Simple Summary
The Danube Clouded Yellow (Colias myrmidone) is a species that is under special conservation concern in Europe. The reasons for its quick and dramatic decline and local extirpations are not fully understood; genetic data are vital to understanding these reasons. We studied the last two neighboring populations from Central Europe—one that was recently extirpated and another that still exists—using a mitochondrial barcoding marker and a nuclear gene. We also tested samples for the endosymbiotic bacterium Wolbachia, which is known to induce reproductive manipulations in their hosts, affecting mtDNA diversity among populations. The results of our research suggest that a relatively high genetic polymorphism has decreased in recent years. Restricted gene flow that is related to habitat fragmentation and isolation has led to the quick differentiation of populations; however, these populations probably used to be genetically uniform, at least at the regional scale. DNA barcode-based analysis revealed the presence of two haplogroups, but, interestingly, only individuals from one of them were infected by Wolbachia. This phenomenon turned out to be female-biased, which has a complex impact on the host. Finally, a comparison of our data with other available data indicated significant barcode sharing between C. myrmidone and its sibling C. caucasica, which allows us to question the distinctiveness of both vulnerable taxa.
Abstract
The Danube Clouded Yellow (Colias myrmidone) has experienced one of the most dramatic declines among European butterflies. To estimate genetic diversity in the last population in Poland that has survived in the Knyszyn Forest (KF), we analyzed mitochondrial (COI) and nuclear (EF-1α) polymorphisms in individuals sampled in 2014 and 2022. The results were compared with genetic data obtained in 2014 from a recently extirpated nearby population (Czerwony Bór, CB). Because mtDNA polymorphisms in insects can be modulated by endosymbionts, the samples were screened for Wolbachia. The polymorphism of EF-1α indicated that diversity was gradually decreasing. The KF experienced rapid demographic processes, manifested by a significant change in allele frequency. The small differentiation in nuclear markers between the KF and CB in 2014 suggests that the regional population used to be genetically uniform. Four COI haplotypes that were identified in this study probably belong to two different haplogroups. Wolbachia was detected only in individuals with one specific haplotype, and the prevalence was female-biased, suggesting the induction of two reproductive manipulations. The most common COI haplotype found in Poland was the same as that reported from other parts of Europe, not only for C. myrmidone but also C. caucasica. These results allow us to question the distinctiveness of each taxa.
1. Introduction
The Danube Clouded Yellow Colias myrmidone is an example of one of the most dramatic declines in population among all European butterflies. The beginning of the century coincided with the process of the rapid extinction of the species. Van Swaay and Warren [1] evaluated its status as “Vulnerable” at the continental scale; however, only 20 years later, the weighted Red List value index in Europe put it among the top five most endangered species [2]. The butterfly is critically endangered in the European Union [3], being considered as a species of conservation concern due to its presence in annexes of the Habitats Directive. It was extirpated in several EU countries and has recently been confirmed to now only be found in Poland [4] and Romania [5].
The need to protect this species was first highlighted in 1990 in Bayern (Germany) [6]. The causes of the species decrease are complex and not fully understood. Some habitats deteriorated or ceased to exist as a consequence of agricultural intensification or due to the abandonment of grazing, as well as afforestation and changes in woodland management. Changes in water balance and the system of habitats have also been reported [4,7,8,9,10,11]. However, the butterfly has also disappeared from biotopes, i.e., with a large presence of larval food plants. The last populations have survived in fragmented landscapes, and recent studies have indicated that C. myrmidone requires a relatively large space and well-connected habitat patches [4]. Moreover, it is considered to be a high-risk species in the context of climate change [12], so some sudden local disappearances can be explained by unfavorable seasonal weather conditions [4].
Spatially isolated populations are particularly vulnerable to the effects of various factors that can cause extirpation [13]. Although the role of genetic factors in the extinction of populations and species is still a controversial issue and a hotly debated topic [14,15,16], there is no doubt that in isolated populations, genetic variability is reduced as a result of genetic drift and the increasing relatedness of individuals [17]. In turn, it is known that genetic variability is crucial for maintaining the adaptive potential of populations and species in the face of changing environmental conditions [18]. There are many examples of narrowed genetic diversity in butterfly populations threatened with extinction [19]. Low diversity and high differentiation are typical of sedentary specialists. On the other hand, it has been considered that intermediate species between generalists and specialists (like C. myrmidone) can suffer even more from genetic impoverishment when the gene pool cannot be refreshed due to a lack of connectivity [20]. It is also known that changes in the level of genetic diversity can be a valuable clue when identifying populations that should be classified as endangered [21]; this approach is applicable in the case of butterflies [22]. Therefore, estimating the level of genetic variability and tracking its changes may prove crucial in planning conservation strategies for endangered species [23,24].
Being a large and spectacular butterfly, C. myrmidone remains, in some respects, an insufficiently studied species. Taking into account the genetic aspect, there exist taxonomic studies of the genus Colias or higher taxonomic units based on analyses of single individuals from some species within the genus. In particular, genomic studies have provided knowledge on the evolution of Colias [25]. However, population studies of C. myrmidone have not been conducted to date. Therefore, there are significant gaps in the knowledge regarding, for example, genetic structure. Understanding genetic differentiation, diversity, and other ongoing processes is vital from a conservation point of view; without such information, it is hard to imagine, for example, possible reintroduction efforts [26]. The majority of available data on C. myrmidone are in relation to mtDNA, namely the cytochrome c oxidase subunit I (COI), a section of which has become the standard DNA barcode for animals [27] and has been added to barcoding libraries [28,29].
The aim of the present study was to obtain insights into the last regional population inhabiting the Knyszyn Forest in Northeastern Poland. Over the last thirty years, the species has disappeared from the entire country, except for in this isolated area [4]; the next nearest known localities are in Southern Belarus [30]. Hence, we carried out extensive sampling in the Knyszyn Forest and obtained some material from the recently extirpated penultimate population in the same region. We analyzed COI and the nuclear gene elongation factor 1α (EF-1α), which is commonly used in phylogenetically oriented butterfly surveys to complement mtDNA markers [31,32].
Analyses of COI sequences belonging to over 300 butterfly species in Western Europe indicate that the spatial differentiation in mtDNA is negatively correlated with species traits determining dispersal capability and colonization ability [33]. However, variation in mtDNA may not be correlated with genomic variation due to inheritance mechanisms and other factors, e.g., microbiological infections. Hence, we tested samples for the presence of α-proteobacteria Wolbachia (Rickettsiaceae), i.e., maternally inherited symbiotic bacterium of ca. two-thirds of all arthropod species [34], which can affect the reproduction of its host [35].
The consequence of segregation distortion may be the origin of different mtDNA lineages or the reduction in genetic diversity when a single haplotype (or haplogroup) dominates a population/region/range [26,36,37]. Wolbachia infections may affect the mitochondrial structure of hosts [38] and even mimic cryptic speciation [39]. In the case of species of conservation concern, this bacterium is a possible serious threat since the introduction of Wolbachia into uninfected populations or the introduction of a new strain may increase the risk of extinction. Therefore, knowledge about the pattern of infection is vital in the context of actions that incorporate captive breeding or translocation [40,41].
2. Materials and Methods
2.1. Study Species
The Danube Clouded Yellow Colias myrmidone (Esper, 1781) is a relatively large pierid butterfly (Lepidoptera, Pieridae), with a wingspan of about 40–46 mm. Sexual dimorphism is distinct, with two main female forms, i.e., gynomorphic orange and andromorphic pale yellow. Two or three generations are on the wing every year, depending on season and locality. A variety of flowers are used as nectar resources. Males are patrollers; they move in fast flight looking for mates. Females lay eggs singly on the upper side of the leaves of several Chamaecytisus species. Caterpillars initially leave window-like traces of feeding; later, they eat whole leaves. Pupation takes place on food plants or vegetation close to the ground (Figure 1). The medium-sized larvae of the first generation overwinter among fallen leaves of the food plant [4].

Figure 1.
Colias myrmidone and its habitat in the Knyszyn Forest: (a) a mating pair with a gynomorphic (alba) female (below), (b) an ovipositing andromorphic (orange) female on the only local larval food plant, i.e., Chamaecytisus ruthenicus, (c) an egg, (d) a young caterpillar, (e) a full-grown caterpillar, (f) a chrysalis, (g) a typical patch of habitat, i.e., recently replanted clear-cut, (h) a flowering Chamaecytisus ruthenicus plant in the spring; © Marcin Sielezniew.
The distribution range of C. myrmidone is almost entirely limited to Europe. A dramatic decline has been observed across the continent with extinctions in Germany, Czech Republic, Hungary, Austria, Slovenia, and Lithuania, and the butterfly has been reported recently only from Poland, Romania, Belarus, Ukraine, and Russia [9]. In Balkans, C. myrmidone is replaced by a sibling taxon Colias caucasica balcanica Rebel, 1901, whose status is, however, still disputable [29].
A severe contraction of its range has been observed in Poland, where the species at present occurs only in the Knyszyn Forest in the northeastern part of the country (Figure 2) [4,42]. The last metapopulation depends on forestry, and almost all its habitats are recent clear-cuts. The only larval food plant available, i.e., Chaemocytisus ruthenicus, is present in some tree stands, and after logging, it typically grows lush and covers the ground extensively in some patches due to increased sun exposure. Therefore, clear-cuts may be colonized by C. myrmidone and support local populations for several years until an increase in shading is caused by growth of replanted trees or natural succession [4]. The penultimate locality was the former military area (Czerwony Bór) situated ca. 100 km away (Figure 2), where C. myrmidone was extirpated around 2019, although the habitat looked still suitable [4].
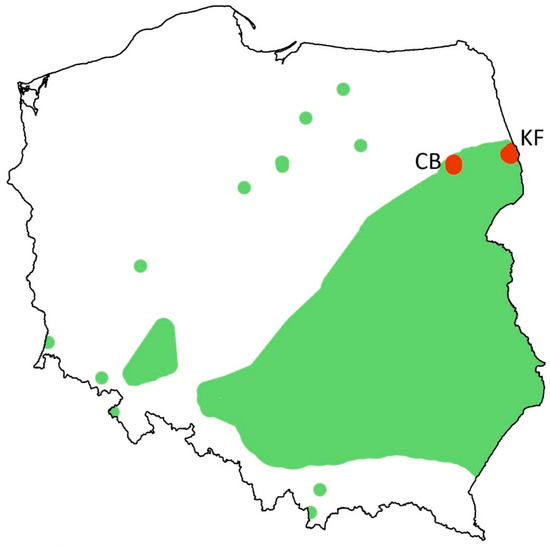
Figure 2.
Map of Poland, showing sampling sites (red dots) at the background of the former distribution range (green patches) and isolated sightings (green dots) of Colias myrmidone. KF—the last population in the Knyszyn Forest, CB—Czerwony Bór, where the butterfly was extirpated around 2019 (based on [42], modified).
2.2. Sampling for Genetic Studies
The main sampling was performed in 2022 in the Knyszyn Forest along intensive mark–release–recapture studies (see [4]). A total number of 125 individuals was sampled, including 53 females (28 andromorphic and 25 gynomorphic) and 73 males. Sex was recorded for all individuals, and for females, color forms were also recorded (Table S1). Additionally, we used samples collected in the Knyszyn Forest (12) and in Czerwony Bór (21) in 2014, but details concerning sex were not recorded at that time. All tissue samples were taken using a nonlethal method. Small fragments of hind wings (~2 mm2) or middle legs were torn off using tweezers and then individually stored in 95% ethanol. Pierid butterflies (contrasting to nymphalids) use all six legs for walking, so such damage is insignificant. No approval of research ethics committees was required for such procedure in Poland since sampling was conducted with unregulated invertebrate species. Here, it is worth mentioning that sampled individuals were often recorded again during mark–release–recapture studies [4]. The lack of negative effects of single-leg removal on the survival, behavior, and mobility of individuals has also been confirmed for other butterfly species [43].
2.3. Laboratory Procedures
DNA was extracted from a single leg or a single piece of wing. In the case of fresh samples (collection 2022), the GeneMATRIX Bio-Trace DNA Purification Kit was used, whereas for older sample specimens (2014), we used the GeneMAGNET Human and Animal Tissue DNA Purification Kit (EURx, Gdańsk, Poland). The fragment of wing or leg was cut into pieces in a sterile Petri dish using a sterile scalpel. The tissue fragments were placed in tubes with lysing buffer with Proteinase K and incubated overnight at 56 °C. Then, extraction of DNA was performed according to a standard protocol. DNA extracts were stored at −20 °C for further procedures.
To estimate genetic diversity, we analyze fragments of mitochondrial COI (Cytochrome Oxidase Subunit 1) and nuclear EF-1α (Elongation Factor-1α) genes. We aimed to amplify and sequence ~658-base pair (bp) fragment of COI and ~650 bp of EF-1α. Additionally, we amplified fragments of Wsp (Wolbachia surface protein) gene (~540 bp) to detect presence of endosymbiotic Wolbachia. In the case of COI, PCR amplification was carried out using primers LCO1490 (forward) and HCO2198 (reverse) [44]. Fragments of EF-1α were amplified using primers ELF2 (forward) [45] and efrcM4 (reverse) [46]. To detect Wolbachia, we used primers wsp81F (forward) and 691R (reverse) [47], amplifying the fragment of Wsp gene.
In the case of COI and EF-1α the reaction mixture contained 2 or 3 μL of DNA extract depending on the initial concentration (ranging from 10 to 100 ng/μL), 7.5 μL or 8.5 μL of water (depending on the amount of DNA), 12.5 μL of PCR Master Mix (Promega GoTaq G2 Hot Start Green Master Mix) and 1 μL of forward and reverse primer. The amplification conditions for COI were as follows: initial denaturation for 7 min at 94 °C, 35 cycles of 1 min at 94 °C, 1 min at 54 °C, 2 min at 72 °C and final elongation for 7 min at 72 °C. For EF1α, we used the following PCR conditions: initial denaturation for 5 min at 94 °C, 40 cycles of 1 min at 94 °C, 1 min at 62 °C, 1 min 30 s at 72 °C and final elongation for 10 min. at 72 °C. Testing presence of Wolbachia, the following PCR conditions were applied: initial denaturation for 5 min at 94 °C, 40 cycles of 1 min at 94 °C, 1 min at 53 °C, 1 min at 72 °C and final elongation for 10 min at 72 °C.
Agarose gel electrophoresis was performed to visualize the PCR products. In the case of no visible bands, re-amplification was performed. Namely, as a template for PCR reaction, we used 2 μL of previous PCR final mixture. In addition, the reaction mixture contained 8.5 μL of water, 12.5 μL of PCR Master Mix (Promega GoTaq G2 Hot Start Green Master Mix), and 1 μL of forward and reverse primer. The amplified fragments were sequenced using the ABI 3500XL Genetic Analyzer. The PCR products were purified using ExoSAP-IT (Applied Biosystems, Waltham, MA, USA), following the standard protocol. Next, we performed sequencing PCR using the BigDye Terminator v3.1 cycle sequencing kit (Applied Biosystems). The reaction mixture contained 2 μL of template, 4 μL of BigDye Terminator, 2 μL of BigDye terminator sequencing buffer, 2 μL of forward or reverse primer (3.2 μM), and 10 μL of water. The last stage before sequencing was purifying PCR products using HighPrep DTR (MAGBIO) following standard protocol. The results were sequenced using the ABI 3500XL Genetic Analyzer.
2.4. Statistical Analysis
DNA sequences were aligned in BioEdit v 7.0.4 [48] and revised manually. The haplotype reconstruction of the nuclear EF-1 α gene from unphased/genotype data was conducted using the algorithms provided in PHASE as implemented in DNASP 5.0 [49].
To characterize the diversity of amplified genes, the number of haplotypes (H), haplotype diversity (h), nucleotide diversity (π), and mean number of nucleotide differences among the haplotypes (k) in the overall sample and both populations were calculated using DNAsp 5.10 [49]. ARLEQUIN v3.5.2.2 [50] was used to estimate genetic differentiation among the groups of samples (ΘST), dividing by localization (Knyszyn Forest vs. Czerwony Bór) and year of sampling (2014 vs. 2022). Permutation test (1000 repetition) was applied to estimate significance of the genetic differentiation.
To analyze whether previous fluctuations in population size had affected current genetic diversity, we used DNAsp and ARLEQUIN to perform basic neutrality tests, detecting past population expansion, Fu’s FS [51], and Tajima’s D [52]. Both tests use the infinite site model without recombination to test for departures from selective neutrality and population equilibrium for intraspecific data. Fu’s FS, Tajima’s D, and related statistics signal an excess of rare mutations when the values are negative and differ significantly from zero. Fu’s FS uses information from the haplotype distribution and has greater statistical power to detect population expansion than other available tests [51]. Low FS values indicate an excess of single substitutions, usually due to expansion.
Tajima’s D-test, in turn, compares the number of nucleotide differences between sequences and the number of differences between segregating sites. In this case, population expansion will result in a significant negative departure from zero [52].
The significance of both values was determined by way of a coalescent simulation, using 1000 replicates in DNAsp. Because Fu [51] showed that a significance of 0.02 was equivalent to a 0.05 level where Fs was concerned, the test was considered significant when p ≤ 0.02.
To identify past demographic processes that could have influenced the observed genetic diversity in C. myrmidone in Poland, a mismatch distribution analysis [53] was performed in ARLEQUIN. We compared distributions for numbers of nucleotide differences among haplotypes in sampling locations and in the whole Polish population of the species to the distribution of differences in a model population that had undergone recent expansion. In a stable population, the distribution is usually multimodal, whereas in a population following recent expansion, a unimodal distribution of differences may be anticipated. A thousand bootstrap replicates were used to generate an expected distribution using a model of sudden demographic expansion. The sum of squared deviations (SSD) between the observed and model distributions was used as a test statistic. Significant values for SSD indicate deviation from sudden expansion. Additionally, we estimated the raggedness index and its significance. This measure quantifies “smoothness” of the observed mismatch distribution. Low values for raggedness are typical for an expanding population, whereas higher values are expected in the case of stationary or bottlenecked populations [54].
For both genes, a median-joining haplotype network [55] was constructed in NETWORK v4.6.1.1. (Fluxus Technology Ltd., Suffolk, UK) and PopArt v4.8.4. (http://popart.otago.ac.nz, accessed on 12 December 2024). In the case of COI, haplotypes identified in Colias caucasica were also included (Supplementary Table S2), as the comparison of the obtained haplotypes with the sequences from GenBank showed that one of the C. myrmidone haplotypes (CmCOI-2) was identical to the haplotype found in another species―C. caucasica.
Finally, the COI haplotypes identified in this study were compared with homologous sequences for the C. myrmidone obtained from GenBank (https://www.ncbi.nlm.nih.gov/genbank, accessed on 12 December 2024) (Supplementary Table S2). To resolve the evolutionary relationship between haplotypes identified in this study and those found in other European Colias species, we constructed a phylogenetic tree using the neighbour-joining method, as based on Tamura-3 model with gamma distribution (T92+G). The Tamura model was selected in jMODELTEST [56], in line with the corrected Akaike information criterion (AICc); and the tree was inferred using MEGA 11.0 [57].
3. Results
3.1. Cytochrome Oxidase Subunit 1
We obtained a 587 bp fragment of the COI from a total of 159 C. myrmidone from Poland, including 137 (125 from 2022 and 12 from 2014) specimens from the Knyszyn Forest and 22 from Czerwony Bór (2014). This fragment was found to include 15 polymorphic sites (13 parsimony-informative sites). The variable sites represented 10 transitions and 5 transversions. The base composition of the C. myrmidone COI fragment was as follows: A—31.03%; C—37.06%; T—22.98%; G—8.93%. Among the 159 sequences obtained, we identified four haplotypes (Table 1, GenBank Acc. no: PV031351–54). However, just one of these proved clearly dominant in terms of the frequency of occurrence in Poland. This was CmCOI-1, which was found in 87% of the samples. This haplotype further corresponded to the haplotypes from GenBank that were previously identified in C. myrmidone (Table S2). CmCOI-1 was present in both investigated populations from Poland and was the only haplotype found in Czerwony Bór. CmCOI-2 was present in ≈12% of samples, and the remaining haplotypes (CmCOI-3 and CmCOI-4) were identified in single individuals from the Knyszyn Forest (Table 1).

Table 1.
Distribution of haplotypes and basic indicators of genetic diversity in the investigated populations of C. myrmidone, as estimated on the basis of a polymorphism of a (587 bp) fragment of mtDNA COI and absolute numbers of particular haplotypes in populations. N—sample size; H—number of haplotypes; h—haplotype diversity; π—nucleotide diversity; k—mean number of nucleotide differences among haplotypes; SSD—sum of squared deviations.
The overall haplotype diversity was low, at 0.224 ± 0.041, with an associated nucleotide diversity of 0.00452 ± 0.00086. The average number of nucleotide differences among haplotypes (k) was 2.652 ± 0.023 (Table 1). We did not find any evidence for the recent expansion in the populations from Poland, as indicated by non-significant values of Fu’s FS and Tajima’s D. Similarly, significant SSD suggested a deviation from the sudden expansion model. The relatively high value of the raggedness index (r) may indicate that the studied population has passed the demographic bottleneck.
The median-joining network for haplotypes identified in this study (Figure 3) revealed one “core haplotype”, which referred to 163 individuals, including 139 individuals from the Knyszyn Forest and Czerwony Bór (Table S1) and 24 individuals from GenBank (Table S2), as well as several satellite, low-frequency haplotypes found in the Knyszyn Forest (CmCOI-3, CmCOI-4) and GenBank, i.e., single individuals of C. caucasica from Greece (CmCOI-6, MW501563.1) and C. myrmidone from Russia (CmCOI-7, MW501352.1), differing by single substitution. However, we also identified a second haplogroup consisting of CmCOI-2, which was found in the Knyszyn Forest, and the case of a single individual from Russia (MW499355.1), as well as a closely related haplotype, identified in C. caucasica from Greece (CmCOI-8, MW499440.1) (Table S2). Both haplogroups differed by nine substitutions.
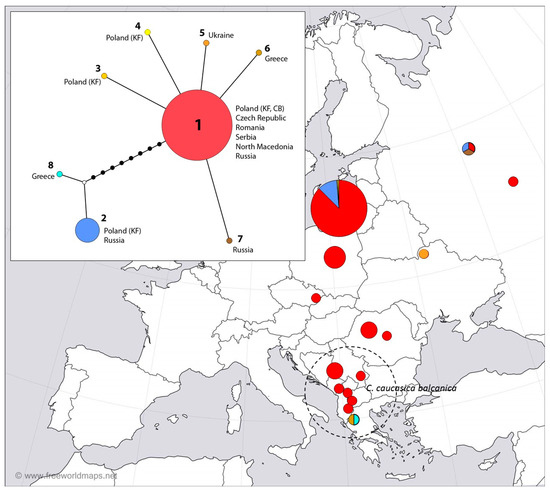
Figure 3.
Distribution of COI haplotypes among populations of C. myrmidone in Poland (KF—the Knyszyn Forest, CB—Czerwony Bór) and different localities of C. myrmidone and C. caucasica within its range (sequences available in the GenBank database). Haplotype network reconstruction was drawn using the median-joining method. The sizes of the circles on the haplotype network are directly proportional to the number of individuals analyzed.
The neighbour-joining phylogenetic tree (Figure 4) for the combined data set (novel haplotypes identified in this study and homologous sequences deposited in GenBank—49 haplotypes in total) revealed a surprising distribution of C. myrmidone COI haplotypes. Firstly, all sequences were grouped in a common clade with haplotypes of C. caucasica. Secondly, CmCOI-2, grouped with a C. caucasica haplotype from Greece, was located in a separated part of the phylogenetic tree, within a common clade with C. wiskotti. It should be noted that most nodes on the phylogenetic tree had low statistical support. However, the analysis highlighted the distinctiveness of the CmCOI-2 haplotype.

Figure 4.
The phylogenetic tree of COI haplotypes, inferred using the neighbour-joining method. All haplotypes identified in this study (CmCOI-1 to CmCOI-4) were included together with homologous sequences of European Colias sp. from GenBank. The optimal tree is shown. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (500 replicates) are shown next to the branches, but only values over 50% were displayed. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Tamura 3-parameter method, as described in Material and Methods. COI sequence of Eurema hecabe was used as an “out-group”.
3.2. Elongation Factor EF-1α
The successful amplification of a nuclear gene fragment occurred in 151 of the 159 individuals analyzed. In the 581 bp fragment, 18 polymorphic sites were found, which resulted in the identification of 18 haplotypes (GenBank Acc. no: PV033841–PV033858). In the Knyszyn Forest population, we found 14 haplotypes, and h was 0.600 (Table 2). We found no differences in haplotype frequencies in the two largest COI haplogroups in the Knyszyn Forest (CmCOI-1 vs. CmCOI-2; ΘST = 0.003; p > 0.05), while the genetic differentiation between samples collected in 2014 and 2022 was clearly higher and statistically significant (ΘST = 0.08; p < 0.05). In 2022, there were more haplotypes, but the haplotype diversity was higher in samples collected in 2014 (Table 2). This suggests that a higher number of haplotypes was strictly interlinked with a higher number of samples. Similarly, we found a higher nucleotide diversity (π) and k in the 2014 samples than in the 2022 samples. This suggests that genetic diversity has been gradually decreasing in the Knyszyn Forest over recent years.
Table 2.
Distribution of haplotypes and basic indicators of genetic diversity in the investigated populations of C. myrmidone in Poland, as estimated on the basis of a polymorphism of a (581 bp) fragment of the nuclear EF-1α gene and absolute numbers of particular haplotypes in populations. N—sample size; H—number of haplotypes; h—haplotype diversity; π—nucleotide diversity; k—mean number of nucleotide differences among haplotypes; SSD—sum of squared deviations.
In the case of Czerwony Bór, among 16 analyzed samples, we found as many as 9 haplotypes and a very high corresponding allelic diversity (h = 0.934). Interestingly, there was no significant genetic differentiation between samples from the Knyszyn Forest, collected in 2014, and Czerwony Bór (ΘST = 0.019; p > 0.05), but we found a much higher and significant differentiation for comparison between Czerwony Bór and the Knyszyn Forest in 2022 (ΘST = 0.16; p < 0.01). This suggests that, over the last decade, there has been a significant change in the frequency of nuclear alleles in the Knyszyn Forest as a result of, for example, increased genetic drift in an isolated population.
Neutrality tests were non-significant in the majority of analyzed groups of sequences, although negative values suggested some signs of the historical expansion of the species in central Europe. Moreover, we found a negative and significant value of the Fu’s test for overall analysis (Table 2), indicating that the excess of allele numbers in the population of C. myrmidone from Poland was possibly due to their recent expansion. Interestingly, estimating neutrality tests in COI haplogroups (CmCOI-1 and CmCOI-2), we found a negative and significant value of Fu’s test (FS = −5.781; p < 0.01) for haplogroup CmCOI-1 but not for haplogroup CmCOI-2 (Fs = −2.308; p > 0.05). It should be noted, however, that a non-significant value of Fu’s FS could be interlinked with a low sample size (N = 17 for haplogroup CmCOI-2 vs. N = 105 for haplogroup CmCOI-1). We found low values of raggedness index r and non-significant SSD, suggesting that the studied nuclear marker preserves traces of demographic expansion.
A comparison of the frequency of haplotype occurrence in both populations showed the presence of one dominant haplotype in terms of frequency haplotype—CmEF-1—occurring in almost 60% of individuals. The second most frequent haplotype—CmEF-5—was found in slightly over 12% of individuals. Both haplotypes differ by only two substitutions and occurred in both studied populations. In the Knyszyn Forest, CmEF-1 was definitely dominant in terms of frequency, while in Czerwony Bór, it was one of the two most frequent haplotypes (Table 2). In general, the studied populations clearly differed in terms of the occurrence of the EF-1α haplotypes. Only four haplotypes were shared between both populations, while as many as fourteen haplotypes were unique to one or the other (Figure 5).
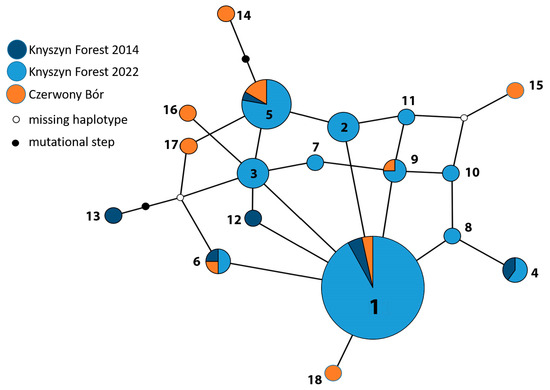
Figure 5.
The median-joining network of 18 haplotypes was identified in this study. To simplify the picture, in the case of low-frequency haplotypes, the prefix “CmEF” was omitted from the haplotype names. The size of a circle is approximately proportional to the frequency of occurrence of the given haplotype in the sample of 151 individuals. Each line connecting two circles represents a single substitution differentiating the haplotypes.
The haplotype network (Figure 5) was typical for EF-1α, indicating that the dominant haplotype—CmEF-1—occupied the core position, while most of its satellite haplotypes differed by single substitutions and showed a low frequency. Additionally, many alternative linkages were found between the satellite haplotypes. A characteristic feature of this network is the presence of a second, relatively frequent haplotype elsewhere in the network. CmEF-5 was also found in both populations and seems to be quite frequent in the Knyszyn Forest, although it occupies a distal position in the network.
3.3. Wolbachia Infection
To explore the prevalence of Wolbachia infection, Wolbachia DNA was detected in C. myrmidone samples via PCR amplification of the surface protein gene wsp. We tested 53 samples from the Knyszyn Forest, collected in 2022, including 26 males and 27 females. Only the two most frequent haplotypes were screened. We found that 30% of C. myrmidone in the Knyszyn Forest were positive for Wolbachia. Considering the sex of the infected individuals, we found 11 infected females and only 4 males. Considering the COI haplogroups, we found infected individuals only within the haplogroup CmCOI-2, and no infections were observed in the haplogroup CmCOI-1. The analysis of the sequence of the amplified fragment of the wsp gene indicated the presence of three haplotypes, differing by a single substitution. A comparison of the obtained sequences from the Wolbachia sequences deposited in GenBank indicated that the identified strain was not previously identified in the Colias species. Instead, two Wolbachia haplotypes identified in this study (CmWsp-1, CmWsp-2; GenBank Acc. no: PV085849–PV085850) exhibit a very high sequence similarity (99.82 and 99.86, respectively) with one of the Wolbachia strains identified previously in Pseudophilotes bavius (GenBank Acc. no: MK784199.1) [58].
4. Discussion
4.1. Genetic Diversity of the Last Remaining Polish Population
While analyzing the level of genetic variability in the mitochondrial COI gene in the Knyszyn Forest, we found the occurrence of four haplotypes (CmCOI-1–4), which is the same number as those found so far within the species [29]. Nonetheless, we should assume that haplotype and nucleotide diversity were rather low. Here, it should be noted that previous studies indicate that the diversity in COI in terms of the representatives of Colias Fabricius, 1807 across Europe, is relatively low compared to other butterfly genera [28,29,59]. This pattern can be explained to some extent by the short evolutionary history of the Colias genus, which evolved only ~3.5 Ma [25]. Nonetheless, the presence of four COI haplotypes in spatially and temporally (for at least 20 years) isolated populations seems surprising, especially considering that we found only one haplotype in the recently extirpated Czerwony Bór population. However, the identification of only one haplotype in Czerwony Bór, as well as in the Knyszyn Forest in 2014, may be related to the small sample from the population. It is unlikely that additional haplotypes appeared in the isolated Knyszyn Forest after 2014. It should be assumed that when analyzing only a few individuals, only the haplotype would be detected, which would definitely be dominant in terms of frequency.
A comprehensive data set of C. myrmidone COI sequences indicated the presence of two haplogroups, but the one with CmCOI-1 as the central haplotype is definitely more widespread. Among European butterflies, the star-shaped topology of the COI network (a low number of prevalent haplotypes and numerous, but of low frequency variants resulting from subsequent mutations) is the most typical [29], which can be seen in Phengaris arion [60]. In the case of clearly distinct lineages, the pattern can be different with several relatively equal clades, e.g., for Parnassius mnemosyne [61]. CmCOI-2 belonging to the separate haplogroup differed from CmCOI-1 by nine substitutions. Even higher intraspecific divergences have been reported for Colias palaeno, where most individuals were placed in the typical star-like network, but three haplotypes were clearly distinct (differing by up to as many as 50 mutations) and were likely the result of introgression from other Colias species [62]. For C. myrmidone, there is no indication of such a scenario, but the possibility of ancient hybridization cannot be completely ruled out.
The number of EF-1α alleles in the present study was clearly higher in both localities compared to the number of COI haplotypes. Divergent patterns of mtDNA and nDNA diversity are not uncommon among butterflies, and a higher sequence polymorphism in EF-1α than COI seems to be common [60,63,64,65,66]. Mitochondrial lineages exhibit a much faster lineage sorting rate, and the random elimination of haplotypes is much faster than the nuclear allele extinction rate; therefore, evolutionary relationships could be oversimplified [67].
Accordingly, we found a moderate genetic diversity in EF-1α (h = 0.60); thus, we found no evidence that isolation reduced genetic diversity, which may be surprising for such a highly endangered species. However, in butterflies, the level of genetic diversity has been shown to be unrelated to their abundance or natural history traits [68]. The relationship between population size and genetic variability is not straightforward. In general, there are examples of populations that exist despite low numbers and reduced genetic variability [69]. Conversely, there are known populations that have experienced significant declines despite having a rich gene pool [70]. Also, some populations maintain a higher-than-predicted heterozygosity and allelic diversity, although they are small and long-isolated [71]. In the case of the Melitaea cinxia metapopulation, the relationship between heterozygosity and extinction disappeared when demographic and environmental variables were introduced into the model analyses, indicating that the association between genetic diversity and extinction could be detected only under some conditions. For example, the rate of extinction does not decline with increasing heterozygosity in small populations, whereas such a pattern is observed in the case of large populations [22]. Hence, we can assume that our study population from the Knyszyn Forest could be another example of a small, isolated population, maintaining quite a high genetic diversity.
In the face of the complete lack of gene flow in recent years resulting from isolation [4], one possible explanation is that the current levels of genetic diversity reflect pre-disturbance values, and insufficient time has passed for a detectable reduction in genetic diversity. The population of C. myrmidone in Poland collapsed at the beginning of the 21st century, and the butterfly survived only in several isolated regions. In 2022, i.e., the year of intensive sampling, the metapopulation in the Knyszyn Forest was relatively large due to the previous enlargement of the habitat network being a consequence of intensive forestry, creating ephemeral patches [4].
The Knyszyn Forest is located in Northeastern Poland. For many animal species, including butterflies, a high genetic diversity on mtDNA was found here, which can be attributed to the secondary contact zone of lineages from different glacial refuges [72,73,74,75]; this is an example that contradicts the “southern richness, northern purity” paradigm. Secondary contact following postglacial recolonization and interspecific hybridization may be crucial in shaping the pattern of genetic diversity of populations in western Eurasia, especially in insects [76].
Nonetheless, the genetic diversity of the Knyszyn Forest population seems to have gradually decreased. Despite the lower sample size, we found clearly higher indices of genetic diversity in the group of specimens from 2014 than from the 2022 group. Moreover, genetic differentiation in EF-1α allele frequency was found between both periods. Similarly, mean values of genetic diversity indices for EF-1α were higher in Czerwony Bór, where samples were also collected in 2014. Nuclear data also suggest that the Polish population used to be genetically uniform, as we did not confirm significant genetic differentiation between the Knyszyn Forest and Czerwony Bór for samples from 2014.
According to the classification of Habel and Schmitt [20], C. myrmidone can be considered as an intermediate species between specialists and generalists. Such species often occur in large population networks and are characterized by a relatively high genetic diversity, which is sustained by gene flow. Population connectivity rescues genetic diversity after a demographic bottleneck [77] and a lack of interconnectivity due to the fragmentation, degradation, and isolation of habitats, which may lead quickly to inbreeding depressions contrasting to specialists. As a consequence, populations suffer from sudden genetic impoverishment and possibly reduced adaptability (see, e.g., [78]) and, therefore, may be at the highest risk of extinction even compared to specialists, for which a low genetic diversity is often typical. The fragmentation and isolation of breeding areas are considered as one of the main causes of the collapse of the Hungarian population [11]. Although our study suggests a depletion of the gene pool in the Knyszyn Forest, an analysis of genetic material from more distant periods is needed to confirm that C. myrmidone fits in with the predictions for intermediate species in terms of genetic diversity. The source of such specimens may be museums and private collections. Additional molecular markers, relying on short DNA fragments (e.g., microsatellites or single-nucleotide polymorphisms), would be more appropriate for often fragmented genetic material [79].
Indeed, in the past, C. myrmidone used to be considered a mobile species, but studies of the metapopulation in the Knyszyn Forest show that when the inhabited area is very limited in space, a selection against dispersal is very likely to occur [4,80]. It can be suspected that there was a significant reduction in numbers in the Knyszyn Forest around 2019 (after the first sampling), i.e., when the species was extirpated in Czerwony Bór, which, in turn, was probably largely related to extreme weather conditions. Therefore, both bottleneck and genetic drift resulting from isolation may explain the observed pattern of diversity and differentiation. Our studies show that the process of genetic impoverishment can progress very quickly, even within a decade, also affecting the frequency of alleles. As a result, genetic differentiation between the Knyszyn Forest and Czerwony Bór has clearly increased over a period of ten years. Here, it should be mentioned that C. myrmidone is bivoltine and, in some years, even a three-generation species. The first (spring) brood can be clearly less numerous than the summer one [4], so intra-seasonal breakdowns are a serious problem and a threat to the existence of the population both directly and indirectly.
4.2. Wolbachia in Colias Myrmidone
The presence of Wolbachia was reported in C. myrmidone for the first time, and all infected individuals shared the CmCOI-2 haplotype. Our screening indicated a 100% prevalence in this genotype, whereas in the case of the most common and widespread CmCOI-1 haplotype, we did not record any cases of infection. The observed pattern can be easily explained by unidirectional cytoplasmic incompatibility, i.e., males infected with Wolbachia are not able to produce functional zygotes with uninfected females or those that do not host the same Wolbachia strain. Females, on the contrary, if infected, produce offspring regardless of their mate’s infection status [36,81]. Wolbachia is transmitted maternally, but nuclear gene flow between mtDNA lineages is not affected. This is consistent with our data indicating no differentiation in EF-1α between infected and uninfected genotypes. However, with the intermediate frequency of the Wolbachia infection, i.e., when not all individuals are infected, the patterns of infection can be surprising, e.g., in the case of Colias mongola/C. tamerlana when Wolbachia was exclusively detected in one haplogroup and was restricted to (not all) females randomly found in the geographically remote populations [82]. On the contrary, for two congeneric species, i.e., C. croceus and C. erate, studies in Slovakia indicate 100% infection [83].
The Wolbachia strain identified in our sample has been recorded so far only in an unrelated butterfly species—Pseudophilotes bavius [58]. Because either the origin of the infection or the impact on the host was not resolved by that study and remains unknown, it would be interesting to perform a detailed comparative examination of both species sharing the same Wolbachia sequence. The only identified strain in the population of any Colias species in Europe, i.e., ST141 belonging to supergroup B, exhibits a perfect vertical transmission and is known to induce strong cytoplasmic incompatibility when different infection status or different strains of a male and a female can cause the death of developing embryos [35].
The presence of a strain other than that previously detected in Colias spp., combined with the fact that the only infected haplotype (CmCOI-2) differs from the most common haplotype in the Polish population by a relatively high number of substitutions, and in phylogenetic analyses occupies a position in a separate part of the tree (close to C. wisskoti), which may suggest the transmission of the endoparasite by interspecies hybridization [84]. This concept, however, requires confirmation by further studies.
It is more puzzling that the prevalence of the CmCOI-2 haplotype and, therefore, the prevalence of infected genotypes was clearly, although incompletely, female-biased in the Knyszyn Forest (26.9% females and only 6.1% males), while the sex ratios in the studied metapopulation were fairly well balanced, as estimated during intensive mark–release–recapture performed in the same year [4]. Such cases are very rare, and the mechanisms behind this phenomenon, as well as their consequences, are insufficiently understood [85]. Narita et al. [86] suggest that a single strain of Wolbachia may induce two distinct reproductive manipulations in the same host. Additionally, complete sex biases resulting from male killing or feminization caused by Wolbachia are well documented for lepidopterans [87,88,89].
We did not find the “Wolbachia-infected” haplotype (CmCOI-2) in material collected a decade earlier; however, it should be noted that, either in the Knyszyn Forest and Czerwony Bór, a significantly smaller number of individuals was sampled. Moreover, information on the sex of most individuals was missing (because it was regarded as redundant at that time by a collector). Sampling for genetic studies is frequently male-biased due to the higher catchability of this sex, which is related to more conspicuous behavior. Studies often tend to sample material from males or simply collect worn males (see, e.g., [90]), as they are considered less important for populations than gravid females. This concern is often even more emphasized in the case of scarce and/or endangered species. Our experience shows that complete information and/or unbiased sampling would be recommended. We suspect that CmCOI-2 was present in the Knyszyn Forest in 2014, but this issue cannot be resolved in relation to Czerwony Bór, taking the sampling constraints into consideration. Samples were collected from a dozen or so individuals and mostly males, among which the “Wolbachia-infected” haplotype (CmCOI-2) was rare. The same could be applied to all C. myrmidone populations sampled for barcoding libraries to date, especially those from which there have been sequences from only 11 individuals available until now. The presence of Wolbachia in C. myrmidone individuals, for which sequences were deposited in GenBank, remains unknown; it would also be especially interesting to obtain insight into populations from Russia, where CmCOI-2 was previously detected.
Mitochondrial DNA not being neutral, as was considered in the past, may influence some functional traits, including adaptation to climatic conditions [91]. Variation in mtDNA sequences may, e.g., potentially counterbalance some environmental changes, and adaptive mutations spread, subsequently producing selective sweeps in populations [92]. Therefore, the presence of both infected and uninfected genotypes (as detected in the Knyszyn Forest) could be beneficial for the population in some way, but further studies are needed. The focal population history of infection is unknown, and it is not possible to judge whether the Wolbachia prevalence rate, estimated at 14.4% for samples collected in 2022, is stable among seasons.
The only identified strain (ST141 belonging to supergroup B) in the population of any Colias in Europe exhibits perfect vertical transmission, and it is known to induce strong cytoplasmic incompatibility when different infection status or different strains of a male and a female can cause death of developing embryos [83]. Studies of C. erate in Japan indicate that Wolbachia seems to have positive effects on host fitness, as assessed by the survival rates of the infected brood [93].
4.3. Differentiation Between Colias myrmidone and Colias caucasica
Interestingly, the most common CmCOI-1 haplotype detected in C. myrmidone samples was also shared with Colias caucasica balcanica. Searching GenBank, we found that 16 of 18 sequences were identical to CmCOI-1, and they were detected in individuals from Macedonia and Serbia. Only two specimens of C. caucasica from Greece had distinct and unique haplotypes that differed significantly (14 substitutions) from each other, with one being very close to CmCOI-1 (differing by a single substitution) and another one differing from CmCOI-2 by two substitutions. This may suggest that two different haplogroups may also occur in the Balkans.
Barcode sharing is reported for 69 butterfly species (15%) occurring in Europe and may be explained by biological processes (e.g., hybridization resulting in introgression or incomplete lineage sorting), as well as uncertainties related to taxonomy or simply the misidentification of sampled material [29]. Then, among Colias butterflies, introgression following hybridization may explain the observed barcode sharing as seen in, e.g., C. behrii and C. meadii in North America [94].
In the case of C. myrmidone/C. caucasica, incomplete lineage sorting cannot be excluded. It seems unlikely that barcode sharing is related to Wolbachia infection since we did not find infected individuals within the CmCOI-1 haplogroup. Recent analyses based on sequences of multiple genes indicate that C. myrmidone and Colias caucasica Staudinger, 1871 are sister species, separated by ~1.2 Ma [25]. However, only two individuals (from Russia and Georgia, respectively) were included. Further studies are desired, especially taking into consideration the ca. 1500 km disjunction between nominotypical C. caucasica occupying mountains of eastern Turkey [95] and part of the Caucasus [96], as well as the nearest populations of C. caucasica balcanica, which is considered as an endemic Balkan subspecies ranging from Croatia to Greece [97,98,99,100,101,102,103]. Initially, it was considered as a mountain variant of C. myrmidone, a subspecies of C. myrmidone, or a distinct species, Colias balcanica [104], and the present status was evaluated by Wegner [105]. Generally, C. caucasica resembles C. myrmidone, but it is somewhat larger, and in males, the ground color of the wing upper side is a deeper orange [106]; however, both taxa share the same configuration of UV pattern on their wings [107]. Colias caucasica inhabits mostly alpine grasslands (850–2300 m) and uses local Chamaecytisus species (e.g., Ch. hirsutus) as larval food plants. Contrasting to C. myrmidone, there is a single generation per year in nature, but the second brood is easily produced in captive rearing [106], which suggests that some traits may be adapted to harsh environmental conditions at high altitudes.
There are approximately 90 described Colias species—the vast majority of which are Holoarctic ones—which makes this genus one of the largest among Pieridae [108,109]. Although they are quite spectacular butterflies with a long history of both collecting and studies, there are still a lot of taxonomic ambiguities resulting from intraspecies variation in wing pattern. At the same time, genitalia structures are largely uniform in this genus and do not possess reliable diagnostic features contrasting to most other lepidopterans. Moreover, allopatric populations with similar phenotypes were traditionally treated as separate species. Many weakly supported subspecies have already been synonymised [110]. Recent studies have investigated the conspecificity of C. mongola and C. tamerlana, enigmatic taxa in which COI haplotypes display four strongly supported lineages while nuclear markers demonstrate a very shallow divergence [82].
In particular, the case of C. myrmidone and C. caucasica, including C. c. balcanica, clearly shows that an ultimate taxonomic decision requires extensive morphological, ecological, genetic, and/or molecular evidence. We are unanimous with Dincă et al. [29] that further research is recommended and simultaneously emphasizes the need for extensive analyses on Wolbachia prevalence in populations of C. myrmidone/C. caucasica, as Wolbachia-mediated mitochondrial introgressions may obscure taxonomy [19,111,112].
4.4. Conservation Implications
The last Polish population retained a relatively high genetic diversity, although we also found signs of recent deterioration. To counteract this process, it is vital to conserve a network of habitats to minimize the probability of serious bottlenecks resulting from both abiotic and biotic (e.g., parasitoids) factors affecting seasonal abundance. However, to increase genetic diversity, assisted gene flow should be considered, i.e., exchanging individuals with other extant European populations. Such actions should be taken very carefully and proceeded by genetic studies to avoid negative effects related to Wolbachia.
The issue of evolutionary relationships between C. myrmidone and C. caucasica balcanica also remains to be resolved. Final conclusions will be important from a conservation point of view. If Colias caucasica balcanica is not distinct from C. myrmidone, the range of the latter in Europe would actually be wider, and the Balkan EU countries (Bulgaria, Croatia, and Greece) will receive a new Natura 2000 species because C. caucasica is not listed in the Habitats Directive.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/insects16020220/s1, Table S1: Data summary for samples of Colias myrmidone collected in Poland and used in this study; Table S2: Data summary for the COI sequences of Colias myrmidone and C. caucasica available in GenBank and used in this study.
Author Contributions
Conceptualization, A.G., R.R. and M.S.; methodology, A.G.; software, A.G.; validation, A.G., R.R. and M.S.; formal analysis, A.G.; investigation, A.G.; resources, M.S.; data curation, A.G.; writing—original draft preparation, A.G. and M.S.; writing—review and editing, A.G. and M.S.; visualization, A.G. and M.S.; supervision, R.R. and M.S.; project administration, M.S.; funding acquisition, M.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research has received financial support from the General Directorate of the Polish State Forests under a project entitled “Forest management for endangered species: conservation of the Danube Clouded Yellow (Colias myrmidone) in the Knyszyn Forest” (contract no. EZ.271.3.7.2021).
Data Availability Statement
All the analyzed DNA sequences are available via the GenBank links provided.
Acknowledgments
The Regional Director for Environmental Protection in Białystok provided relevant permissions for sampling of Colias myrmidone for genetic studies. Three anonymous reviewers made valuable comments on the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Van Swaay, C.A.M.; Warren, M.S. Red Data Book of European Butterflies (Rhopalocera); Nature and Environment No. 99; Council of Europe: Strasbourg, France, 1999. [Google Scholar]
- Maes, D.; Verovnik, R.; Wiemers, M.; Brosens, D.; Beshkov, S.; Bonelli, S.; Buszko, J.; Cantú-Salazar, L.; Cassar, L.-F.; Collins, S.; et al. Integrating national Red Lists for prioritising conservation actions for European butterflies. J. Insect Conserv. 2019, 23, 301–330. [Google Scholar] [CrossRef]
- Van Swaay, C.A.M.; Cuttelod, A.; Collins, S.; Maes, D.; López Munguira, M.; Šašić, M.; Settele, J.; Verovnik, R.; Verstrael, T.; Warren, M.; et al. European Red List of Butterflies; Publications Office of the European Union: Luxembourg, 2010. [Google Scholar]
- Sielezniew, M.; Jaworski, T.; Sielezniew, I.; Deoniziak, K.; Bystrowski, C.; Hilszczański, J.; Nowicki, P. Clear-cuts support the metapopulation of a critically endangered butterfly. For. Ecol. Manag. 2024, 562, 121939. [Google Scholar] [CrossRef]
- Loos, J.; Schröer, C.; Becker, T.; Kastal, A.; Kortmann, E.; Dolek, M. Mosaic landscapes provide conservation pockets for an endangered species: Colias myrmidone in Romania. Insect Conserv. Divers. 2022, 15, 359–369. [Google Scholar] [CrossRef]
- Kudrna, O.; Mayer, L. Grundlagen zu einem Artenhilfsprogramm für Colias myrmidone (Esper, 1780) in Bayern. Oedippus 1990, 1, 1–46. [Google Scholar]
- Freese, A.; Dolek, M.; Geyer, A.; Stetter, H. Biology, distribution, and extinction of Colias myrmidone (Lepidoptera, Pieridae) in Bavaria and its situation in other European countries. J. Res. Lepid. 2005, 38, 51–58. [Google Scholar] [CrossRef]
- Konvička, M.; Benes, J.; Cizek, O.; Kopecek, F.; Konvicka, O.; Vitaz, L. How too much care kills species: Grassland reserves, agri-environmental schemes and extinction of Colias myrmidone (Lepidoptera: Pieridae) from its former stronghold. J. Insect Conserv. 2008, 12, 519–525. [Google Scholar] [CrossRef]
- Marhoul, P.; Dolek, M. Action Plan for the Conservation of the Danube Clouded Yellow Colias myrmidone in the European Union; European Commission: Brussels, Belgium, 2012. [Google Scholar]
- Szentirmai, I.; Mesterházy, A.; Varga, I.; Schubert, Z.; Sándor, L.C.; Ábrahám, L.; Kőrösi, Á. Habitat use and population biology of the Danube Clouded Yellow butterfly Colias myrmidone (Lepidoptera: Pieridae) in Romania. J. Insect Conserv. 2014, 18, 417–425. [Google Scholar] [CrossRef]
- Bálint, Z.; Sáfián, S. Narancsszínű kéneslepke Colias myrmidone (Esper 1781). In Natura 2000 Fajok és Élőhelyek Magyarországon; Haraszthy, L., Ed.; Pro Vértes Közalapítvány: Csákvár, Hungary, 2014; pp. 306–307. [Google Scholar]
- Settele, J.; Kudrna, O.; Harpke, A.; Kühn, I.; van Swaay, C.; Verovnik, R.; Warren, M.; Wiemers, M.; Hanspach, J.; Hickler, T.; et al. Climatic Risk Atlas of European Butterflies; Pensoft: Sofia, Bulgaria, 2008. [Google Scholar]
- Steiner, C.F.; Asgari, M. Habitat isolation reduces intra- and interspecific biodiversity and stability. R. Soc. Open Sci. 2022, 9, 211309. [Google Scholar] [CrossRef]
- Spielman, D.; Brook, B.W.; Frankham, R. Most species are not driven to extinction before genetic factors impact them. Proc. Natl. Acad. Sci. USA 2004, 101, 15261–15264. [Google Scholar] [CrossRef]
- Frankham, R. Genetics and extinction. Biol. Conserv. 2005, 126, 131–140. [Google Scholar] [CrossRef]
- Kardos, M.; Armstrong, E.E.; Fitzpatrick, S.W.; Hauser, S.; Hedrick, P.W.; Miller, J.M.; Tallmon, D.A.; Funk, W.C. The crucial role of genome-wide genetic variation in conservation. Proc. Natl. Acad. Sci. USA 2021, 118, e2104642118. [Google Scholar] [CrossRef] [PubMed]
- Keyghobadi, N.; Roland, J.E.N.S.; Strobeck, C. Genetic differentiation and gene flow among populations of the alpine butterfly, Parnassius smintheus, vary with landscape connectivity. Mol. Ecol. 2005, 14, 1897–1909. [Google Scholar] [CrossRef]
- Lynch, M.; Lande, R. Evolution and extinction in response to environmental change. In Biotic Interactions and Global Change; Kareiva, P.M., Kingsolver, J.G., Huey, R.B., Eds.; Sinauer: Sunderland, MA, USA, 1993; pp. 234–250. [Google Scholar]
- Smith, M.A.; Bertrand, C.; Crosby, K.; Eveleigh, E.S.; Fernandez-Triana, J.; Fisher, B.L.; Gibbs, J.; Hajibabaei, M.; Hallwachs, W.; Hind, K.; et al. Wolbachia and DNA barcoding insects: Patterns, potential, and problems. PLoS ONE 2012, 7, e36514. [Google Scholar] [CrossRef] [PubMed]
- Habel, J.C.; Schmitt, T. The burden of genetic diversity. Biol. Conserv. 2012, 147, 270–274. [Google Scholar] [CrossRef]
- Petit-Marty, N.; Vázquez-Luis, M.; Hendriks, I.E. Use of the nucleotide diversity in COI mitochondrial gene as an early diagnostic of conservation status of animal species. Conserv. Lett. 2021, 14, e12756. [Google Scholar] [CrossRef]
- DiLeo, M.F.; Nair, A.; Kardos, M.; Husby, A.; Saastamoinen, M. Demography and environment modulate the effects of genetic diversity on extinction risk in a butterfly metapopulation. Proc. Natl. Acad. Sci. USA 2024, 121, e2309455121. [Google Scholar] [CrossRef]
- Alves, F.; Banks, S.C.; Edworthy, M.; Stojanovic, D.; Langmore, N.E.; Heinsohn, R. Using conservation genetics to prioritise management options for an endangered songbird. Heredity 2023, 130, 289–301. [Google Scholar] [CrossRef]
- van der Valk, T.; Dalèn, L. From genomic threat assessment to conservation action. Cell 2024, 187, 1038–1041. [Google Scholar] [CrossRef]
- Mo, S.; Zhu, Y.; Braga, M.P.; Lohman, D.J.; Nylin, S.; Moumou, A.; Wheat, C.W.; Wahlberg, N.; Wang, M.; Ma, F.; et al. Rapid evolution of host repertoire and geographic range in a young and diverse genus of montane butterflies. Syst. Biol. 2024, 74, 141–157. [Google Scholar] [CrossRef]
- Dincă, V.; Bálint, Z.; Vodă, R.; Dapporto, L.; Hebert, P.D.; Vila, R. Use of genetic, climatic, and microbiological data to inform reintroduction of a regionally extinct butterfly. Conserv. Biol. 2018, 32, 828–837. [Google Scholar] [CrossRef]
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological identifications through DNA barcodes. Proc. Roy. Soc. Lond. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Dincă, V.; Zakharov, E.V.; Hebert, P.D.; Vila, R. Complete DNA barcode reference library for a country’s butterfly fauna reveals high performance for temperate Europe. Proc. Roy. Soc. Lond. B Biol. Sci. 2011, 278, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Dincă, V.; Dapporto, L.; Somervuo, P.; Vodă, R.; Cuvelier, S.; Gascoigne-Pees, M.; Huemer, P.; Mutanen, M.; Hebert, P.D.N.; Vila, R. High resolution DNA barcode library for European butterflies reveals continental patterns of mitochondrial genetic diversity. Commun. Biol. 2021, 4, 315. [Google Scholar] [CrossRef]
- Kulak, A.W. Colias myrmidone (Esper, 1771). In Krasnaya kniga Respubliki Belarusʹ. Zhivotnyye, 4th ed.; Katchanovski, I.M., Nikiforov, M.E., Parfienov, W.I., Eds.; Belorusskaya Entsiklopediya im. Petrusja Brovki: Minsk, Belarus, 2015; pp. 208–209. [Google Scholar]
- Wahlberg, N.; Brower, A.V.Z.; Nylin, S. Phylogenetic relationships and historical biogeography of tribes and genera in the subfamily Nymphalinae (Lepidoptera: Nymphalidae). Biol. J. Linn. Soc. 2005, 86, 227–251. [Google Scholar] [CrossRef]
- Kodandaramaiah, U.; Wahlberg, N. Phylogeny and biogeography of Coenonympha butterflies (Nymphalidae: Satyrinae)—Patterns of colonization in the Holarctic. Syst. Entomol. 2009, 34, 315–323. [Google Scholar] [CrossRef]
- Dapporto, L.; Cini, A.; Vodă, R.; Dincă, V.; Wiemers, M.; Menchetti, M.; Magini, G.; Talavera, G.; Shreeve, T.; Bonelli, S.; et al. Integrating three comprehensive data sets shows that mitochondrial DNA variation is linked to species traits and paleogeographic events in European butterflies. Mol. Ecol. Resour. 2019, 19, 1623–1636. [Google Scholar] [CrossRef] [PubMed]
- Hilgenboecker, K.; Hammerstein, P.; Schlattmann, P.; Telschow, A.; Werren, J.H. How many species are infected with Wolbachia?—A statistical analysis of current data. FEMS Microbiol. Lett. 2008, 281, 215–220. [Google Scholar] [CrossRef]
- Charlat, S.; Hurst, G.D.D.; Merçot, H. Evolutionary consequences of Wolbachia infection. Trends Genet. 2003, 19, 217–223. [Google Scholar] [CrossRef]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef]
- Wendt, M.; Kulanek, D.; Varga, Z.; Rákosy, L.; Schmitt, T. Pronounced mito-nuclear discordance and various Wolbachia infections in the water ringlet Erebia pronoe have resulted in a complex phylogeographic structure. Sci. Rep. 2022, 12, 5175. [Google Scholar] [CrossRef]
- Bartoňová, A.S.; Konvička, M.; Marešová, J.; Wiemers, M.; Ignatev, N.; Wahlberg, N.; Schmitt, T.; Fric, Z.F. Wolbachia affects mitochondrial population structure in two systems of closely related Palaearctic blue butterflies. Sci. Rep. 2021, 11, 3019. [Google Scholar] [CrossRef]
- Ritter, S.; Michalski, S.G.; Settele, J.; Wiemers, M.; Fric, Z.F.; Sielezniew, M.; Šašić, M.; Rozier, Y.; Durka, W. Wolbachia infections mimic cryptic speciation in two parasitic butterfly species, Phengaris teleius and P. nausithous (Lepidoptera: Lycaenidae). PLoS ONE 2013, 8, e78107. [Google Scholar] [CrossRef] [PubMed]
- Nice, C.C.; Gompert, Z.; Forister, M.L.; Fordyce, J.A. An unseen foe in arthropod conservation efforts: The case of Wolbachia infections in the Karner blue butterfly. Biol. Conserv. 2009, 142, 3137–3146. [Google Scholar] [CrossRef]
- Hamm, C.A.; Handley, C.A.; Pike, A.; Forister, M.L.; Fordyce, J.A.; Nice, C.C. Wolbachia infection and Lepidoptera of conservation concern. J. Insect Sci. 2014, 14, 6. [Google Scholar] [CrossRef] [PubMed]
- Buszko, J. Colias myrmidone (Esper 1780)—Szlaczkoń szafraniec. In Polish Red Data Book of Animals. Invertebrates; Głowaciński, Z., Nowacki, J., Eds.; Institute of Nature Conservation PAS: Kraków, Poland, 2004; pp. 243–244. [Google Scholar]
- Marschalek, D.A.; Jesu, J.A.; Berres, M.E. Impact of non-lethal genetic sampling on the survival, longevity and behaviour of the Hermes copper (Lycaena hermes) butterfly. Insect Conserv. Divers. 2013, 6, 658–662. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Kim, M.I.; Wan, X.; Kim, M.J.; Jeong, H.C.; Ahn, N.H.; Kim, K.G.; Han, Y.S.; Kim, I. Phylogenetic relationships of true butterflies (Lepidoptera: Papilionoidea) inferred from COI, 16S rRNA and EF-1α sequences. Mol. Cells 2010, 30, 409–425. [Google Scholar] [CrossRef]
- Monteiro, A.; Pierce, N. Phylogeny of Bicyclus (Lepidoptera: Nymphalidae) inferred from COI, COII, and EF1-α gene sequences. Mol. Phylogenet. Evol. 2001, 18, 264–281. [Google Scholar] [CrossRef]
- Braig, H.R.; Zhou, W.; Dobson, S.L.; O’Neill, S.L. Cloning and characterization of a gene encoding the major surface protein of the bacterial endosymbiont Wolbachia pipientis. J. Bacteriol. 1998, 180, 2373–2378. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Res. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Tajima, F. Statistical method for testing the neutral mutational hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Rogers, A.R.; Harpending, H. Population growth makes waves in the distribution of pairwise genetic differences. Mol. Biol. Evol. 1992, 9, 552–569. [Google Scholar]
- Harpending, H.C.; Sherry, S.T.; Rogers, A.R.; Stoneking, M. Genetic structure of ancient human populations. Curr. Anthropol. 1993, 34, 483–496. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Sucháčková Bartoňová, A.; Konvička, M.; Marešová, J.; Kolev, Z.; Wahlberg, N.; Fric, Z.F. Recently lost connectivity in the Western Palaearctic steppes: The case of a scarce specialist butterfly. Conserv. Gen. 2020, 21, 561–575. [Google Scholar] [CrossRef]
- Dapporto, L.; Menchetti, M.; Vodă, R.; Corbella, C.; Cuvelier, S.; Djemadi, I.; Gascoigne-Pees, M.; Hinojosa, J.C.; Lam, N.T.; Serracanta, M.; et al. The atlas of mitochondrial genetic diversity for Western Palaearctic butterflies. Glob. Ecol. Biogeogr. 2022, 31, 2184–2190. [Google Scholar] [CrossRef]
- Patricelli, D.; Sielezniew, M.; Ponikwicka-Tyszko, D.; Ratkiewicz, M.; Bonelli, S.; Barbero, F.; Witek, M.; Buś, M.M.; Rutkowski, R.; Balletto, E. Contrasting genetic structure of rear edge and continuous range populations of a parasitic butterfly infected by Wolbachia. BMC Evol. Biol. 2013, 13, 14. [Google Scholar] [CrossRef] [PubMed]
- Gratton, P.; Konopiński, M.K.; Sbordoni, V. Pleistocene evolutionary history of the Clouded Apollo (Parnassius mnemosyne): Genetic signatures of climate cycles and a ‘time-dependent’ mitochondrial substitution rate. Mol. Ecol. 2008, 17, 4248–4262. [Google Scholar] [CrossRef] [PubMed]
- Kramp, K.; Cizek, O.; Madeira, P.M.; Ramos, A.A.; Konvicka, M.; Castilho, R.; Schmitt, T. Genetic implications of phylogeographical patterns in the conservation of the boreal wetland butterfly Colias palaeno (Pieridae). Biol. J. Linn. Soc. 2016, 119, 1068–1081. [Google Scholar] [CrossRef]
- Sielezniew, M.; Ponikwicka-Tyszko, D.; Ratkiewicz, M.; Dziekanska, I.; Kostro-Ambroziak, A.; Rutkowski, R. Diverging patterns of mitochondrial and nuclear diversity in the specialized butterfly Plebejus argus (Lepidoptera: Lycaenidae). Eur. J. Entomol. 2011, 108, 537–545. [Google Scholar] [CrossRef]
- Sielezniew, M.; Rutkowski, R.; Ponikwicka-Tyszko, D.; Ratkiewicz, M.; Dziekańska, I.; Švitra, G. Differences in genetic variability between two ecotypes of endangered myrmecophilous butterfly Phengaris (=Maculinea) alcon—The setting of conservation priorities. Insect Conserv. Divers. 2012, 5, 223–236. [Google Scholar] [CrossRef]
- Czajkowska, M.; Dawidowicz, Ł.; Borkowska, A.; Dziekańska, I.; Sielezniew, M. Population genetic structure and demography of the critically endangered Chequered blue butterfly (Scolitantides orion) in a highly isolated part of its distribution range. Insects 2020, 11, 608. [Google Scholar] [CrossRef]
- Zupan, S.; Jugovic, J.; Čelik, T.; Buzan, E. Population genetic structure of the highly endangered butterfly Coenonympha oedippus (Nymphalidae: Satyrinae) at its southern edge of distribution. Genetica 2021, 149, 21–36. [Google Scholar] [CrossRef]
- Zhang, D.-X.; Hewitt, G.M. Nuclear DNA analyses in genetic studies of populations: Practice, problems and prospects. Mol. Ecol. 2003, 12, 563–584. [Google Scholar] [CrossRef]
- Mackintosh, A.; Laetsch, D.R.; Hayward, A.; Charlesworth, B.; Waterfall, M.; Vila, R.; Lohse, K. The determinants of genetic diversity in butterflies. Nat. Commun. 2019, 10, 3466. [Google Scholar] [CrossRef]
- Robinson, J.A.; Ortega-Del Vecchyo, D.; Fan, Z.; Kim, B.Y.; vonHoldt, B.M.; Marsden, C.D.; Lohmueller, K.E.; Wayne, R.K. Genomic Flatlining in the Endangered Island Fox. Curr. Biol. 2016, 26, 1183–1189. [Google Scholar] [CrossRef] [PubMed]
- Dussex, N.; van der Valk, T.; Morales, H.E.; Wheat, C.W.; Díez-Del-Molino, D.; von Seth, J.; Foster, Y.; Kutschera, V.E.; Guschanski, K.; Rhie, A.; et al. Population genomics of the critically endangered kākāpō. Cell Genom. 2021, 1, 100002. [Google Scholar] [CrossRef] [PubMed]
- Thomas, W.J.W.; Anthony, J.M.; Dobrowolski, M.P.; Krauss, S.L. Optimising the conservation of genetic diversity of the last remaining population of a critically endangered shrub. AoB Plants 2021, 13, plab005. [Google Scholar] [CrossRef] [PubMed]
- Rutkowski, R.; Jagołkowska, P.; Zawadzka, D.; Bogdanowicz, W. Impacts of forest fragmentation and post-glacial colonization on the distribution of genetic diversity in the Polish population of the hazel grouse Terastes bonasia. Eur. J. Wildl. Res. 2016, 62, 293–306. [Google Scholar] [CrossRef]
- Andersen, J.C.; Havill, N.P.; Caccone, A.; Elkinton, J.S. Postglacial recolonization shaped the genetic diversity of the winter moth (Operophtera brumata) in Europe. Ecol. Evol. 2017, 7, 3312–3323. [Google Scholar] [CrossRef]
- Maresova, J.; Habel, J.C.; Neve, G.; Sielezniew, M.; Bartonova, A.; Kostro-Ambroziak, A.; Fric, Z.F. Cross-continental phylogeography of two Holarctic Nymphalid butterflies, Boloria eunomia and Boloria selene. PLoS ONE 2019, 14, e0214483. [Google Scholar] [CrossRef]
- Niedziałkowska, M.; Tarnowska, E.; Babik, W.; Konczal, M.; Gharbi, K.; Cezard, T.; Jędrzejewska, B. Different waves of postglacial recolonisation and genomic structure of bank vole populations in NE Poland. Heredity 2023, 130, 269–277. [Google Scholar] [CrossRef]
- Schwenk, K.; Brede, N.; Streit, B. Introduction. Extent, processes and evolutionary impact of interspecific hybridization in animals. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 2805–2811. [Google Scholar] [CrossRef]
- Jangjoo, M.; Matter, S.F.; Roland, J.; Keyghobadi, N. Connectivity rescues genetic diversity after a demographic bottleneck in a butterfly population network. Proc. Natl. Acad. Sci. USA 2016, 113, 10914–10919. [Google Scholar] [CrossRef]
- Schmitt, T.; Hewitt, G.M. The genetic pattern of population threat and loss: A case study of butterflies. Mol. Ecol. 2004, 13, 21–31. [Google Scholar] [CrossRef]
- Habel, J.C.; Husemann, M.; Finger, A.; Danley, P.D.; Zachos, F.E. The relevance of time series in molecular ecology and conservation biology. Biol. Rev. 2014, 89, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Sielezniew, M.; Deoniziak, K.; Dziekańska, I.; Nowicki, P. Dispersal in a metapopulation of the critically endangered Danube Clouded Yellow butterfly Colias myrmidone: Implications for conservation. J. Insect Conserv. 2019, 23, 291–300. [Google Scholar] [CrossRef]
- Duffy, E.; Archer, C.R.; Sharma, M.D.; Prus, M.; Joag, R.A.; Radwan, J.; Wedell, N.; Hosken, D.J. Wolbachia infection can bias estimates of intralocus sexual conflict. Ecol. Evol. 2019, 9, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Shapoval, N.A.; Kir’yanov, A.V.; Krupitsky, A.V.; Yakovlev, R.V.; Romanovich, A.E.; Zhang, J.; Grishin, N.V.; Kovalenko, M.G.; Shapoval, G.N. Phylogeography of Two Enigmatic Sulphur Butterflies, Colias mongola Alphéraky, 1897 and Colias tamerlana Staudinger, 1897 (Lepidoptera, Pieridae), with Relations to Wolbachia Infection. Insects 2023, 14, 943. [Google Scholar] [CrossRef]
- Dzurinka, M.; Šemeláková, M.; Panigaj, L. Taxonomy of hybridizing Colias croceus (Geoffroy, 1785) and Colias erate (Esper, 1805) (Lepidoptera, Pieridae) in light of mitochondrial and nuclear DNA, with occurrence and effects of Wolbachia infection. Zool. Anz. 2022, 299, 73–811. [Google Scholar] [CrossRef]
- Funkhouser-Jones, L.J.; Sehnert, S.R.; Martínez-Rodríguez, P.; Toribio-Fernández, R.; Pita, M.; Bella, J.L.; Bordenstein, S.R. Wolbachia co-infection in a hybrid zone: Discovery of horizontal gene transfers from two Wolbachia supergroups into an animal genome. PeerJ 2015, 7, e1479. [Google Scholar] [CrossRef]
- Kemp, D.J.; Thomson, F.E.; Edwards, W.; Iturbe-Ormaetxe, I. Incomplete offspring sex bias in Australian populations of the butterfly Eurema hecabe. Heredity 2017, 118, 284–292. [Google Scholar] [CrossRef]
- Narita, S.; Nomura, M.; Kageyama, D. A natural population of the butterfly Eurema hecabe with Wolbachia-induced female-biased sex ratio not by feminization. Genome 2007, 50, 365–372. [Google Scholar] [CrossRef]
- Charlat, S.; Hornett, E.A.; Dyson, E.A.; Ho, P.P.Y.; Loc, N.T.; Schilthuizen, M.; Davies, N.; Roderick, G.K.; Hurst, G.D.D. Prevalence and penetrance variation of male-killing Wolbachia across Indo-Pacific populations of the butterfly Hypolimnas bolina. Mol. Ecol. 2005, 14, 3525–3530. [Google Scholar] [CrossRef]
- Kern, P.; Cook, J.M.; Kageyama, D.; Riegler, M. Double trouble: Combined action of meiotic drive and Wolbachia feminization in Eurema butterflies. Biol. Lett. 2015, 11, 20150095. [Google Scholar] [CrossRef]
- Kageyama, D.; Ohno, M.; Sasaki, T.; Yoshido, A.; Konagaya, T.; Jouraku, A.; Kuwazaki, S.; Kanamori, H.; Katayose, Y.; Narita, S.; et al. Feminizing Wolbachia endosymbiont disrupts maternal sex chromosome inheritance in a butterfly species. Evol. Lett. 2017, 1, 232–244. [Google Scholar] [CrossRef]
- Després, L.; Henniaux, C.; Rioux, D.; Capblancq, T.; Zupan, S.; Čelik, T.; Sielezniew, M.; Bonato, L.; Ficetola, G.F. Inferring the biogeography and demographic history of an endangered butterfly in Europe from multilocus markers. Biol. J. Linn. Soc. 2019, 126, 95–113. [Google Scholar] [CrossRef]
- Toews, D.P.; Mandic, M.; Richards, J.G.; Irwin, D.E. Migration, mitochondria, and the yellow-rumped warbler. Evolution 2014, 68, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Bazin, E.; Glémin, S.; Galtier, N. Population size does not influence mitochondrial genetic diversity in animals. Science 2006, 312, 570–572. [Google Scholar] [CrossRef] [PubMed]
- Narita, S.; Shimajiri, Y.; Nomura, M. Strong cytoplasmic incompatibility and high vertical transmission rate can explain the high frequencies of Wolbachia infection in Japanese populations of Colias erate poliographus (Lepidoptera: Pieridae). Bull. Entom. Res. 2009, 99, 385–391. [Google Scholar] [CrossRef]
- Schoville, S.D.; Stuckey, M.; Roderick, G.K. Pleistocene origin and population history of a neoendemic alpine butterfly. Mol. Ecol. 2011, 20, 1233–1247. [Google Scholar] [CrossRef]
- Karaçetin, E.; Welch, H.J. Red Book of Butterflies in Turkey; Nature Conservation Center: Ankara, Turkey, 2011. [Google Scholar]
- Nekrutenko, Y.P. Butterflies of the Caucasus: Keys to Their Identification (Papilionidae, Pieridae, Satyridae, Danidae); Naukova Dumka: Kiev, Ukraine, 1990. [Google Scholar]
- Pamperis, L.N. The Butterflies of Greece; Editions Pamperis: Athens, Greece, 2009. [Google Scholar]
- Verovnik, R.; Micevski, B.; Đurić, M.; Jakšić, P.; Keymeulen, A.; Van Swaay, C.; Veling, K. Contribution to the knowledge of the butterfly fauna of the Republic of Macedonia (Lepidoptera: Papilionoidea & Hesperioidea). Acta Entomol. Slov. 2010, 18, 31–46. [Google Scholar] [CrossRef]
- Tvrtković, N.; Mihoci, I.; Šašić, M. Colias caucasica balcanica Rebel, 1901 (Pieridae) u Hrvatskoj–najzapadnija točka rasprostranjenja. Nat. Croat. Period. Musei Hist. Nat. Croat. 2011, 20, 375–385. [Google Scholar]
- Nahirnić, A.; Jakšić, P.; Viborg, A.L. Colias caucasica balcanica (Pieridae) rediscovered in Montenegro, with additional new records for Serbia. Phegea 2015, 43, 6–10. [Google Scholar]
- Tot, I.S.; Slacki, A.; Đurić, M.; Popović, M. Butterflies of the Vlasina region in southeast Serbia (Lepidoptera: Papilionoidea). Acta Entomol. Serbica 2015, 20, 117–135. [Google Scholar]
- Hristova, H.O.; Beshkov, S. Checklist of the superfamilies Hesperioidea and Papilionoidea (Insecta: Lepidoptera) of Bulgaria, with application of the IUCN Red List criteria at national level. Acta Zool. Bulg. 2017, 69, 105–114. [Google Scholar]
- Cuvelier, S.; Parmentier, L.; Paparisto, A.; Couckuyt, J. Butterflies of Albania–Fluturat e Shqipërisë: New surveys, new species and a new checklist (Lepidoptera: Papilionoidea). Phegea 2018, 46, 48–69. [Google Scholar] [CrossRef]
- Wiemers, M.; Balletto, E.; Dincă, V.; Fric, Z.F.; Lamas, G.; Lukhtanov, V.; Munguira, M.L.; van Swaay, C.A.M.; Vila, R.; Vliegenthart, A.; et al. An updated checklist of the European butterflies (Lepidoptera, Papilionoidea). ZooKeys 2018, 811, 9. [Google Scholar] [CrossRef][Green Version]
- Wagener, S. Colias caucasica balcanica Rebel, 1901 (comb. nov.; stat. nov.)(Lepidoptera: Pieridae). Phegea 1990, 18, 59–63. [Google Scholar]
- Tolman, T.; Lewington, R. Collins Butterfly Guide: The Most Complete Field Guide to the Butterflies of Britain and Europe; Harper Collins: London, UK, 2009. [Google Scholar]
- Stella, D.; Fric, F.Z.; Rindoš, M.; Kleisner, K.; Pecháček, P. Distribution of ultraviolet ornaments in Colias butterflies (Lepidoptera: Pieridae). Environ. Entomol. 2018, 47, 1344–1354. [Google Scholar] [CrossRef]
- Grieshuber, J.; Worthy, B.; Lamas, G. The Genus Colias Fabricius, 1807. Jan Haugum’s Annotated Catalogue of the Old World Colias (Lepidoptera, Pieridae); Tshikolovets Publications: Pardubice, Czech Republic, 2012. [Google Scholar]
- Grieshuber, J. Guide to the Butterflies of the Palearctic Region; Pieridae Part II; Omnes Artes: Milano, Italy, 2014. [Google Scholar]
- Grieshuber, J.; Lamas, G. A synonymic list of the genus Colias Fabricius, 1807 (Lepidoptera: Pieridae). Mitt. Münch. Ent. Ges. 2007, 97, 131–171. [Google Scholar]
- Gaunet, A.; Dincă, V.; Dapporto, L.; Montagud, S.; Vodă, R.; Schär, S.; Badiane, A.; Font, E.; Vila, R. Two consecutive Wolbachia-mediated mitochondrial introgressions obscure taxonomy in Palearctic swallowtail butterflies (Lepidoptera, Papilionidae). Zool. Scr. 2019, 48, 507–519. [Google Scholar] [CrossRef]
- Miyata, M.N.; Nomura, M.; Kageyama, D. Wolbachia have made it twice: Hybrid introgression between two sister species of Eurema butterflies. Ecol. Evol. 2020, 10, 8323–8330. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).