
Abundance, Diversity and Distribution of Mosquito Species and Molecular Detection of Its Associated Hepatitis C Virus in Sharkia Governorate, Egypt

 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Trips and Sample Collection
2.3. Identification of Species
2.4. Real-Time PCR Analysis
2.5. Working Protocol
2.6. Data Analysis
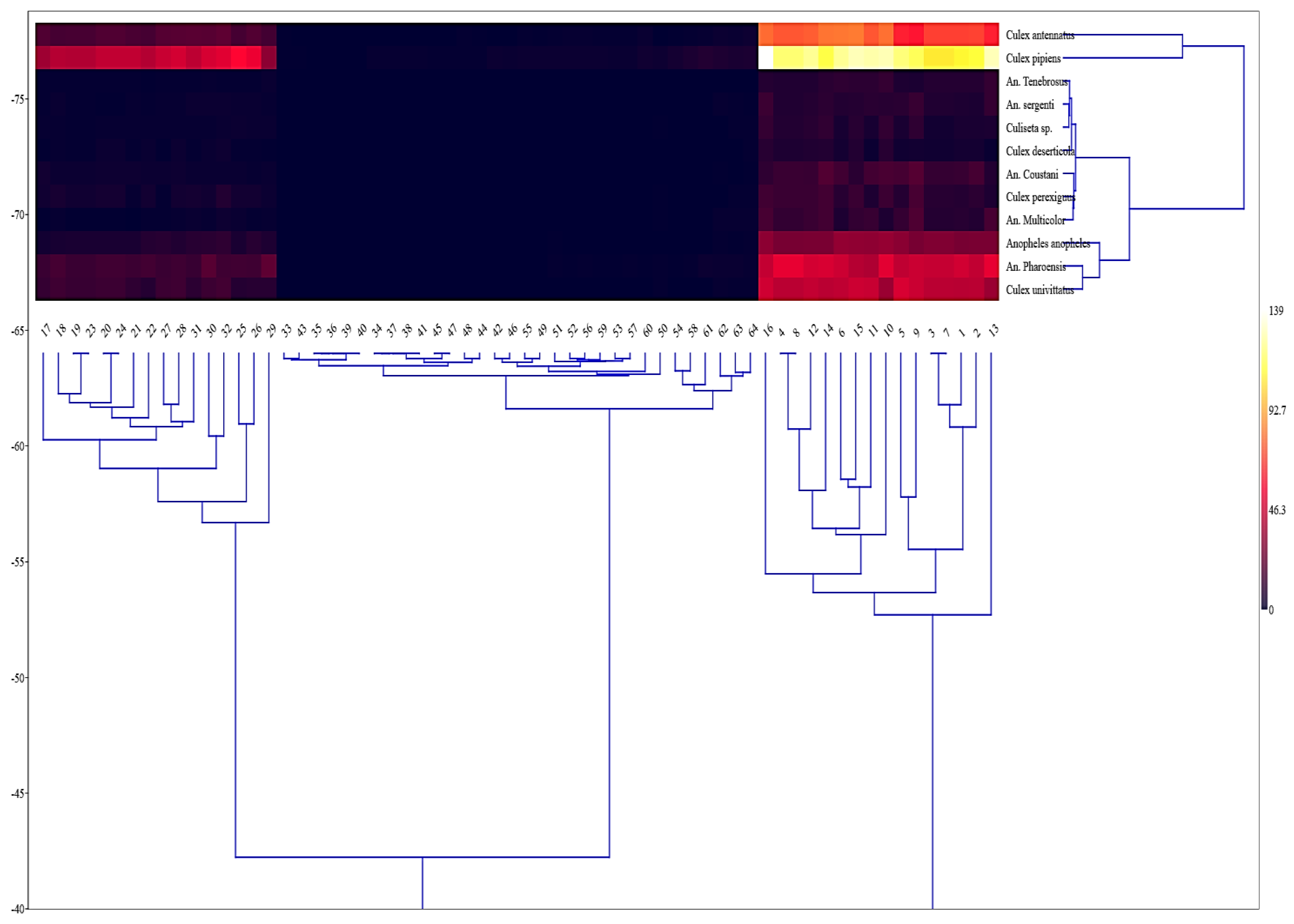
3. Results
3.1. The Spatial Distribution of Mosquito Populations
3.2. The Spatial and Temporal Patterns of Mosquito Populations
3.3. Environmental Indices
3.4. Detecting of HCV on Collecting Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Impoinvil, D.E.; Ahmad, S.; Troyo, A.; Keating, J.; Githeko, A.K.; Mbogo, C.M.; Kibe, L.; Githure, J.I.; Gad, A.M.; Hassan, A.N.; et al. Comparison of mosquito control programs in seven urban sites in Africa, the Middle East, and the Americas. Health Policy 2007, 83, 196–212. [Google Scholar] [CrossRef] [PubMed]
- Hasaballah, A.I.; Selim, T.A.; Tanani, M.A.; Nasr, E.E. Lethality and vitality efficiency of different extracts of Salix safsaf leaves against the house fly, Musca domestica L.(Diptera: Muscidae). Afr. Entomol. 2021, 29, 479–490. [Google Scholar] [CrossRef]
- Hasaballah, A.I.; El-Naggar, H.A.; Abdelbary, S.; Bashar, M.A.; Selim, T.A. Eco-friendly Synthesis of Zinc Oxide Nanoparticles by Marine Sponge, Spongia officinalis: Antimicrobial and Insecticidal Activities Against the Mosquito Vectors, Culex pipiens and Anopheles pharoensis. BioNanoScience 2022, 12, 89–104. [Google Scholar] [CrossRef]
- Selim, T.A.; Abd-El Rahman, I.E.; Mahran, H.A.; Adam, H.A.M.; Imieje, V.; Zaki, A.A.; Bashar, M.A.E.; Hwihy, H.; Hamed, A.; Elhenawy, A.A.; et al. Mosquitocidal Activity of the Methanolic Extract of Annickia chlorantha and Its Isolated Compounds against Culex pipiens, and Their Impact on the Non-Target Organism Zebrafish, Danio rerio. Insects 2022, 13, 676. [Google Scholar] [CrossRef]
- Hashem, A.H.; Selim, T.A.; Alruhaili, M.H.; Selim, S.; Alkhalifah, D.H.M.; Al Jaouni, S.K.; Salem, S.S. Unveiling antimicrobial and insecticidal activities of biosynthesized selenium nanoparticles using prickly pear peel waste. J. Funct. Biomater. 2022, 13, 112. [Google Scholar] [CrossRef]
- Abdallah, F.I.; Rady, M.H.; Merdan, B.A.; Shaarawi, F.A.; Mohammed, A.F.; Alshammery, K.A.; Al-Khalaf, A.A.; Selim, T.A.; Dahab, A.A. Effects of blood sources and artificial blood feeding membranes on the biological parameters and hepatitis C virus infectivity of Culex pipiens (Diptera: Culicidae). Afr. Entomol. 2021, 29, 262–273. [Google Scholar] [CrossRef]
- Gabarty, A.; Selim, T.A.; Hassaballah, A.I. Effect of gamma irradiation on protease and nuclease enzymes activity and egg oviposition of Culex pipiens mosquito engorged with Hepatitis C Virus (HCV). J. Radiat. Res. Appl. Sci. 2022, 15, 1–6. [Google Scholar] [CrossRef]
- Abdallah, F.I.; Merdan, B.A.; Shaarawi, F.A.; Mohamed, A.F.; Selim, T.A.; Dahesh, S.M.; Rady, M.H. The potentiality of Culex pipiens (Diptera: Culicidae) complex holobiont in transmitting the hepatitis C virus (HCV) with the aid of bacterial microbiota in the midgut. Beni-Suef Univ. J. Basic Appl. Sci. 2024, 13, 119. [Google Scholar] [CrossRef]
- Selim, T.A.; Hammad, K. Distribution of mosquitoes along Wadi El-Rayan protected area. J. Nucl. Technol. Appl. Sci. 2019, 7, 237–248. [Google Scholar] [CrossRef]
- Fawzy, M.; Helmy, Y.A. The one health approach is necessary for the control of Rift Valley fever infections in Egypt: A comprehensive review. Viruses 2019, 11, 139. [Google Scholar] [CrossRef]
- Hassan, M.I.; Mohammed, A.F.; Hammad, K.M.; Gabarty, A.; Selim, T.A. Experimental evaluation of the role of symbiotic and aposymbiotic, culex pipiens mosquitoes in the transmission of hepatitis C virus (HCV). J. Egypt. Soc. Parasitol. 2018, 48, 417–424. [Google Scholar] [CrossRef]
- Bellini, R.; Casali, B.; Carrieri, M.; Zambonelli, C.; Rivasi, P.; Rivasi, F. Aedes albopictus (Diptera: Culicidae) is incompetent as a vector of hepatitis C virus. APMIS 1997, 105, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Memon, M.I.; Memon, M.A. Hepatitis C: An epidemiological review. J. Viral Hepat. 2002, 9, 84–100. [Google Scholar] [CrossRef] [PubMed]
- Ragab, S.H.; Alqurashi, S.I.; Aljameeli, M.M.; Tyshenko, M.G.; Abdelwahab, A.H.; Selim, T.A. Predicting the Global Distribution of Gryllus bimaculatus Under Climate Change: Implications for Biodiversity and Animal Feed Production. Sustainability 2024, 16, 10278. [Google Scholar] [CrossRef]
- Ragab, S.H.; Alkhaibari, A.M.; Alharbi, J.; Areshi, S.M.; Mashlawi, A.M.; Embaby, D.M.; Tyshenko, M.G.; Selim, T.A.; Kamel, M. Impact of Climate Change on Culex pipiens Mosquito Distribution in the United States. Sustainability 2024, 17, 102. [Google Scholar] [CrossRef]
- Linthicum, K.J.; Britch, S.C.; Anyamba, A. Rift Valley Fever: An Emerging Mosquito-Borne Disease. Annu. Rev. Entomol. 2016, 61, 395–415. [Google Scholar] [CrossRef]
- Johnson, T.; Braack, L.; Guarido, M.; Venter, M.; Gouveia Almeida, A.P. Mosquito Community Composition and Abundance at Contrasting Sites in Northern South Africa, 2014–2017. J. Vector Ecol. 2020, 45, 104–117. [Google Scholar] [CrossRef]
- Rodrigue Simonet, P.N.; Alexandre Michel, N.-N.; Abel, W.; Albert, E.; Martin Hermann, G.; Franziska, S. Diversity and Abundance of Potential Vectors of Rift Valley Fever Virus in the North Region of Cameroon. Insects 2020, 11, 814. [Google Scholar] [CrossRef]
- Gillies, M.T.; Coetzee, M. A supplement to the Anopheles of Arica South of the Sahara (Afrotropical region). Publ. South Afr. Inst. Med. Res. 1987, 55, 1–143. [Google Scholar]
- Harbach, R.E. The mosquitoes of the subgenus Culex in the southwestern Asia and Egypt (Diptera: Culicidae). Contrib. Am. Entomol. Inst. 1988, 24, 1–240. [Google Scholar]
- Azari-Hamidian, S.; Harbach, R.E. Keys to the adult females and fourth-instar larvae of the mosquitoes of Iran (Diptera: Culicidae). Zootaxa 2009, 2078, 1–33. [Google Scholar] [CrossRef]
- El-Bashier, Z.M.; Hassan, M.I.; Mangoud, A.M.; Morsy, T.A.; Mohammad, K.A. A preliminary pilot survey (Culex pipiens), Sharkia governorate, Egypt. J. Egypt. Soc. Parasitol. 2006, 36, 81–92. [Google Scholar] [PubMed]
- Muturi, E.J.; Shililu, J.; Jacob, B.; Gu, W.; Githure, J.; Novak, R. Mosquito species diversity and abundance in relation to land use in a Riceland agroecosystem in Mwea, Kenya. J. Vector Ecol. 2006, 31, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Ward, W.; Hassan, M.I.; Shehata, A.Z. Spatial Distribution and Relative Abundance of Some Mosquito Species Transmitted Diseases in Beheira Governorate, Egypt. Egypt. J. Hosp. Med. 2022, 89, 7775–7785. [Google Scholar] [CrossRef]
- El-Sheikh, T.M.; Hammad, K.M.; Moselhi, W.A. Mosquito species diversity and abundance in relation to rice land agroecosystem and filarial infection in Kafr El-Sheikh Governorate, Egypt. Egypt. J. Hosp. Med. 2010, 38, 100–114. [Google Scholar] [CrossRef]
- Rifaat, M.A.; Mahdi, A.H.; Wassif, S.F. Some ecological studies on Aedes (Ochlerotatus) caspius in the Nile Delta. J. Egypt. Public Health Assoc. 1970, 45, 451–457. [Google Scholar]
- Mohamed, N.H.; Salem, S.A.; Abdel Baki, M.H.; Fawzy, A.F.A. Types of mosquitoes in Giza governorate in reference to filarial. J. Egypt. Soc. Parasitol. 1981, 28, 449–459. [Google Scholar]
- Ragab, S.H.; Khaled, M.A.; Taha, R.H.; El-Tabakh, M. Spatial Distribution of Appropriate Aquatic Mosquitos’ Larval Sites Occurrence Using Integration of Field Data and GIS Techniques. Egypt. J. Aquat. Biol. Fish. 2023, 27, 355–371. [Google Scholar] [CrossRef]
- El-Shazly, A.M.; Ali, M.E.; Handoussa, A.E.; Abdalla, K.F. Studies on culicini larvae in Mansoura center, Dakahlia governorate, Egypt. J. Egypt. Soc. Parasitol. 1998, 28, 839–847. [Google Scholar]
- Mostafa, A.A.; Allam, K.A.M.; Osman, M.Z. Mosquito species and their densities in some Egyptian governorate. J. Egypt. Soc. Parasitol. 2002, 32, 9–20. [Google Scholar]
- Kirkpatrick, T.W. The Mosquitoes of Egypt; The Egyptian Government Press: Cairo, Egypt, 1925.
- Salem, H.M. New records of some Egyptian mosquitoes. Bull. Soc. Entomol. d′Égypte 1933, 17, 83–90. [Google Scholar]
- Wassif, S.F. Survey of the Egyptian culicines in the Nile Delta with Special Reference to Filarial Transmission. Master’s Thesis, Faculty of Medicine, Ain Shams University, Cairo, Egypt, 1969. [Google Scholar]
- Hurlbut, H.S.; Weitz, B. Some observation on the bionomics of the common mosquitoes of the Nile Delta. Am. J. Trop. Med. Hyg. 1956, 5, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Gad, A.M. Mosquitoes of the Oases of the Libyan Desert of Egypt. Bull. Soc. Entomol. d′Égypte 1956, 40, 131–139. [Google Scholar]
- Morsy, T.A.; El Okbi, L.M.A.; Kamal, A.M.; Ahmed, M.M.; Bo-shra, E.F. Mosquitoes of the genus Culex in the Suez Canal governorate, Egypt. J. Egypt. Soc. Parasitol. 1990, 11, 441–451. [Google Scholar]
- Mwangangi, J.M.; Muturi, E.J.; Mbogo, C.M. Seasonal mosquito larval abundance andcomposition in Kibwezi, lower eastern Kenya. J. Vector Borne Dis. 2009, 46, 65–71. [Google Scholar]
- Kenawy, M.A.; El-Said, S. Characterization of Culicine mosquito habitats in the Nile Delta, Egypt. In Proceedings of the 14th International Conference Statistics, Computer Science, Social & Demographic Research, Cairo, Egypt, 25–30 March 1989; Volume 1, pp. 211–231. [Google Scholar]
- Ijumba, J.; Mosha, F.; Lindsay, S.W. Malaria transmission risk variations derived from different agricultural practices in an irrigated area of northern Tanzania. Med. Vet. Entomol. 2002, 16, 28–38. [Google Scholar] [CrossRef]
- Shililu, J.; Tewolde, G.; Fessahaye, S.; Mengistu, S.; Fekadu, H.; Mehari, Z.; Asmelash, G.; Sintasath, D.; Bretas, G.; Mbogo, C.; et al. Larval habitat diversity and ecology of anopheline larvae in Eritrea. J. Med. Entomol. 2003, 40, 921–929. [Google Scholar] [CrossRef]
- Silverman, A.L.; McCruy, D.G.; Gordon, S.C.; Morgan, W.T.; Walker, E.D. Experimental evidence against replication or dissemination of hepatitis C virus in mosquitoes (Diptera: Culicidae) using detection by reverse transcriptase polymerase chain reaction. J. Med. Entomol. 1996, 33, 398–401. [Google Scholar] [CrossRef]
- Chang, T.T.; Chang, T.Y.; Chen, C.C.; Young, K.C.; Roan, J.N.; Lee, Y.C.; Cheng, P.N.; Wu, H.L. Existence of hepatitis C virus in Culex quinquefasciatus after ingestion of infected blood: Experimental approach to evaluating transmission by mosquitoes. J. Clin. Microbiol. 2001, 39, 3353–3355. [Google Scholar] [CrossRef]
- Tarish, H.R.; Al-Mola, G.A.; Abdulsada, K.M.; Lateef, R.H.; Auda, G.R. Role of Culex mosquito in the transmission of hepatitis C virus: An experimental study in Iraq. Am. J. Res. Commun. 2014, 2, 176–185. [Google Scholar]
- Yeom, J.S. Current status and outlook of mosquito-borne diseases in Korea. J. Korean Med. Assoc. 2017, 60, 468–474. [Google Scholar] [CrossRef]
- Hamer, G.L.; Kitron, U.D.; Brawn, J.D.; Loss, S.R.; Ruiz, M.O.; Goldberg, T.L.; Walker, E.D. Culex pipiens (Diptera: Culicidae): A bridge vector of West Nile virus to humans. J. Med. Entomol. 2008, 45, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Hasaballah, A.I.; El-Naggar, H.A. Antimicrobial activities of some marine sponges, and its biological, repellent effects against Culex pipiens (Diptera: Culicidae). Annu. Res. Rev. Biol. 2017, 12, 1–14. [Google Scholar] [CrossRef]
- Shehabeldine, A.M.; Elbahnasawy, M.A.; Hasaballah, A.I. Green Phytosynthesis of Silver Nanoparticles Using Echinochloa stagnina Extract with Reference to their Antibacterial, Cytotoxic, and Larvicidal Activities. BioNanoScience 2021, 11, 526–538. [Google Scholar] [CrossRef]
- Chatterjee, S.; Kim, C.M.; Yun, N.R.; Kim, D.M.; Song, H.J.; Chung, K.A. Molecular detection and identification of Culex flavivirus in mosquito species from Jeju, Republic of Korea. Virol. J. 2021, 18, 150. [Google Scholar] [CrossRef]
- Bryant, J.E.; Crabtree, M.B.; Nam, V.S.; Yen, N.T.; Duc, H.M.; Miller, B.R. Isolation of arboviruses from mosquitoes collected in northern Vietnam. Am. J. Trop. Med. Hyg. 2005, 73, 470–473. [Google Scholar] [CrossRef]
- Ochieng, C.; Lutomiah, J.; Makio, A.; Koka, H.; Chepkorir, E.; Yalwala, S.; Mutisya, J.; Musila, L.; Khamadi, S.; Richardson, J.; et al. Mosquito-borne arbovirus surveillance at selected sites in diverse ecological zones of Kenya; 2007–2012. Virol. J. 2013, 10, 140. [Google Scholar] [CrossRef]
- de Morais Bronzoni, R.V.; Baleotti, F.G.; Ribeiro Nogueira, R.M.; Nunes, M.; Moraes Figueiredo, L.T. Duplex reverse transcription-PCR followed by nested PCR assays for detection and identification of Brazilian alphaviruses and flaviviruses. J. Clin. Microbiol. 2005, 43, 696–702. [Google Scholar] [CrossRef]
- Young, Y.B.; Seo, H.P.; Myung-Deok, K.-J. Monitoring Culicine Mosquitoes (Diptera: Culicidae) as a Vector of Flavivirus in Incheon Metropolitan City and Hwaseong-Si, Gyeonggi-Do, Korea, during 2019. Korean J. Parasitol. 2020, 58, 551–558. [Google Scholar]
- Alhaj, M.; Aljidhean, G.; Almanae, A.A.; Abdalla, M.; Muhanna, H. Abundance and Spatial Distribution of Culex and Aedes Mosquitoes as Potential Vectors of Rift Valley Fever in Jazan Region. Int. J. Mosq. Res. 2017, 4, 42–47. [Google Scholar]
- Mweya, C.N.; Kimera, S.I.; Kija, J.B.; Mboera, L.E. Predicting Distribution of Aedes Aegypti and Culex Pipiens Complex, Potential Vectors of Rift Valley Fever Virus in Relation to Disease Epidemics in East Africa. Infect. Ecol. Epidemiol. 2013, 3, 21748. [Google Scholar]
- Abdel-Hamid, Y.M.; Soliman, M.I.; Kenawy, M.A. Population ecology of mosquitoes and the status of bancroftian filariasis in El Dakahlia Governorate, the Nile Delta, Egypt. J. Egypt. Soc. Parasitol. 2013, 43, 103–113. [Google Scholar] [PubMed]
- El-Bahnasawy, M.M.; Abdel Fadil, E.E.; Morsy, T.A. Mosquito vectors of infectious diseases: Are they neglected health disaster in Egypt? J. Egypt. Soc. Parasitol. 2013, 43, 373–386. [Google Scholar] [PubMed]
- El-naggar, A.; Elbanna, S.M.; Abo-ghalia, A. The impact of some environmental factors on the abundance of mosquitoes larvae in certain localities of Sharkia Governorate in Egypt. Egypt. Acad. J. Biol. Sci. A Entomol. 2013, 6, 49–60. [Google Scholar] [CrossRef]
- World Health Organization. Guidance for Country Validation of Viral Hepatitis Elimination and Path to Elimination: Technical Report; World Health Organization: Geneva, Switzerland, 2023.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Volume (µL) |
|---|---|
| oasig OneStep2X RT-qPCR master mix | 10 |
| HCV primer/probe mix | 1 |
| RNA template | 5 |
| DNA/RNA-free water | 4 |
| Final volume | 20 |
| Kafr Saqr | Abo Kebeer | Diarb Negm | El-Zakazik | ||
|---|---|---|---|---|---|
| Summer | Individuals | 349.75 ± 6.815 | 359.75 ± 8.913 | 380.93 ± 6.418 | 392.06 ± 10.838 |
| Dominance_D | 0.18 ± 0.001 | 0.18 ± 0.002 | 0.17 ± 0.005 | 0.18 ± 0.006 | |
| Simpson_1-D | 0.81 ± 0.001 | 0.81 ± 0.002 | 0.82 ± 0.005 | 0.81 ± 0.006 | |
| Shannon_H | 2 ± 0.008 | 1.99 ± 0.012 | 2.03 ± 0.025 | 2.03 ± 0.026 | |
| Evenness_e^H/S | 0.61 ± 0.005 | 0.61 ± 0.007 | 0.63 ± 0.016 | 0.63 ± 0.016 | |
| Brillouin | 1.91 ± 0.009 | 1.91 ± 0.012 | 1.95 ± 0.025 | 1.95 ± 0.026 | |
| Menhinick | 0.64 ± 0.006 | 0.63 ± 0.007 | 0.61 ± 0.005 | 0.6 ± 0.008 | |
| Margalef | 1.87 ± 0.006 | 1.87 ± 0.007 | 1.85 ± 0.005 | 1.84 ± 0.008 | |
| Equitability_J | 0.8 ± 0.003 | 0.8 ± 0.005 | 0.81 ± 0.01 | 0.81 ± 0.01 | |
| Fisher_alpha | 2.4 ± 0.011 | 2.39 ± 0.014 | 2.35 ± 0.009 | 2.34 ± 0.015 | |
| Berger-Parker | 0.31 ± 0.002 | 0.32 ± 0.003 | 0.32 ± 0.005 | 0.32 ± 0.012 | |
| Autumn | Individuals | 107.18 ± 4.358 | 104.18 ± 3.402 | 112.12 ± 2.175 | 113.93 ± 9.445 |
| Dominance_D | 0.22 ± 0.002 | 0.23 ± 0.009 | 0.24 ± 0.014 | 0.2 ± 0.004 | |
| Simpson_1-D | 0.77 ± 0.002 | 0.76 ± 0.009 | 0.75 ± 0.014 | 0.79 ± 0.004 | |
| Shannon_H | 1.8 ± 0.006 | 1.77 ± 0.033 | 1.77 ± 0.03 | 1.91 ± 0.024 | |
| Evenness_e^H/S | 0.59 ± 0.018 | 0.58 ± 0.012 | 0.54 ± 0.013 | 0.58 ± 0.013 | |
| Brillouin | 1.61 ± 0.008 | 1.58 ± 0.028 | 1.59 ± 0.028 | 1.7 ± 0.023 | |
| Menhinick | 1 ± 0.028 | 1.01 ± 0.03 | 1.03 ± 0.02 | 1.1 ± 0.036 | |
| Margalef | 1.99 ± 0.07 | 1.99 ± 0.045 | 2.1 ± 0.032 | 2.25 ± 0.026 | |
| Equitability_J | 0.77 ± 0.01 | 0.76 ± 0.01 | 0.74 ± 0.01 | 0.77 ± 0.009 | |
| Fisher_alpha | 2.84 ± 0.118 | 2.84 ± 0.09 | 3.01 ± 0.064 | 3.27 ± 0.072 | |
| Berger-Parker | 0.38 ± 0.011 | 0.4 ± 0.012 | 0.44 ± 0.021 | 0.37 ± 0.008 | |
| Winter | Individuals | 1.91 ± 0.315 | 1.68 ± 0.236 | 1.87 ± 0.484 | 1.81 ± 0.199 |
| Dominance_D | --- | --- | --- | 0.66 ± 0.152 | |
| Simpson_1-D | --- | --- | --- | 0.33 ± 0.152 | |
| Shannon_H | 0.16 ± 0.103 | 0.05 ± 0.058 | 0.16 ± 0.056 | 0.22 ± 0.087 | |
| Evenness_e^H/S | 1.01 ± 0.017 | 1.01 ± 0.017 | 1.04 ± 0.018 | 1.03 ± 0.023 | |
| Brillouin | 0.08 ± 0.05 | 0.02 ± 0.021 | 0.07 ± 0.024 | 0.09 ± 0.037 | |
| Menhinick | 0.91 ± 0.078 | 0.85 ± 0.071 | 0.91 ± 0.078 | 0.96 ± 0.053 | |
| Margalef | 0.18 ± 0.115 | 0.09 ± 0.09 | 0.21 ± 0.087 | 0.27 ± 0.115 | |
| Equitability_J | --- | --- | --- | --- | |
| Fisher_alpha | 0.91 ± 0.431 | 0.4 ± 0.162 | 0.34 ± 0.164 | 0.65 ± 0.25 | |
| Berger-Parker | 0.93 ± 0.039 | 0.96 ± 0.031 | 0.91 ± 0.029 | 0.9 ± 0.041 | |
| Spring | Individuals | 5.37 ± 0.88 | 5.04 ± 0.715 | 5.75 ± 0.94 | 16.12 ± 1.252 |
| Domice_D | 0.38 ± 0.089 | 0.47 ± 0.076 | 0.36 ± 0.078 | 0.25 ± 0.081 | |
| Simpson_1-D | 0.61 ± 0.089 | 0.52 ± 0.076 | 0.63 ± 0.078 | 0.74 ± 0.081 | |
| Shannon_H | 0.97 ± 0.25 | 0.78 ± 0.108 | 0.89 ± 0.098 | 1.69 ± 0.279 | |
| Evenness_e^H/S | 1.06 ± 0.023 | 1.02 ± 0.053 | 1.04 ± 0.051 | 0.92 ± 0.039 | |
| Brillouin | 0.53 ± 0.145 | 0.42 ± 0.06 | 0.49 ± 0.053 | 1.15 ± 0.185 | |
| Menhinick | 1.21 ± 0.165 | 1.03 ± 0.075 | 1.1 ± 0.086 | 1.61 ± 0.273 | |
| Margalef | 1.02 ± 0.289 | 0.79 ± 0.108 | 0.89 ± 0.098 | 1.97 ± 0.421 | |
| Equitability_J | 1.05 ± 0.011 | 1 ± 0.069 | 1.03 ± 0.065 | 0.93 ± 0.044 | |
| Fisher_alpha | 3.16 ± 1.061 | 2.11 ± 0.42 | 2.38 ± 0.492 | 4.75 ± 1.358 | |
| Berger-Parker | 0.62 ± 0.087 | 0.69 ± 0.053 | 0.62 ± 0.06 | 0.44 ± 0.072 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selim, T.A.; Ragab, S.H.; Riad, S.A.; Eltaly, R.I.; Mohammed, S.H.; Sharawi, S.E.; Alkenani, N.A.; Almahallawi, R.S.; Al-Rashidi, H.S.; El-Tabakh, M.A.M. Abundance, Diversity and Distribution of Mosquito Species and Molecular Detection of Its Associated Hepatitis C Virus in Sharkia Governorate, Egypt. Insects 2025, 16, 433. https://doi.org/10.3390/insects16040433
Selim TA, Ragab SH, Riad SA, Eltaly RI, Mohammed SH, Sharawi SE, Alkenani NA, Almahallawi RS, Al-Rashidi HS, El-Tabakh MAM. Abundance, Diversity and Distribution of Mosquito Species and Molecular Detection of Its Associated Hepatitis C Virus in Sharkia Governorate, Egypt. Insects. 2025; 16(4):433. https://doi.org/10.3390/insects16040433
Chicago/Turabian StyleSelim, Tharwat A., Sanad H. Ragab, Saber A. Riad, Randa I. Eltaly, Shaimaa H. Mohammed, Somia Eissa Sharawi, Naser Ahmed Alkenani, Ruoa Saleh Almahallawi, Hayat S. Al-Rashidi, and Mohamed A. M. El-Tabakh. 2025. "Abundance, Diversity and Distribution of Mosquito Species and Molecular Detection of Its Associated Hepatitis C Virus in Sharkia Governorate, Egypt" Insects 16, no. 4: 433. https://doi.org/10.3390/insects16040433
APA StyleSelim, T. A., Ragab, S. H., Riad, S. A., Eltaly, R. I., Mohammed, S. H., Sharawi, S. E., Alkenani, N. A., Almahallawi, R. S., Al-Rashidi, H. S., & El-Tabakh, M. A. M. (2025). Abundance, Diversity and Distribution of Mosquito Species and Molecular Detection of Its Associated Hepatitis C Virus in Sharkia Governorate, Egypt. Insects, 16(4), 433. https://doi.org/10.3390/insects16040433