science Center—Digital Stewardship (2024). NMNH Extant Specimen Records (USNM, USA). Version 1.93. National Museum of Natural History, Smithsonian Institution. Occurrence dataset
https://doi.org/10.15468/hnhrg3 accessed via GBIF.org, accessed on 24 December 2024;
https://www.gbif.org/occurrence/1321900501, accessed on 24 December 2024).
Figure 3.
Holotype specimen of
Tineobius brachartonae: (
A) body, lateral; (
B) label; (
C) body, dorsal; (
D) head, frontal; (
E) fore wing; (
F) mesosoma, dorsal (collection number: USNMENT 01570826; Orrell T, Informatics and Data Science Center—Digital Stewardship (2024). NMNH Extant Specimen Records (USNM, USA). Version 1.93. National Museum of Natural History, Smithsonian Institution. Occurrence dataset
https://doi.org/10.15468/hnhrg3 accessed via GBIF.org, accessed on 24 December 2024;
https://www.gbif.org/occurrence/1321900501, accessed on 24 December 2024).
Brief description of female. Head dark, scrobal depression (
Figure 2D), and gena (
Figure 2F) with slight greenish-yellow metallic luster, interantennal prominence and purple to reddish-purple lower face, with short, translucent, scalelike setae (
Figure 2D). Antenna (
Figure 2E) with yellowish-brown scape, pedicel blue-metallic luster, flagellum black. Pronotum black with strong-blue metallic luster. Mesonotum (
Figure 2G) dark, except posterior depression region with strong bluish-green and coppery metallic lusters. Acropleuron (
Figure 2F) bare, anterior region with alutaceous sculpture and posterior mesh-like longitudinally alutaceous coriaceous sculpture. Mesopectus with intensely white hair-like setae. Propodeum (
Figure 2I) slightly emarginate anteromedially (cf. Gibson 1995, Figure 201) to anterior margin straight (cf. Gibson 1995, Figure 202) [
1], plical region transverse almost without depression. Legs mostly dark-brown to black, anterior and median coxa mostly testaceous, hind coxa purple. Mesotibia with 10–13 apical pegs (
Figure 2K) in 2 or 3 rows. Basal cell of forewing bare (
Figure 2H), setae below marginal and stigma vein dense-brown and scalelike, beyond apex of stigma vein hair-like. Gaster (
Figure 2J) dark, ovipositor sheaths dark, almost equal in length to gaster.
Description of male. Body length about 2.3 mm. Head (
Figure 4D,E) with green-and-blue metallic lusters, gena and interantennal prominence with purplish-blue metallic luster, scrobal depression with green to greenish-yellow metallic luster, green lower face with greenish-yellow metallic lusters, dark-brown maxillary palpi, apical paler, frons and vertex dark with slightly metallic luster. Eyes and frons (
Figure 4E) densely setose with brown hair-like setae, gena and lower face (
Figure 4D) densely setose with pale-gray hair-like setae. Frons coriaceous, scrobal depression and interantennal prominence reticulate–imbricate transversely, lower face and gena imbricate. In frontal view, wider head 1.1× height, distance between eyes below 2.6× distance between eyes above, distance between toruli 2.0× distance between torulus and clypeal edge, and 3.3× distance between torulus and eye. In dorsal view, head wider 1.6× length, interocular distance 0.3× head width. In lateral view, malar space 0.3× height of eye. OOL: POL: LOL: MPOD = 1.0: 7.1: 7.4: 6.0. Scrobal depression ^-shaped with dorsal margin wide. Antenna (
Figure 4B) dark in color, scape and pedicel with slightly greenish-yellow metallic luster; relative length (width) of scape: 56.8 (18.3), pedicel: 20.5 (12.7), 1st to 8th flagellomeres: 5.4 (10.0), 10.7 (12.2), 12.0 (13.2), 10.7 (13.9), 12.2 (13.7), 12.0 (14.4), 12.0 (14.4), 14.1 (16.3), and clava 32.7 (17.6).
Mesosoma (
Figure 4F,G) generally with greenish-blue metallic luster, notaulus conspicuous, mesoscutal medial-lobe region triangular with greenish-blue metallic luster, dark mesoscutal lateral lobe ranging from greenish-yellow to purple metallic lusters, dark-purple scutellum with greenish-blue metallic luster, acropleuron, mesepisternum, mesepimeron, and metapleuron bare, with variable greenish-blue to greenish-yellow metallic lusters from different angles. Axilla and scutellum non-separated, scutellum convex significantly with evenly distributed dark hair-like setae at the front end, lateral sides of pronotum and mesoscutum (
Figure 4F) with densely brown setae; prepectus (
Figure 4G) reticulate–imbricate with blue metallic luster. Pronotum and mesoscutum reticulate, mesepisternum, acropleuron, mesepimeron and metapleuron reticulate–imbricate. Propodeum (
Figure 4H) with variable greenish-blue metallic luster, coriaceous sculptured, several long white setae around spiracle. Forewing (
Figure 4C) transparent, light-brown stigma vein, lighter than marginal vein, evenly setose, basal cell with three rows of setae along mediocubital fold, relative length of cc: mv: pmv: stv = 3.7: 2.4: 1.4: 1.0. Legs (
Figure 4A) dark except mesotarsus (
Figure 4I) and metatarsus pale gray to dark brown, first and second segments pale gray, third, fourth, and fifth segments dark brown, mesotibia spur pale gray.
Gaster dark with variable coppery metallic luster from different angles, except Gt1 with distinct blue metallic luster.
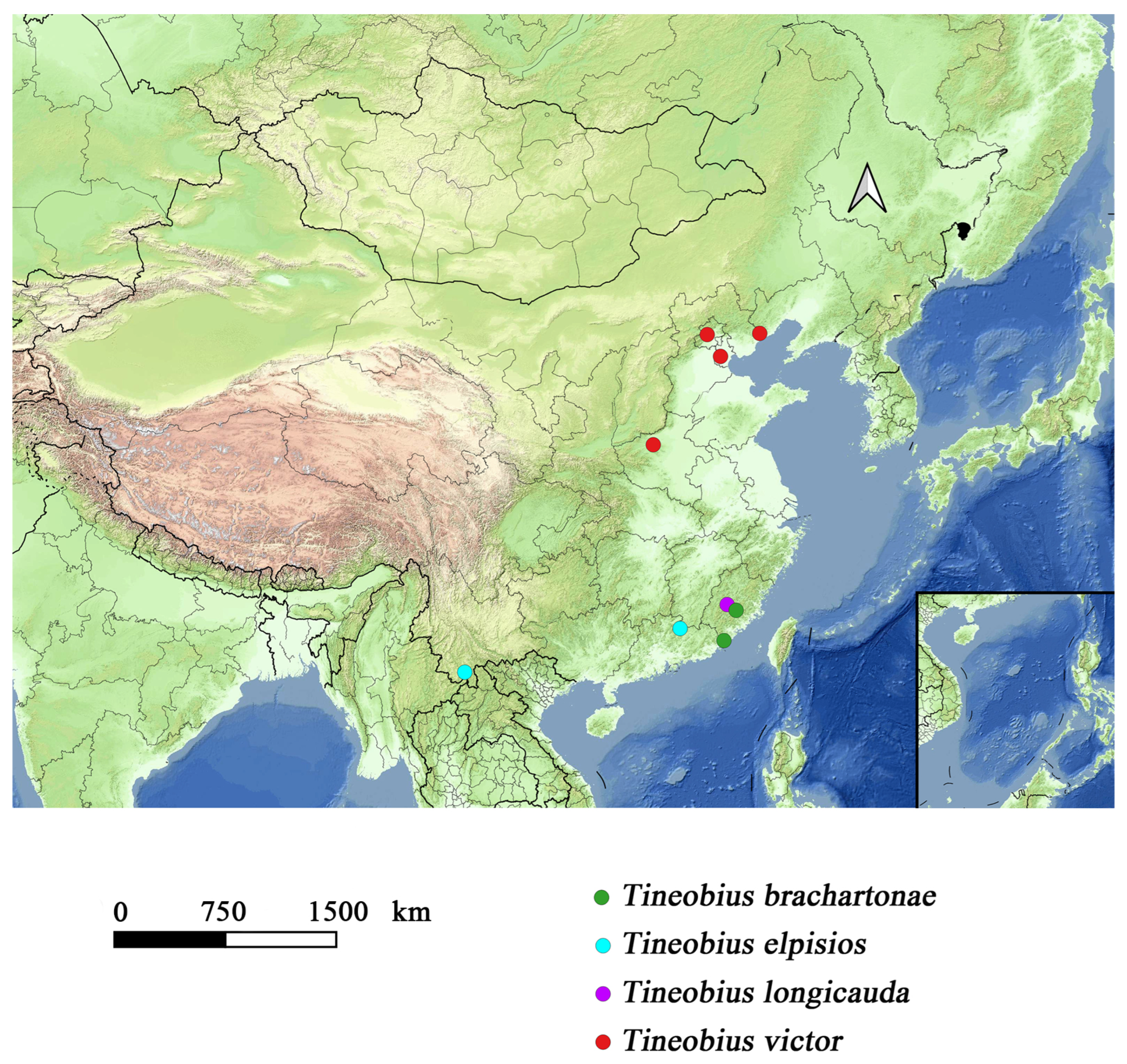
Distribution. *China (Fujian), Malaysia, Sri Lanka, Indonesia.
Observation record: China (Taiwan).
Biology. According to the original description, this species is a primary parasitoid of the larvae or pupae of Brachartona catoxantha Hampson, 1893 (Lepidoptera: Zygaenidae) in Indonesia. When functioning as a secondary parasitoid, it parasitizes Degeeria albiceps Macquart, 1851 (Diptera: Tachinidae), Bessa remota (Aldrich, 1925) (Diptera: Tachinidae), Apanteles sp. (Hymenoptera: Braconidae), and an Ichneumonidae species. The host of this species in China is unknown.
Figure 4.
Tineobius brachartonae (Gahan, 1927) n. rec. (FAFU-DNA1104): (A) body, dorsal; (B) antenna; (C) forewing; (D) head, frontal; (E) head, dorsal; (F) mesosoma, dorsal; (G) mesosoma, lateral; (H) propodeum; (I) apex of mesotibia, dorsal.
Figure 4.
Tineobius brachartonae (Gahan, 1927) n. rec. (FAFU-DNA1104): (A) body, dorsal; (B) antenna; (C) forewing; (D) head, frontal; (E) head, dorsal; (F) mesosoma, dorsal; (G) mesosoma, lateral; (H) propodeum; (I) apex of mesotibia, dorsal.
Remarks. We examined the type specimens through publicly accessible data hosted on the Global Biodiversity Information Facility (GBIF). The forewing of the type specimens has a special-shaped setae (
Figure 3E), and also white bristles of a similar shape on the face (
Figure 3D). Gahan [
25] described these setae as scalelike and mentioned that a similar type of setae is also present in
Tineobius superbus (Dodd, 1917), which is from Townsville, Australia. Upon a careful comparison of Gibson’s description of the head (cf. pp. 284, 288, 291, 327, Figures 41 and 42) [
1] and Fusu and Ribes’s description [
2] of the wings (cf. pp. 8–9), we interpret the term “scalelike” as having the same meaning as “spatulate”. Thus, the known species of
Tineobius in Asia that also have scalelike (spatulate) setae on their forewings include
Tineobius philippinensis (Ferrière, 1938) and
Tineobius indicus (Ferrière, 1938).
Gahan provided a detailed description of the species based on four female specimens. In this study, we identified a male of this species based on COI analysis.
3.3.2. Tineobius (Tineobius) elpisios Li & Peng sp. nov. (Figure 5 and Figure 6)
Figure 5.
Tineobius (Tineobius) elpisios Li & Peng sp. nov.: (A) body, lateral; (B) body, lateral; (C) mesosoma, dorsal; (D) mesosoma, dorsal; (E) mesosoma, lateral; (F) mesosoma, lateral ((A,C,E) from FAFU-DNA962, holotype; (B,D,F) from FAFU-DNA1062, paratype).
Figure 5.
Tineobius (Tineobius) elpisios Li & Peng sp. nov.: (A) body, lateral; (B) body, lateral; (C) mesosoma, dorsal; (D) mesosoma, dorsal; (E) mesosoma, lateral; (F) mesosoma, lateral ((A,C,E) from FAFU-DNA962, holotype; (B,D,F) from FAFU-DNA1062, paratype).
Figure 6.
Tineobius (Tineobius) elpisios Li & Peng sp. nov. (holotype, FAFU-DNA962): (A) head, frontal; (B) head, dorsal; (C) forewing; (D) antenna; (E) propodeum; (F) apex of gaster, ventral.
Figure 6.
Tineobius (Tineobius) elpisios Li & Peng sp. nov. (holotype, FAFU-DNA962): (A) head, frontal; (B) head, dorsal; (C) forewing; (D) antenna; (E) propodeum; (F) apex of gaster, ventral.
Type material. Holotype: 1♀ (FAFU), Jiulianshan National Nature Reserve of Jiangxi, Jiangxi province, China|29 December 2021 to 12 February 2022|Malaise trap/DNA962. Paratypes: 1♀ (FAFU), same data as holotype|10 October 2020|Broken/DNA950; 1♀ (FAFU), Ji’nuo mount, Jinghong, Xishuangbanna, Yunnan province, China|1157 m|Flight interception trap (Black)/DNA1062.
Etymology. The name is derived from the Latin word for ‘elpis’ (‘hope’), we hope that we will never lose infinite hope, regardless of time or place.
Description of female. Holotype (
Figure 5A,C,E and
Figure 6), body length about 3.9 mm (ovipositor excluded). Head (
Figure 6A,B) with green metallic luster except frons dark, vertex blue with green and coppery metallic lusters lower portion of scrobal depression with blue metallic luster, lower face coppery, dark-brown maxillary palpi. Setae of eyes and lower face white and lanceolate, setae of frons, gena and vertex white and hair-like. Vertex and temple rugose, frons mesh-like coriaceous to reticulate, scrobal depression and interantennal prominence reticulate–imbricate. In frontal view, head wider 1.1× height, distance between eyes below 0.3× distance between eyes above, distance between toruli 2.3× distance between torulus and clypeal edge, and 2.3× distance between torulus and eye. In dorsal view, head wider 1.8× length, interocular distance 0.22× head width. In lateral view, malar space 0.4× height of eye. OOL: POL: LOL: MPOD = 1.0: 4.5: 5.2: 3.8. Scrobal depression ∩-shape. Antenna (
Figure 6D) dark brown in color except scape yellowish-brown, pedicel with slightly greenish-blue metallic luster; relative length (width) of scape: 93.2 (12.1), pedicel: 30.5 (10.2), 1st to 8th flagellomeres: 15.4 (10.0), 26.3 (12.0), 25.0 (12.5), 23.4 (12.9), 22.3 (14.8), 20.9 (16.4), 19.3 (17.7), 17.0 (17.9); clava 43.9 (20.0).
Pronotum (
Figure 5C) and anterior region of acropleuron (
Figure 5E) with yellowish-green and blue metallic lusters, posterior region of acropleuron dark, anterior convex region of mesoscutal medial lobe (
Figure 5C) reddish to greenish, posterior concave region of mesoscutal medial lobe sky-blue, propodeum (
Figure 5E) with bluish-purple metallic luster. Setae of pronotum, mesonotum notaulus, posterior concave region of mesoscutal medial lobe, anterior region of acropleuron and mesopectus white hair-like, posterior region of acropleuron bare. Pronotum divided medially, coriaceous to reticulate. Mesoscutum with anterior convex region of mesoscutal medial lobe punctate reticulate sculptured, lateral lobe coarsely coriaceous, acropleuron with anterior region reticulate–imbricate sculpture and posterior longitudinally alutaceous sculpture, mesoscutellar–axillar punctate–reticulate. Metanotum (
Figure 6E) convex, propodeum emarginate anteromedially, plical region transverse with V-shaped plical depression, interior coriaceous sculptured. Forewing (
Figure 6C) extended to near apex of gaster, basal region hyaline and partly bare, except cubital area of forewing with dense-brown hair-like setae. Setae below marginal and stigma vein brown, dense, hair-like to scalelike, beyond apex of stigma vein unobvious light short hair-like, relative length of cc: mv: pmv: stv = 3.4: 3.1: 2.2: 1.0. Front leg dark-brown; middle leg dark to brown except coxa with yellowish-green to blue metallic luster, tarsus white to dark brown, mesotibia with 9 apical pegs in 2 rows with spur brown, tarsus pale to dark, first to third joint pale, fourth and fifth brown; hind leg similar in color to middle leg, but coxa with coppery to yellowish-green metallic luster, tarsus dark to white to light brown.
Gaster (
Figure 6F) brown with slightly blue and green metallic luster from different angles; Mt
6 with posterior margin straight, posterior of Mt
7 convex. Syntergum with posterior margin omega-like emarginated. Ovipositor sheath length about 1.6 mm, light brown to dark brown, as long as gaster, in posterior part with white band.
Variation. Body length is 3.6–3.9 mm, the ovipositor sheath length is in the range of 1.6–2.1 mm. cc: mv: pmv: stv = 3.4–3.7: 3.1–3.5: 2.2–2.3: 1.0, OOL: POL: LOL: MPOD= 1.0: 4.5–5.1: 5.2–5.7: 3.8–4.3. Relative length (and width) of antenna is: scape 93.2–95.8 (11.8–12.1); pedicel: 27.5–30.5 (10.2–10.5); and 1st to 8th flagellomeres: 14.9–15.4 (10.0), 26.0–26.3 (10.9–12.0), 24.4–25.0 (12.2–12.5), 22.5–23.4 (12.9–13.5), 21.1–22.3 (14.5–14.8), 19.8–20.9 (15.5–16.4), 17.5–19.3 (16.5–17.7), 15.6–17.0 (17.3–17.9); clava: 43.8–43.9 (19.8–20.0).
Distribution. China (Jiangsu, Yunnan).
Figure 8.
Tineobius (Tineobius) victor Li & Peng sp. nov. (holotype, FAFU-DNA1111): (A) body, lateral; (B) head, dorsal; (C) head lateral and antenna; (D) head, frontal; (E) mesosoma, dorsal; (F) mesosoma, lateral; (G) propodeum; (H) forewing; (I) apex of mesotibia, dorsal; (J) gaster, dorsal; (K) apex of mesotibia, ventral.
Figure 8.
Tineobius (Tineobius) victor Li & Peng sp. nov. (holotype, FAFU-DNA1111): (A) body, lateral; (B) head, dorsal; (C) head lateral and antenna; (D) head, frontal; (E) mesosoma, dorsal; (F) mesosoma, lateral; (G) propodeum; (H) forewing; (I) apex of mesotibia, dorsal; (J) gaster, dorsal; (K) apex of mesotibia, ventral.
Type material. Holotype: 1♀ (FAFU), Beihang University (Shahe Campus), Beijing, China|3 September 2024|Zhenshung Huang leg./DNA1111. Paratypes: 5♀ 2♂ (FAFU), same data as holotype|7 October 2024/DNA1112, 1113, 1114, 1123, 1124 (♀), 1125, 1126 (♂); 1♀ (FAFU), Lu’er’bao sika deer breeding base, Qinhuangdao, Hebei province|15 August 2024|Mingxuan Wu leg./DNA1102; 4♀ (FAFU), Sujiazhuang village, Sweeping, Gongyi, Henan province|all ovipositor sheathes broken|26 February 2024|Weiqiong Li leg./DNA1139, 1140, 1141, 1142; 1♀ (FAFU), jujube yards, Tianjin agricultural university, Tianjin|June 2024|Weiqiong Li leg.|Malaise trap/DNA1143.
Etymology. The name is derived from the Latin word for “victor” and was chosen to commemorate the 80th anniversary of the victory of anti-fascism in World War II. It specifically honors the immense sacrifices and contributions made by the people of the world during the war.
Description of female. Holotype. Body length about 4.9 mm (ovipositor excluded). Head (
Figure 8B–D) green with yellow metallic luster except vertex (
Figure 8B) with some slightly purple metallic luster, frons with obvious deep-blue to purple metallic lusters, lower portion of scrobal depression (
Figure 8D) with sky-blue metallic luster, interantennal prominence with purple metallic luster, dark-brown maxillary palpi. Setae of vertex and occiput dark, hair-like; setae of gena and lower face white and lanceolate (
Figure 8C). Frons, vertex, and temple rugose; scrobal depression (
Figure 8D), interantennal prominence and lower-face region reticulate. In frontal view, head wider 1.2× height, distance between eyes below 0.4× distance between eyes above, distance between toruli 1.6× distance between torulus and clypeal edge, and 1.8× distance between torulus and eye. In dorsal view, head wider 1.8× length, interocular distance 0.29× head width. In lateral view, malar space 0.4× height of eye. OOL: POL: LOL: MPOD = 1.0: 4.5: 3.9: 3.7. Scrobal depression ^-shape, dorsally delimited. Antenna (
Figure 8C) dark in color, anterior region of third flagellomere with constriction; relative length (width) of scape 75.8 (13.1), pedicel 18.0 (10.3), 1st to 8th flagellomeres: 14.9 (10.0), 38.6 (9.9), 32.8 (10.8), 29.6 (11.5), 22.8 (13.5), 21.3 (13.9), 17.9 (14.1), 15.4 (13.5); clava 36.6 (19.3).
Mesosoma (
Figure 8E–G) reddish to orangish brown except pronotum with green and blue metallic lusters, posterior depressed region of mesoscutal medial lobe pinkish-purple, propodeum with yellowish-green metallic luster. Setae of pronotum and anterior lobe white, hair-like, setae of posterior lobe white lanceolate; setae of lateral lobe and axillae–scutellum brown hair-like; setae of mesopectus and propodeum densely hair-like. Pronotum divided medially, reticulate sculptured. Mesoscutum (
Figure 8E) with anterior convex region of mesoscutal medial lobe coriaceous, depressed posterior close to smooth, lateral lobe slightly coriaceous. Mesoscutellar–axillar complex coarsely punctate reticulate. Prepectus bare, mesh-like coriaceous. Acropleuron (
Figure 8F) bare, with anterior region punctate sculpture and posterior alutaceous sculpture. Metanotum not convex, propodeum (
Figure 8G) anterior margin straight, plical region transverse with ℧-shaped plical depression, interior coriaceous sculptured. Forewing (
Figure 8H) extended to near apex of gaster, basal cell hyaline with sparse brown setae, below margin vein to postmarginal vein infuscated with relatively long and dense brown hair-like setae, beyond apex of postmarginal hyaline with very short and fine setae; one row of dark setae upright in submarginal vein, about 3 rows of dark setae of parastigma of submarginal vein to postmarginal vein densely, costal cell with 3 rows of short and fine dark setae, margin of costal cell with 1 row of dark setae, relative length of cc: mv: pmv: stv = 3.3: 2.2: 2.2: 1.0. Leg (
Figure 8A) dark to brown except coxa with yellowish-green to bluish-purple metallic luster, hindcoxa bluish-purple obviously, mesotibia with 15 apical pegs (
Figure 8K) in 2 rows and lateral with spur brown, tarsus pale to dark, first joint pale, second and third light brown, fourth and fifth dark (
Figure 8I); apex of metafemur testaceous different with metatibia, metatibia and metabasitarsus compressed.
Gaster (
Figure 8J) black with slightly blue metallic luster from different angles; Mt
6 with posterior margin slightly angulate, Mt
7 partly concealed beneath Mt
6. Syntergum with posterior margin omega-like emarginated. Ovipositor sheath length about 3.6 mm, obviously longer than the abdomen, dark with white band, which is in posterior part of ovipositor sheath, about 1/6 of total length of ovipositor sheath.
Figure 9.
Tineobius (Tineobius) victor Li & Peng sp. nov. (FAFU-DNA1125): (A) body, lateral; (B) antenna; (C) head, frontal; (D) head, dorsal; (E) propodeum; (F) mesosoma, dorsal; (G) mesosoma, lateral; (H) forewing; (I) front leg.
Figure 9.
Tineobius (Tineobius) victor Li & Peng sp. nov. (FAFU-DNA1125): (A) body, lateral; (B) antenna; (C) head, frontal; (D) head, dorsal; (E) propodeum; (F) mesosoma, dorsal; (G) mesosoma, lateral; (H) forewing; (I) front leg.
Description of male. Body length about 3.4 mm. Head (
Figure 9C,D) generally greenish-yellow with metallic luster, vertex (
Figure 9D) with coppery metallic luster, scrobal depression with greenish-yellow metallic luster, interantennal prominence (
Figure 9C) dark verified with other, gena region with variably greenish-blue metallic luster, maxillary and labial palpi dark brown. Setae of eyes and frons brown, hair-like, setae of vertex dark, setae of gena and lower face densely white, hair-like. Frons mesh-like reticulate to rugose, scrobal depression and interantennal prominence reticulate, lower face and gene striate-reticulate. In frontal view, head wider 1.3× height, distance between eyes below 2.0× distance between eyes above, distance between toruli 1.5× distance between torulus and clypeal edge, and 2.4× distance between torulus and eye. In dorsal view, head wider 1.7× length, interocular distance 0.4× head width. In lateral view, malar space 0.4× height of eye. OOL: POL: LOL: MPOD = 1.0: 4.1: 2.8: 2.5. Scrobal depression ∩-shape (
Figure 9C). Antenna (
Figure 9B) shorter than female, brown except scape and pedicel with bluish-purple metallic luster; relative length (width) of scape 42.5 (18.9), pedicel 17.5 (11.3), 1st to 8th flagellomeres: 5.7 (10.0), 19.4 (13.0), 15.5 (17.4), 15.7 (19.1), 15.7 (19.1), 15.3 (19.2), 15.8 (19.4), 15.8 (18.5); clava 32.1 (16.8).
Mesosoma (
Figure 9F,G) generally with greenish-yellow metallic lusters, pronotum with greenish-blue metallic luster, mesoscutum (
Figure 9F) greenish-yellow with greenish-blue metallic luster and densely short-brown setae, scutellum dark with greenish-brown metallic luster. Pronotum divided medially with densely lateral short-brown setae, mesepimeron and metapleuron with variably greenish-yellow to purplish-blue metallic luster and bare, mesepisternum dark and bare. Mesoscutum and axillae rugose, mesoscutellar coriaceous to reticulate, prepectus (
Figure 9G) imbricate with greenish-yellow metallic lusters, mesepisternum, acropleuron coriaceous sculptured, mesepimeron and metapleuron coriaceous to smooth. Propodeum (
Figure 9E) with variably greenish-yellow to greenish-blue metallic luster from different angles, coriaceous sculptured, densely white setae on the lateral side of the spiracle. Forewing (
Figure 9H) transparent, unevenly covered brown setae, setae of basal cell sparser than other, relative length of cc: mv: pmv: stv = 3.9: 2.3: 3.2: 1.0. Front leg (
Figure 9I) dark with variably greenish-yellow to purplish-blue metallic lusters, with white setae, tarsus white to dark brown, first region white, second and third pale gray, fourth and fifth dark brown, spur dark brown; middle leg similar in color to front leg, except mesotibia spur pale yellow; hind coxa similar with female in color.
Gaster (
Figure 9A) dark with variably coppery or greenish-bronze metallic luster from different angles, last of gaster with blue metallic luster.
Variation. Female: Obvious color variations (
Figure 10) are observed across the head, pronotum, mesonotum, axillae–scutellum, and propodeum, the coloration varies from bluish-grey to reddish-brown, and sometimes exhibits a green metallic luster. Certain specimens, such as DNA 1102 and 1143, differed by the lack of a distinct white band outside the ovipositor sheath. These variations are observed across the specimens, leading us to consider them as intraspecific rather than interspecific variations. The body length ranges from 3.8 to 5.3 mm, with the ovipositor sheath length ranges from 2.5 to 3.8 mm, larger individuals possess a longer ovipositor sheath. cc: mv: pmv: stv = 3.2–3.8: 1.9–2.7: 1.5–2.4: 1.0, OOL: POL: LOL: MPOD = 1.0: 3.9–4.6: 3.0–3.7: 2.3–3.7. Relative length (and width) of antenna is: scape 76.6–97.1 (13.2–17.3); pedicel 21.1–30.5 (9.8–11.9); and 1st to 8th flagellomeres: 11.1–17.9 (10.0), 30.2–43.8 (8.9–10.9), 29.6–40.2 (9.6–13.4), 27.2–34.3 (11.8–15.4), 23.5–29.0 (12.6–15.9), 21.9–26.1 (12.8–16.7), 17.1–22.3 (14.0–17.7), 13.7–22.9 (14.2–18.5); clava 27.0–46.1 (13.5–25.8). Mesotibia with 9–16 apical pegs in 2 or 3 rows.
Figure 10.
Tineobius (Tineobius) victor Li & Peng sp. nov variation in colour: (A) head, frontal; (B) head, frontal; (C) mesosoma, dorsal; (D) mesosoma, dorsal; (E) mesosoma, dorsal; (F) mesosoma, lateral; (G) mesosoma, lateral; (H) mesosoma, lateral ((A,C,F) from FAFU-DNA1142; (D,G) from FAFU-DNA1114; (B,E,H) from FAFU-DNA1102).
Figure 10.
Tineobius (Tineobius) victor Li & Peng sp. nov variation in colour: (A) head, frontal; (B) head, frontal; (C) mesosoma, dorsal; (D) mesosoma, dorsal; (E) mesosoma, dorsal; (F) mesosoma, lateral; (G) mesosoma, lateral; (H) mesosoma, lateral ((A,C,F) from FAFU-DNA1142; (D,G) from FAFU-DNA1114; (B,E,H) from FAFU-DNA1102).
Male: OOL: POL: LOL: MPOD = 1.0: 4.1: 2.6: 2.2; relative length (width) of scape 46.5 (17.7), pedicel 14.6 (10.5), 1st to 8th flagellomeres: 4.4 (10.0), 15.3 (12.5), 13.9 (15.3), 13.0 (18.1), 14.0 (18.2), 13.3 (17.2), 13.7 (17.2), 12.8 (16.3); clava 30.4 (15.1).
Distribution. China (Beijing, Hebei, Henan, Tianjing).
Observation record: China (Jiangsu).
Biology. All specimens from Beijing were collected from trees of
Populus ×
canadensis. The collector observed oviposition behavior of this species in the field and noticed the male in the same tree (
Figure 11). After collecting specimens of wasps, an empty cocoon along with a suspected head capsule of a Lepidoptera insect were discovered behind the bark in the oviposition area. During the period of this wasp generation (June to October), the collector conducted long-term observations at the collection site. Based on the previous research on the host of
Tineobius [
1,
2,
25], we list some of the species he observed around the tree that might be related to this new species: two species of Gelechioidea (Lepidoptera) (
Figure 12),
Eophyllophila sp. and
Dolichopodomintho sp. (Diptera: Tachinidae), and one species of Eurytomidae, one species of Braconidae,
Cheiloneurus sp. (Hymenoptera: Encyrtidae), and
Dirhinus sp. (Hymenoptera: Chalcididae)
Figure 11.
Tineobius (Tineobius) victor Li & Peng sp. nov.: (A) female; (B) male. Photo by Zhenshuang Huang, 7 October 2024.
Figure 11.
Tineobius (Tineobius) victor Li & Peng sp. nov.: (A) female; (B) male. Photo by Zhenshuang Huang, 7 October 2024.
Figure 12.
potential host of T. victor beneath the bark (A) adult of Gelechioidea gen. sp. 1; (B) adult of Gelechioidea gen. sp. 2; (C) pupa of Gelechioidea gen. sp. Photo by Zhenshuang Huang, June to September, 2024.
Figure 12.
potential host of T. victor beneath the bark (A) adult of Gelechioidea gen. sp. 1; (B) adult of Gelechioidea gen. sp. 2; (C) pupa of Gelechioidea gen. sp. Photo by Zhenshuang Huang, June to September, 2024.
Diagnosis. The coloration of this new species resembles that of
Tineobius tamaricis Ribes & Fusu 2017,
Tineobius indicus (Ferrière, 1938) and
Tineobius philippinensis (Ferrière, 1938). The differences between this new species and
Tineobius philippinensis and
Tineobius indicus are shown in our key. Furthermore, it can be distinguished from
Tineobius tamaricis by the following combination of characteristics: (1) The axillae–scutellum of
T. tamaricis is marked with isodiametric reticulate rugose sculpture, whereas in
T. victor, the sculpture is coarsely punctate reticulate (
Figure 8E); (2) The metanotum of
T. tamaricis is convex, while
T. victor is not (
Figure 8G); and (3) In
T. tamaricis, the setae in the basal cell region of the forewing are pale, contrasting with the brown setae in other areas, whereas in
T. victor, the setae on the forewings are uniformly brown (
Figure 8H).
Remarks. In addition to the existing specimen sources, we received a photograph (
Figure 13) from Nanjing, Jiangsu province, which is highly likely to depict this new species. We speculate that this species has a broader distribution in China and is likely widespread across the North China Plain, north of the Yangtze River.
Figure 13.
Tineobius victor in Nanjing, Jiangsu province. Photo by Zhenhao Feng, 3 June 2022.
Figure 13.
Tineobius victor in Nanjing, Jiangsu province. Photo by Zhenhao Feng, 3 June 2022.


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}