
Seasonal Dynamics and Damage of Halyomorpha halys in Italian Vineyards

 ,
, 
 , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Seasonal Dynamics of H. halys in the Vineyards
2.2. Assessment of H. halys Damage to the Grape Cluster
2.3. Fungicide and Insecticide Application on H. halys Infestation and Grey Mold and Sour Rot Infection
2.4. Statistical Analyses
3. Results
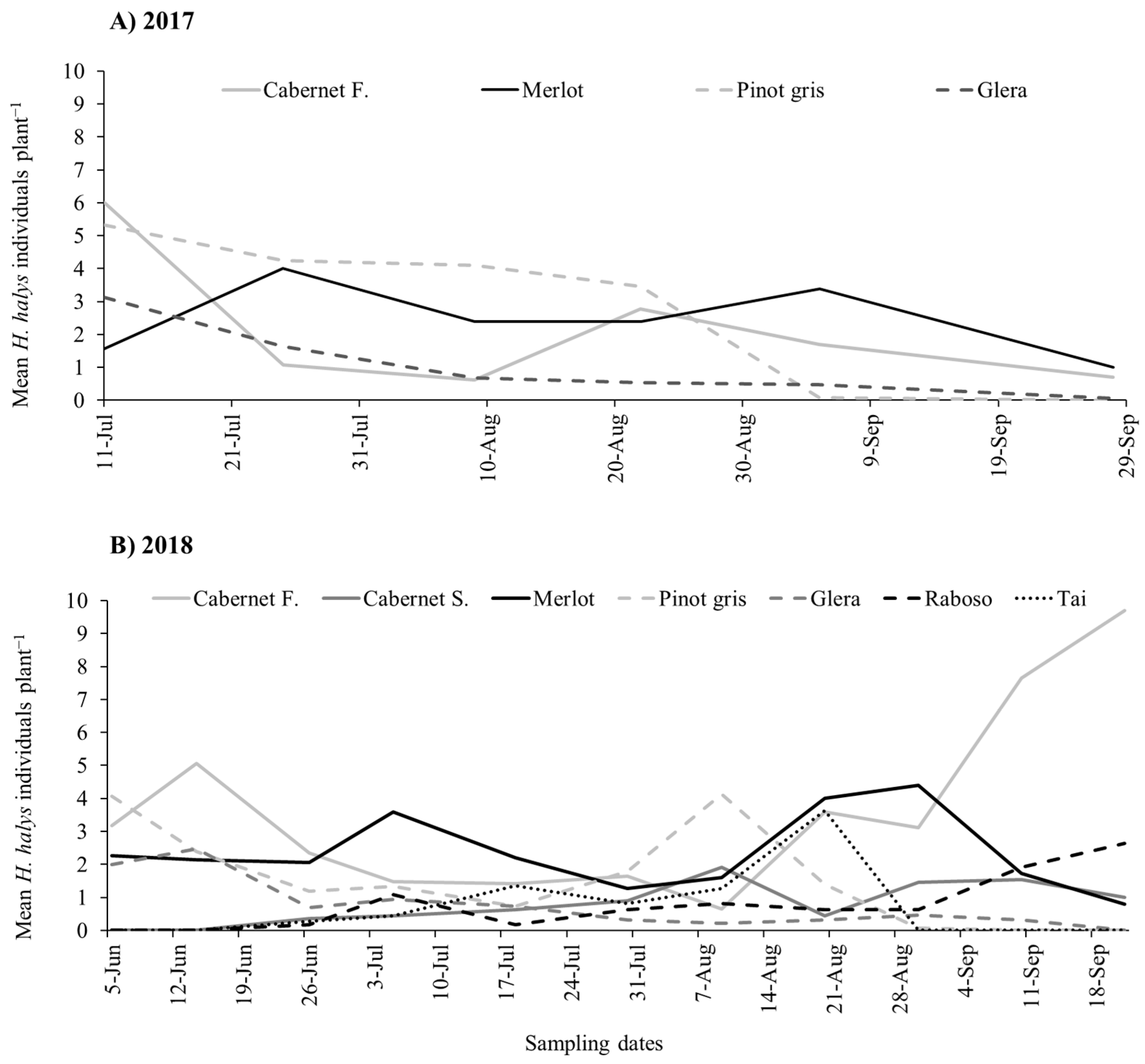
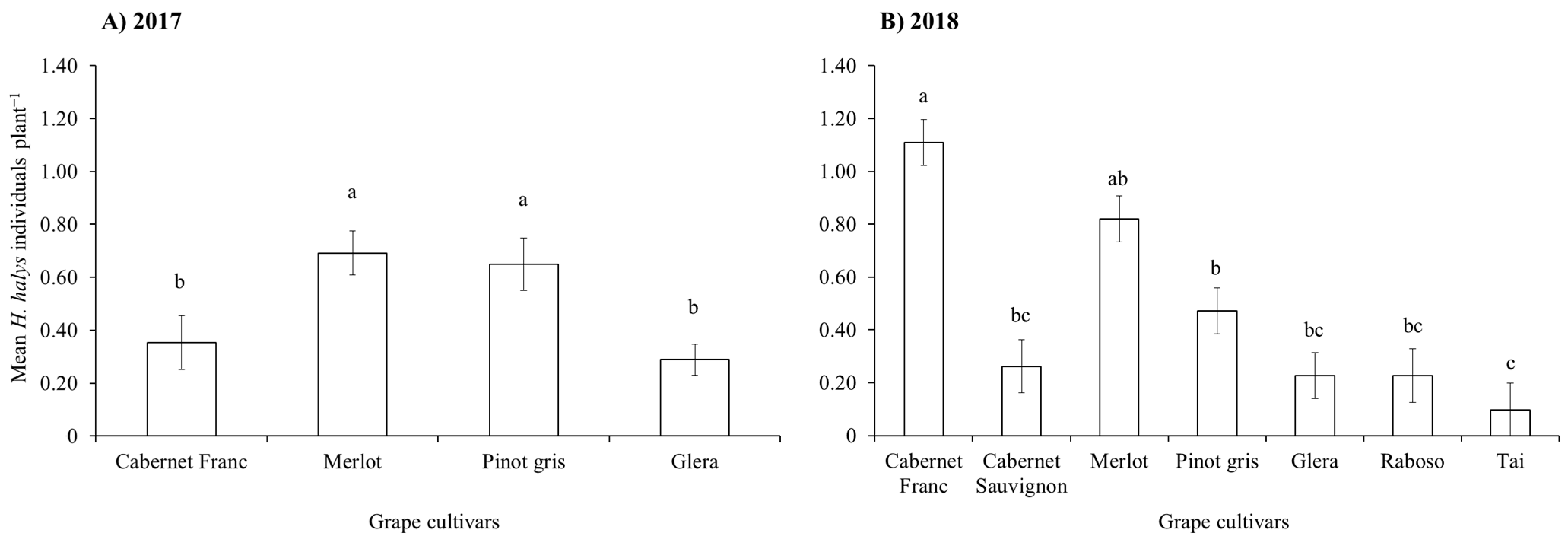
3.1. Seasonal Dynamics of H. halys in the Vineyards
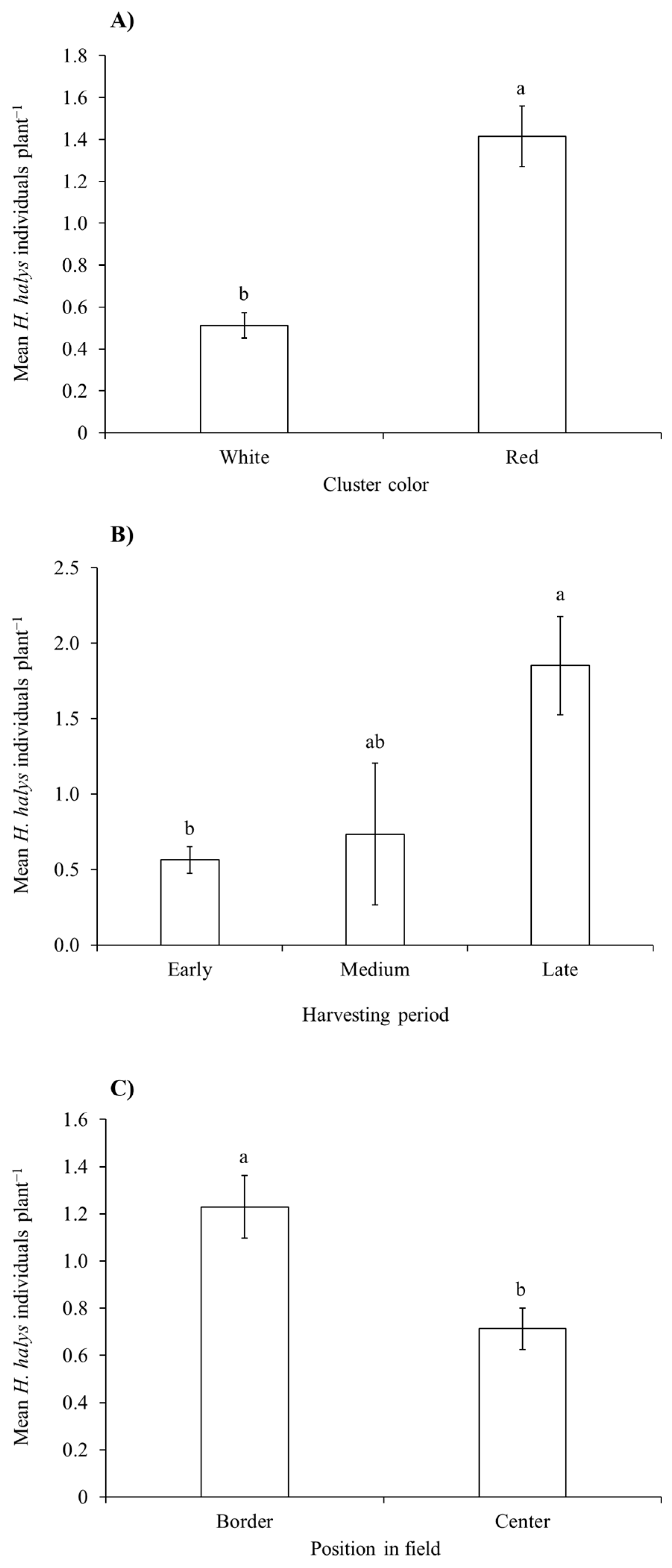
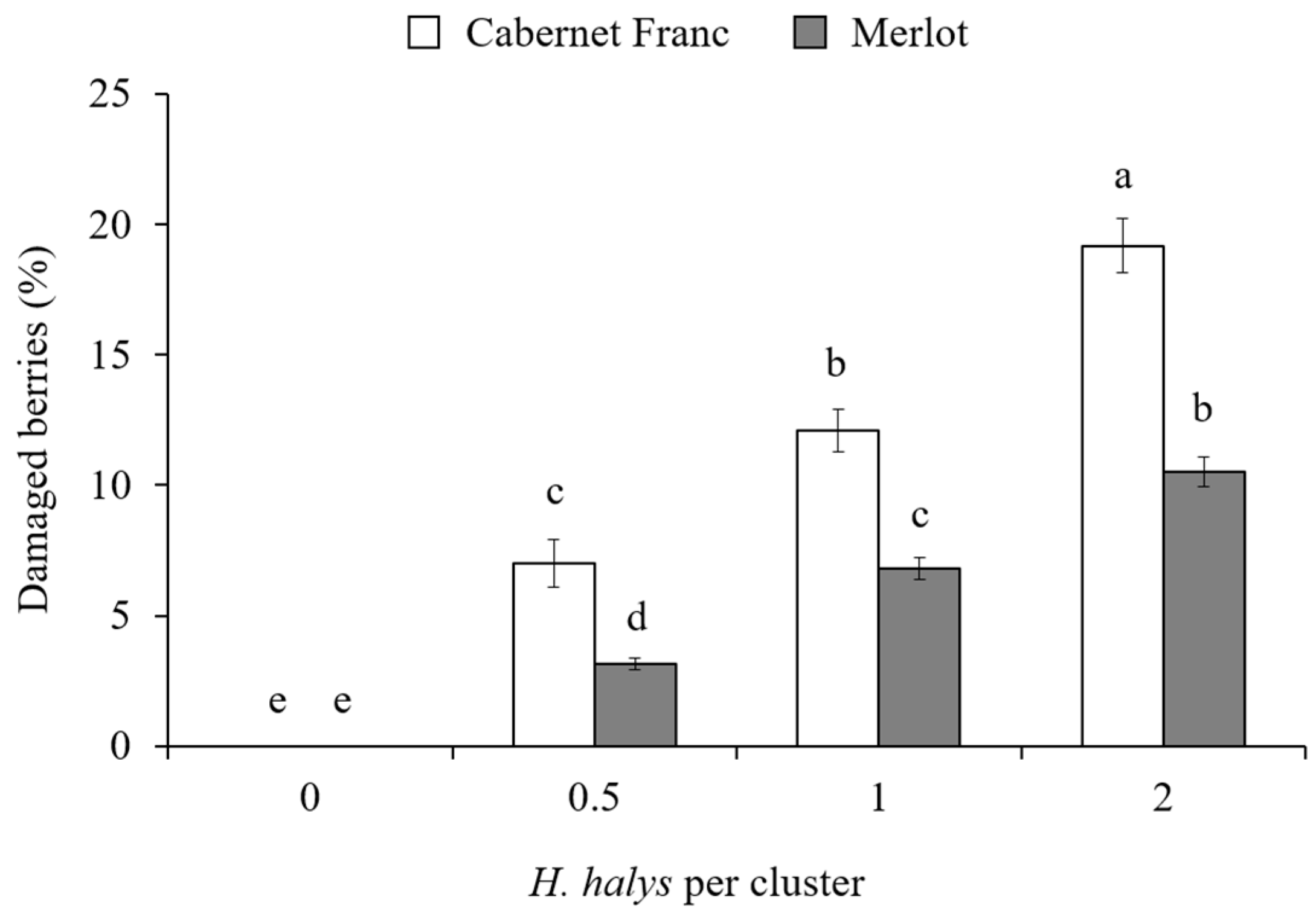
3.2. Assessment of H. halys Damage to the Grape Cluster
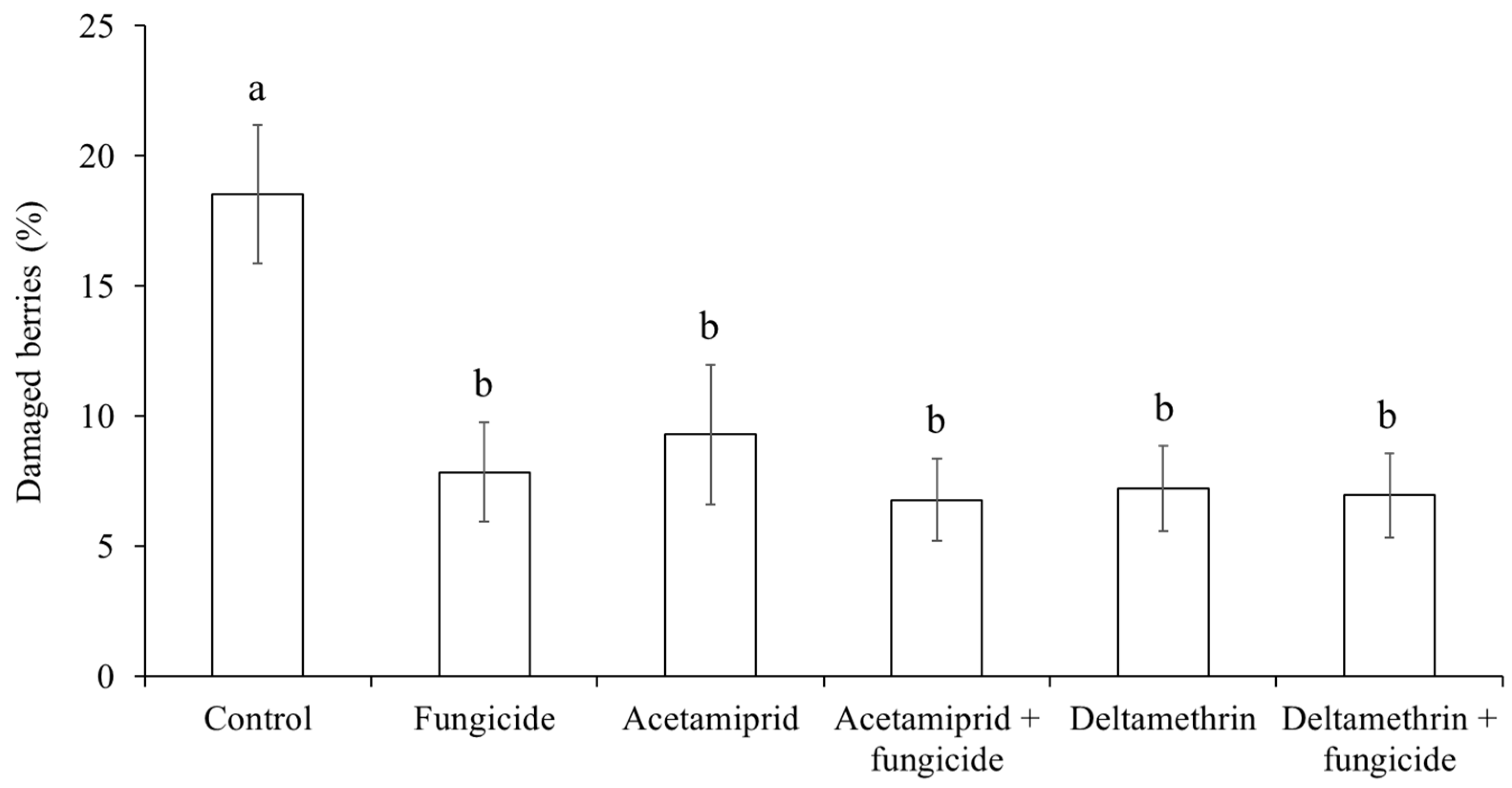
3.3. Fungicide and Insecticide Application on H. halys Infestation and Grey Mold and Sour Rot Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- OIV. State of the World Wine and Wine Sector in 2022. Available online: https://www.oiv.int/sites/default/files/documents/OIV_State_of_the_world_Vine_and_Wine_sector_in_2022_2.pdf (accessed on 11 January 2024).
- Fregoni, M. Viticoltura di Qualità; Edizioni L’Informatore Agrario S.r.l.: Piacenza, Italy, 1998. [Google Scholar]
- Raimondi, S. Vite e vino nel mondo. In La Vite e il Vino; Angelini, R., Ed.; Coltura e Cultura, Bayer CropScience S.r.l. and ART Servizi Editoriali S.p.A.: Milan and Bologna, Italy, 2007; pp. 548–579. [Google Scholar]
- IWC. Winegrape Production by Region. Available online: https://italianwinecentral.com/grape-production-by-region/ (accessed on 11 January 2024).
- Leskey, T.C.; Nielsen, A.L. Impact of The Invasive Brown Marmorated Stink Bug in North America and Europe: History, Biology, Ecology, and Management. Annu. Rev. Entomol. 2018, 63, 599–618. [Google Scholar] [CrossRef] [PubMed]
- Maistrello, L.; Dioli, P.; Bariselli, M.; Mazzoli, G.L.; Giacalone-Forini, I. Citizen Science and Early Detection of Invasive Species: Phenology of First Occurrences of Halyomorpha halys in Southern Europe. Biol. Invasions 2016, 18, 3109–3116. [Google Scholar] [CrossRef]
- Delbac, L.; Rouzes, R.; Hamidi, R.; Thiéry, D. First Occurrence of Halyomorpha halys in Bordeaux Vineyards. OENO One 2022, 56, 253–257. [Google Scholar] [CrossRef]
- Pajač Živković, I.; Čirjak, V.; Miklečić, I.; Pintar, M.; Duralija, B.; Lemic, D. First Evidence of The Brown Marmorated Stink Bug and its Population Size in Perennial Crops in Croatia. J. Cent. Eur. Agric. 2023, 24, 908–915. [Google Scholar] [CrossRef]
- Basnet, S.; Kuhar, T.P.; Laub, C.A.; Pfeiffer, D.G. Seasonality and Distribution Pattern of Brown Marmorated Stink Bug (Hemiptera: Pentatomidae) in Virginia Vineyards. J. Econom. Entomol. 2015, 108, 1902–1909. [Google Scholar] [CrossRef] [PubMed]
- Dingha, B.N.; Jackai, L.E.N. Laboratory Rearing of the Brown Marmorated Stink Bug (Hemiptera: Pentatomidae) and the Impact of Single and Combination of Food Substrates on Development and Survival. Can. Entomol. 2017, 149, 104–117. [Google Scholar] [CrossRef]
- Stahl, J.M.; Scaccini, D.; Daane, K.M. Field Survival Of The Brown Marmorated Stink Bug Halyomorpha halys (Hemiptera: Pentatomidae) on California Tree Crops. Environ. Entomol. 2021, 50, 1187–1193. [Google Scholar] [CrossRef] [PubMed]
- Bettiga, L.J.; Varela, L.G.; Bentley, W.J. Brown Marmorated Stink Bug. In Grape Pest Management, 3rd ed.; Bettiga, L.J., Ed.; University of California–Agriculture and Natural Resources: Berkeley, CA, USA, 2013; pp. 187–188. [Google Scholar]
- Costi, E.; Haye, T.; Maistrello, L. Biological parameters of the invasive brown marmorated stink bug, Halyomorpha halys, in Southern Europe. J. Pest Sci. 2017, 90, 1059–1067. [Google Scholar] [CrossRef]
- Lee, D.-H.; Leskey, T.C. Flight Behavior of Foraging and Overwintering Brown Marmorated Stink Bug, Halyomorpha halys (Hemiptera: Pentatomidae). B. Entomol. Res. 2015, 105, 566–573. [Google Scholar] [CrossRef]
- Kirkpatrick, D.M.; Rice, K.B.; Ibrahim, A.; Fleischer, S.J.; Tooker, J.F.; Tabb, A.; Medeiros, H.; Morrison III, W.R.; Leskey, T.C. The Influence of Marking Methods on Mobility, Survivorship, and Field Recovery of Halyomorpha halys (Hemiptera: Pentatomidae) Adults and Nymphs. Environ. Entomol. 2020, 49, 1026–1031. [Google Scholar] [CrossRef]
- Lee, D.-H.; Nielsen, A.L.; Leskey, T.C. Dispersal Capacity and Behavior of Nymphal Stages of Halyomorpha halys (Hemiptera: Pentatomidae) Evaluated Under Laboratory and Field Conditions. J. Insect Behav. 2014, 27, 639–651. [Google Scholar] [CrossRef]
- Acebes-Doria, A.L.; Leskey, T.C.; Bergh, J.C. Host Plant Effects on Halyomorpha halys (Hemiptera: Pentatomidae) Nymphal Development and Survivorship. Environ. Entomol. 2016, 45, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Wallner, A.M.; Hamilton, G.C.; Nielsen, A.L.; Hahn, N.; Green, E.J.; Rodriguez-Saona, C.R. Landscape Factors Facilitating the Invasive Dynamics and Distribution of the Brown Marmorated Stink Bug, Halyomorpha halys (Hemiptera: Pentatomidae), after Arrival in the United States. PLoS ONE 2014, 9, e95691. [Google Scholar] [CrossRef]
- Bakken, A.J.; Schoof, S.C.; Bickerton, M.; Kamminga, K.L.; Jenrette, J.C.; Malone, S.; Abney, M.A.; Herbert, D.A.; Reisig, D.; Kuhar, T.P.; et al. Occurrence of Brown Marmorated Stink Bug (Hemiptera: Pentatomidae) on Wild Hosts in Nonmanaged Woodlands and Soybean Fields in North Carolina and Virginia. Environ. Entomol. 2015, 44, 1011–1021. [Google Scholar] [CrossRef] [PubMed]
- Laterza, I.; Sabree, Z.L.; Martinez-Sañudo, I.; Scaccini, D.; Pozzebon, A.; Cornara, D.; de Lillo, E.; Tamburini, G. Halyomorpha halys in Mediterranean Areas: Local and Landscape Predictors, Genetic Diversity, and Potential Biological Control. Entomol. Gen. 2023, 43, 981–990. [Google Scholar] [CrossRef]
- Tamburini, G.; Laterza, I.; Nardi, D.; Mele, A.; Mori, N.; Pasini, M.; Scaccini, D.; Pozzebon, A.; Marini, L. Effect of Landscape Composition on the Invasive Pest Halyomorpha halys in Fruit Orchards. Agric. Ecosyst. Environ. 2023, 353, 108530. [Google Scholar] [CrossRef]
- Park, Y.-L.; Choi, K.; Cullum, J.; Hoelmer, K.A.; Weber, D.C.; Morrison, W.R., III; Rice, K.B.; Krawczyk, G.; Fleischer, S.J.; Hamilton, G.; et al. Landscape-Scale Spatiotemporal Dynamics of Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) Populations: Implications for Spatially-Based Pest Management. Pest Manag. Sci. 2024, 80, 953–966. [Google Scholar] [CrossRef] [PubMed]
- Leskey, T.C.; Hamilton, G.C.; Nielsen, A.L.; Polk, D.F.; Rodriguez-Saona, C.; Bergh, J.C.; Herbert, D.A.; Kuhar, T.P.; Pfeiffer, D.; Dively, G.P.; et al. Pest Status of the Brown Marmorated Stink Bug, Halyomorpha halys in the USA. Outlooks Pest Manag. 2012, 23, 218–226. [Google Scholar] [CrossRef]
- Kamminga, K.; Herbert, D.A.; Toews, M.D.; Malone, S.; Kuhar, T. Halyomorpha halys (Hemiptera: Pentatomidae) Feeding Injury on Cotton Bolls. J. Cotton Sci. 2014, 18, 68–74. [Google Scholar]
- Rice, K.B.; Bergh, C.J.; Bergmann, E.J.; Biddinger, D.J.; Dieckhoff, C.; Dively, G.; Fraser, H.; Gariepy, T.; Hamilton, G.; Haye, T.; et al. Biology, Ecology, and Management of Brown Marmorated Stink Bug (Hemiptera: Pentatomidae). J. Integr. Pest Manag. 2014, 5, 1–13. [Google Scholar] [CrossRef]
- Moore, L.C.; Tirello, P.; Scaccini, D.; Toews, M.D.; Duso, C.; Pozzebon, A. Characterizing Damage Potential of the Brown Marmorated Stink Bug (Hemiptera: Pentatomidae) in Cherry Orchards. Entomol. Gen. 2019, 39, 271–283. [Google Scholar] [CrossRef]
- Stahl, J.M.; Scaccini, D.; Pozzebon, A.; Daane, K.M. Comparing the Feeding Damage of the Invasive Brown Marmorated Stink Bug to a Native Stink Bug and Leaffooted Bug on California Pistachios. Insects 2020, 11, 688. [Google Scholar] [CrossRef] [PubMed]
- Martinson, H.M.; Raupp, M.J.; Shrewsbury, P.M. Invasive Stink Bug Wounds Trees, Liberates Sugars, and Facilitates Native Hymenoptera. Ann. Entomol. Soc. Am. 2013, 106, 47–52. [Google Scholar] [CrossRef]
- Scaccini, D.; Pozzebon, A. Invasive Brown Marmorated Stink Bug (Hemiptera: Pentatomidae) Facilitates Feeding of European Wasps and Ants (Hymenoptera: Vespidae, Formicidae) on Plant Exudates. Eur. J. Entomol. 2021, 118, 24–30. [Google Scholar] [CrossRef]
- Hori, K. Possible Causes of Disease Symptoms Resulting from the Feeding of Phytophagous Heteroptera. In Heteroptera of Economic Importance; Schaefer, C.W., Panizzi, A.R., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 11–35. [Google Scholar]
- Nielsen, A.L.; Rivera, M.; Polk, D.; Leskey, T.; Morrison, R.; Dalton, D.; Hedstrom, C.; Tomasino, E.; Walton, V.; Wiman, N.; et al. Integrated Pest Management for Brown Marmorated Stink Bug in Grapes. Available online: https://www.stopbmsb.org/managing-bmsb/management-by-crop/grapes/ (accessed on 11 January 2024).
- Mohekar, P. Brown Marmorated Stink Bug (BMSB), Halyomorpha halys Taint in Wine: Impact on Wine Sensory, Effect of Wine-processing and Management Techniques. Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 2016. [Google Scholar]
- Mohekar, P.; Lapis, T.J.; Wiman, N.G.; Lim, J.; Tomasino, E. Brown Marmorated Stink Bug Taint in Pinot Noir: Detection and Consumer Rejection Thresholds of Trans-2-Decenal. Am. J. Enol. Viticult. 2017, 68, 120–126. [Google Scholar] [CrossRef]
- Mohekar, P.; Osborne, J.; Wiman, N.G.; Walton, W.; Tomasino, E. Influence of Winemaking Processing Steps on the Amounts of (E)-2-Decenal and Tridecane as Off-Odorants Caused by Brown Marmorated Stink Bug (Halyomorpha halys). J. Agric. Food Chem. 2017, 65, 872–878. [Google Scholar] [CrossRef]
- Mohekar, P.; Osborne, J.; Tomasino, E. Effects of Fining Agents, Reverse Osmosis and Wine Age on Brown Marmorated Stink Bug (Halyomorpha halys) Taint in Wine. Beverages 2018, 4, 17. [Google Scholar] [CrossRef]
- Kehrli, P.; Rösti, J.; Lorenzini, F.; Deneulin, P.; Linder, C. Influence of Processed Halyomorpha halys Bugs on the Aroma and Taste of ‘Chardonnay’ and ‘Merlot’ Musts and Wines. Vitis 2021, 60, 43–50. [Google Scholar]
- Smith, J.R.; Hesler, S.R.; Loeb, G.M. Potential Impact of Halyomorpha halys (Hemiptera: Pentatomidae) on Grape Production in the Finger Lakes Region of New York. J. Entomol. Sci. 2014, 49, 290–303. [Google Scholar] [CrossRef]
- Musolin, D.L.; Konjević, A.; Karpun, N.N.; Protsenko, V.Y.; Ayba, L.Y.; Saulich, A.K. Invasive Brown Marmorated Stink Bug Halyomorpha halys (Stål) (Heteroptera: Pentatomidae) in Russia, Abkhazia, and Serbia: History of Invasion, Range Expansion, Early Stages of Establishment, and First Records of Damage to Local Crops. Arthropod-Plant Inte. 2018, 12, 517–529. [Google Scholar] [CrossRef]
- IWC. Get into Italian Wine. Available online: https://italianwinecentral.com/region/veneto/ (accessed on 11 January 2024).
- Donèche, B. Carbohydrate Metabolism and Gluconic Acid Synthesis by Botrytis cinerea. Can. J. Bot. 1989, 67, 2888–2893. [Google Scholar] [CrossRef]
- Hong, Y.-S.; Martinez, A.; Liger-Belair, G.; Jeandet, P.; Nuzillard, J.-M.; Cilindre, C. Metabolomics Reveals Simultaneous Influences of Plant Defence System and Fungal Growth in Botrytis cinerea-Infected Vitis vinifera cv. Chardonnay Berries. J. Exp. Bot. 2012, 63, 5773–5785. [Google Scholar] [CrossRef] [PubMed]
- Albanese, D.; Malvano, F.; Sannini, A.; Pilloton, R.; Di Matteo, M. A Doped Polyaniline Modified Electrode Amperometric Biosensor for Gluconic Acid Determination in Grapes. Sensors 2014, 14, 11097–11109. [Google Scholar] [CrossRef]
- Cinquanta, L.; Albanese, D.; De Curtis, F.; Malvano, F.; Crescitelli, A.; Di Matteo, M. Rapid Assessment of Gray Mold (Botrytis cinerea) Infection in Grapes Using Biosensors System. Am. J. Enol. Viticult. 2015, 66, 502–508. [Google Scholar] [CrossRef]
- Kirchert, S.; Kaiserb, R.E.; Morlock, G.E. In-Process Quality Control of Wine by Planar Chromatography versus Micro Planar Chromatography. J. Chromatogr. A 2019, 1588, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Hill, G.N.; Evans, K.J.; Beresford, R.M.; Dambergs, R.G. Comparison of Methods for the Quantification of Botrytis Bunch Rot in White Wine Grapes. Aust. J. Grape Wine R. 2014, 20, 432–441. [Google Scholar] [CrossRef]
- SAS Institute. PROC User’s Manual, 6th ed.; SAS Institute: Cary, NC, USA, 2016; Available online: http://support.sas.com/documentation/cdl/en/indbug/68442/PDF/default/indbug.pdf (accessed on 11 January 2024).
- Basnet, S. Biology and Pest Status of Brown Marmorated Stink Bug (Hemiptera: Pentatomidae) in Virginia Vineyards and Raspberry Plantings. MSc Thesis, Virginia Polytechnic Institute and State University, Blacksburg, VA, USA, 2014. [Google Scholar]
- Bergmann, E.J.; Venugopal, P.D.; Martinson, H.M.; Raupp, M.J.; Shrewsbury, P.M. Host Plant use by the Invasive Halyomorpha halys (Stål) on Woody Ornamental Trees and Shrubs. PLoS ONE 2016, 11, e0149975. [Google Scholar] [CrossRef] [PubMed]
- Mensah-Bonsu, M.; Dingha, B.N.; Jackai, L.E.; Adjei-Fremah, S.; Worku, M. Evaluation of Preference of Brown Marmorated Stink Bug, Halyomorpha halys (Stål) for Different Colour Bell Peppers and the Role of Plant Protein. Arthropod-Plant Inte. 2020, 14, 363–372. [Google Scholar] [CrossRef]
- Griesser, M.; Weingart, G.; Schoedl-Hummel, K.; Neumann, N.; Becker, M.; Varmuza, K.; Liebner, F.; Schuhmacher, R.; Forneck, A. Severe Drought Stress is Affecting Selected Primary Metabolites, Polyphenols, and Volatile Metabolites in Grapevine Leaves (Vitis vinifera cv. Pinot noir). Plant Physiol. Bioch. 2015, 88, 17–26. [Google Scholar] [CrossRef]
- Midzi, J.; Jeffery, D.W.; Baumann, U.; Capone, D.L.; Rogiers, S.Y.; Pagay, V. Evidence of Bi-Directional Volatile-Mediated Communication between Drought-Stressed and Well-Watered Grapevines (Vitis vinifera L.). Agronomy 2023, 13, 1747. [Google Scholar] [CrossRef]
- Maistrello, L.; Costi, E.; Caruso, S.; Vaccari, G.; Bortolotti, P.; Nannini, R.; Casoli, L.; Montermini, A.; Bariselli, M.; Guidetti, R. Halyomorpha halys in Italy: First Results of Field Monitoring in Fruit Orchards. IOBC-WPRS Bull. 2016, 112, 1–5. [Google Scholar]
- Reeves, R.; Greene, J.; Reay-Jones, F.; Toews, M.; Gerard, P. Effects of Adjacent Habitat on Populations of Stink Bugs (Heteroptera: Pentatomidae) in Cotton as Part of a Variable Agricultural Landscape in South Carolina. Environ. Entomol. 2010, 39, 1420–1427. [Google Scholar] [CrossRef] [PubMed]
- Owens, D.R. Behavior of and Crop Injury Induced by Native and Exotic Stink Bugs in Mid-Atlantic Soybean. Master’s Thesis, Virginia Polytechnic Institute and State University, Blacksburg, VA, USA, 2012. [Google Scholar]
- Venugopal, P.D.; Coffey, P.L.; Dively, G.P.; Lamp, W.O. Adjacent Habitat Influence on Stink Bug (Hemiptera: Pentatomidae) Densities and the Associated Damage at Field Corn and Soybean Edges. PLoS ONE 2014, 9, e109917. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, P.D.; Dively, G.P.; Lamp, W.O. Spatiotemporal Dynamics of the Invasive Halyomorpha halys (Hemiptera: Pentatomidae) in and Between Adjacent Corn and Soybean Fields. J. Econ. Entomol. 2015, 108, 2231–2241. [Google Scholar] [CrossRef] [PubMed]
- Aigner, B.L.; Kuhar, T.P.; Herbert, D.A.; Brewster, C.C.; Hogue, J.W.; Aigner, J.D. Brown Marmorated Stink Bug (Hemiptera: Pentatomidae) Infestations in Tree Borders and Subsequent Patterns of Abundance in Soybean Fields. J. Econ. Entomol. 2017, 110, 487–490. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, P.D.; Martinson, H.M.; Bergmann, E.J.; Shrewsbury, P.M.; Raupp, M.J. Edge Effects Influence the Abundance of the Invasive Halyomorpha halys (Hemiptera: Pentatomidae) in Woody Plant Nurseries. Environ. Entomol. 2015, 44, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Bosco, L.; Moraglio, S.; Tavella, L. Halyomorpha halys, a Serious Threat for Hazelnut in Newly Invaded Areas. J. Pest Sci. 2018, 91, 661–670. [Google Scholar] [CrossRef]
- Blaauw, B.R.; Polk, D.; Nielsen, A.L. IPM-CPR for Peaches: Incorporating Behaviorally-Based Methods to Manage Halyomorpha halys and Key Pests in Peach. Pest Manag. Sci. 2015, 71, 1513–1522. [Google Scholar] [CrossRef]
- Bosco, L.; Nardelli, M.; Tavella, L. First Insights on Early Host Plants and Dispersal Behavior of Halyomorpha halys (Hemiptera: Pentatomidae) from Overwintering to Crop Colonization. Insects 2020, 11, 866. [Google Scholar] [CrossRef]
- Nielsen, A.L.; Hamilton, G.C. Life History of the Invasive Species Halyomorpha halys (Hemiptera: Pentatomidae) in Northeastern United States. Ann. Entomol. Soc. Am. 2009, 102, 608–616. [Google Scholar] [CrossRef]
- Martinson, H.M.; Venugopal, P.D.; Bergmann, E.J.; Shrewsbury, P.M.; Raupp, M.J. Fruit Availability Influences the Seasonal Abundance of Invasive Stink Bugs in Ornamental Tree Nurseries. J. Pest Sci. 2015, 88, 461–468. [Google Scholar] [CrossRef]
- Martinson, H.M.; Bergmann, E.J.; Venugopal, P.D.; Riley, C.B.; Shrewsbury, P.M.; Raupp, M.J. Invasive Stink Bug Favors Native Plants: Testing the Role of Plant Geographic Origin in Diverse, Managed Environments. Sci. Rep. 2016, 6, 32646. [Google Scholar] [CrossRef] [PubMed]
- Blaauw, B.R.; Jones, V.P.; Nielsen, A.L. Utilizing Immunomarking Techniques to Track Halyomorpha halys (Hemiptera: Pentatomidae) Movement and Distribution Within a Peach Orchard. PeerJ 2016, 4, e1997. [Google Scholar] [CrossRef] [PubMed]
- Lecompte, F.; Nicot, P.C.; Ripoll, J.; Abro, M.A.; Raimbault, A.K.; Lopez-Lauri, F.; Bertin, N. Reduced Susceptibility of Tomato Stem to the Necrotrophic Fungus Botrytis cinerea is Associated with a Specific Adjustment of Fructose Content in the Host Sugar Pool. Ann. Bot. 2017, 119, 931–943. [Google Scholar] [PubMed]
- Mitchell, P.L. Heteroptera as Vectors of Plant Pathogens. Neot. Entomol. 2004, 33, 519–545. [Google Scholar] [CrossRef]
- Mitchell, P.L.; Zeilinger, A.R.; Medrano, E.G.; Esquivel, J.F. Pentatomoids as Vectors of Plant Pathogens. In Invasive Stink Bugs and Related Species (Pentatomoidea) – Biology, Higher Systematics, Semiochemistry, and Management; McPherson, J.E., Ed.; CRC Press: Boca Raton, FL, USA, 2018; pp. 611–640. [Google Scholar]
- Brust, G.E.; Rane, K.K. Transmission of the Yeast Eremothecium coryli to Fruits and Vegetables by the Brown Marmorated Stink Bug. University of Maryland Extension. 2011. Available online: https://extension.umd.edu/learn/transmission-yeast-eremothecium-coryli-fruits-andvegetables-brown-marmorated-stink-bug (accessed on 11 January 2024).
- Dobson, R.C.; Rogers, M.; Moore, J.L.C.; Bessin, R.T. Exclusion of the Brown Marmorated Stink Bug from Organically Grown Peppers Using Barrier Screens. HortTechnology 2016, 26, 191–198. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Method | Source of Variation | df | F Value | p Value |
|---|---|---|---|---|
| Visual sampling (2017) | Sampling date (D) | 5, 444 | 0.24 | 0.9440 |
| Position (Pos) | 1, 427.7 | 0.99 | 0.3202 | |
| Cultivar (Cv) | 9, 427.8 | 0.91 | 0.5187 | |
| D × Pos | 5, 444.3 | 0.92 | 0.5812 | |
| D × Cv | 45, 444.2 | 1.01 | 0.4613 | |
| Pos × Cv | 5, 428.2 | 0.65 | 0.6621 | |
| D × Pos × Cv | 25, 444.5 | 0.92 | 0.5812 | |
| Beat sampling (2017) | D | 4, 216 | 3.94 | 0.0042 |
| Pos | 1, 106 | 0.06 | 0.8075 | |
| Cv | 3, 103 | 3.49 | 0.0185 | |
| D × Pos | 4, 216 | 0.46 | 0.7614 | |
| D × Cv | 12, 230 | 1.35 | 0.1903 | |
| Pos × Cv | 3, 103 | 0.89 | 0.4515 | |
| D × Pos × Cv | 12, 230 | 0.57 | 0.8663 | |
| Beat sampling (2018) | D | 10, 339 | 2.32 | 0.0119 |
| Pos | 1, 79.1 | 7.06 | 0.0096 | |
| Cv | 6, 79.1 | 7.36 | <0.0001 | |
| D × Pos | 10, 339 | 0.22 | 0.9944 | |
| D × Cv | 60, 345 | 1.81 | 0.0006 | |
| Pos × Cv | 6, 79.2 | 0.34 | 0.9152 | |
| D × Pos × Cv | 60, 345 | 0.56 | 0.9960 |
| Source of Variation | df | F Value | p Value |
|---|---|---|---|
| Red vs. White | 1, 79.1 | 10.54 | 0.0017 |
| Early vs. Medium | 1, 79.1 | 1.66 | 0.2008 |
| Early vs. Late | 1, 79.1 | 7.22 | 0.0088 |
| Medium vs. Late | 1, 79.1 | 2.83 | 0.0965 |
| Cultivar Category | Source of Variation | df | F Value | p Value |
|---|---|---|---|---|
| White cultivars | Cultivar (Cv) | 1, 440 | 52.50 | <0.0001 |
| Phenological stage (Phen) | 4, 440 | 8.75 | <0.0001 | |
| H. halys density (Dens) | 3, 440 | 319 | <0.0001 | |
| Cv × Phen | 4, 440 | 6.13 | <0.0001 | |
| Cv × Dens | 3, 440 | 18.6 | <0.0001 | |
| Phen × Dens | 12, 440 | 1.53 | 0.1094 | |
| Cv × Phen × Dens | 12, 440 | 2.08 | 0.0174 | |
| Red cultivars | Cv | 1, 440 | 149.30 | <0.0001 |
| Phen | 4, 440 | 2.85 | 0.0235 | |
| Dens | 3, 440 | 715.17 | <0.0001 | |
| Cv × Phen | 4, 440 | 1.00 | 0.4086 | |
| Cv × Dens | 3, 440 | 18.09 | <0.0001 | |
| Phen × Dens | 12, 440 | 1.18 | 0.2945 | |
| Cv × Phen × Dens | 12, 440 | 0.71 | 0.7378 |
| Cultivar Category | Source of Variation | df | F Value | p Value |
|---|---|---|---|---|
| White cultivars | Cultivar (Cv) | 1, 440 | 30.29 | <0.0001 |
| Phenological stage (Phen) | 4, 440 | 5.23 | 0.0004 | |
| H. halys density (Dens) | 3, 440 | 15.32 | <0.0001 | |
| Cv × Phen | 4, 440 | 0.57 | 0.6865 | |
| Cv × Dens | 3, 440 | 4.30 | 0.0053 | |
| Phen × Dens | 12, 440 | 1.54 | 0.1055 | |
| Cv × Phen × Dens | 12, 440 | 0.94 | 0.5113 | |
| Red cultivars | Cv | 1, 440 | 59.04 | <0.0001 |
| Phen | 4, 440 | 0.80 | 0.5232 | |
| Dens | 3, 440 | 1.17 | 0.3220 | |
| Cv × Phen | 4, 440 | 0.95 | 0.4367 | |
| Cv × Dens | 3, 440 | 0.80 | 0.4967 | |
| Phen × Dens | 12, 440 | 0.56 | 0.8752 | |
| Cv × Phen × Dens | 12, 440 | 0.70 | 0.7493 |
| Cultivar Category | Source of Variation | df | F Value | p Value |
|---|---|---|---|---|
| White cultivars | Cultivar (Cv) | 1, 440 | 343.27 | <0.0001 |
| Phenological stage (Phen) | 4, 440 | 1.41 | 0.2297 | |
| H. halys density (Dens) | 3, 440 | 1.17 | 0.3252 | |
| Cv × Phen | 4, 440 | 0.46 | 0.7132 | |
| Cv × Dens | 3, 440 | 0.46 | 0.7099 | |
| Phen × Dens | 12, 440 | 0.69 | 0.7628 | |
| Cv × Phen × Dens | 12, 440 | 0.98 | 0.4707 | |
| Red cultivars | Cv | 1, 440 | 13.42 | 0.0003 |
| Phen | 4, 440 | 1.79 | 0.1302 | |
| Dens | 3, 440 | 10.76 | <0.0001 | |
| Cv × Phen | 4, 440 | 2.66 | 0.0322 | |
| Cv × Dens | 3, 440 | 1.22 | 0.3016 | |
| Phen × Dens | 12, 440 | 1.02 | 0.4259 | |
| Cv × Phen × Dens | 12, 440 | 1.01 | 0.4349 |
| Sugar Content per Cultivar Category | Source of Variation | df | F Value | p Value |
|---|---|---|---|---|
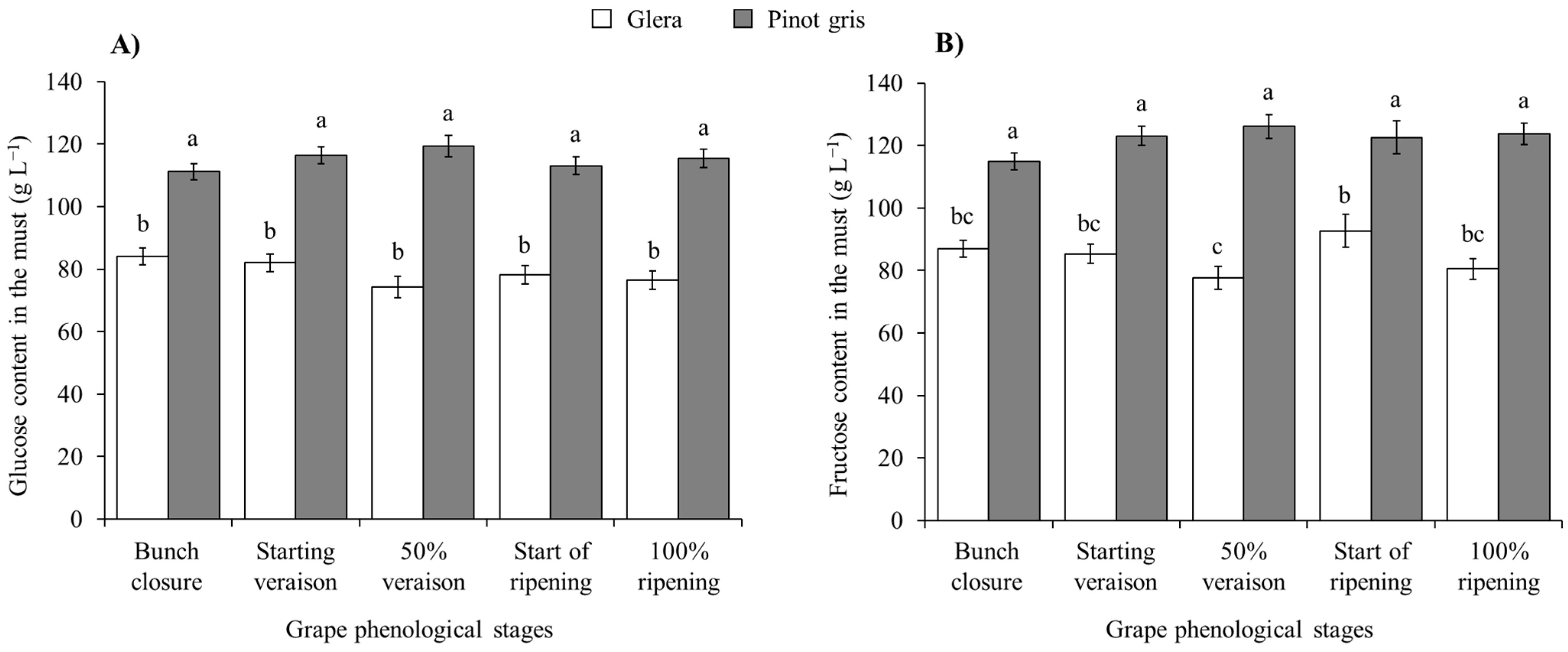
| Glucose (white cultivars) | Cultivar (Cv) | 1, 200 | 924.66 | <0.0001 |
| Phenological stage (Phen) | 4, 200 | 1.70 | 0.1515 | |
| H. halys density (Dens) | 3, 200 | 0.38 | 0.7661 | |
| Cv × Phen | 4, 200 | 6.88 | < 0.0001 | |
| Cv × Dens | 3, 200 | 0.11 | 0.9558 | |
| Phen × Dens | 12, 200 | 0.93 | 0.5225 | |
| Cv × Phen × Dens | 12, 200 | 1.24 | 0.2554 | |
| Glucose (red cultivars) | Cv | 1, 200 | 29.77 | <0.0001 |
| Phen | 4, 200 | 2.69 | 0.0321 | |
| Dens | 3, 200 | 0.88 | 0.4520 | |
| Cv × Phen | 4, 200 | 0.72 | 0.5797 | |
| Cv × Dens | 3, 200 | 1.75 | 0.1581 | |
| Phen × Dens | 12, 200 | 1.78 | 0.0541 | |
| Cv × Phen × Dens | 12, 200 | 1.39 | 0.1734 | |
| Fructose (white cultivars) | Cv | 1, 200 | 530.37 | <0.0001 |
| Phen | 4, 200 | 1.17 | 0.3263 | |
| Dens | 3, 200 | 3.37 | 0.0195 | |
| Cv × Phen | 4, 200 | 4.82 | 0.0010 | |
| Cv × Dens | 3, 200 | 1.31 | 0.2714 | |
| Phen × Dens | 12, 200 | 1.24 | 0.2596 | |
| Cv × Phen × Dens | 12, 200 | 1.63 | 0.0860 | |
| Fructose (red cultivars) | Cv | 1, 200 | 89.50 | < 0.0001 |
| Phen | 4, 200 | 3.30 | 0.0120 | |
| Dens | 3, 200 | 1.94 | 0.1237 | |
| Cv × Phen | 4, 200 | 0.72 | 0.5767 | |
| Cv × Dens | 3, 200 | 0.65 | 0.5832 | |
| Phen × Dens | 12, 200 | 2.16 | 0.0150 | |
| Cv × Phen × Dens | 12, 200 | 0.90 | 0.5462 |
| Acid Content per Cultivar Category | Source of Variation | df | F Value | p Value |
|---|---|---|---|---|
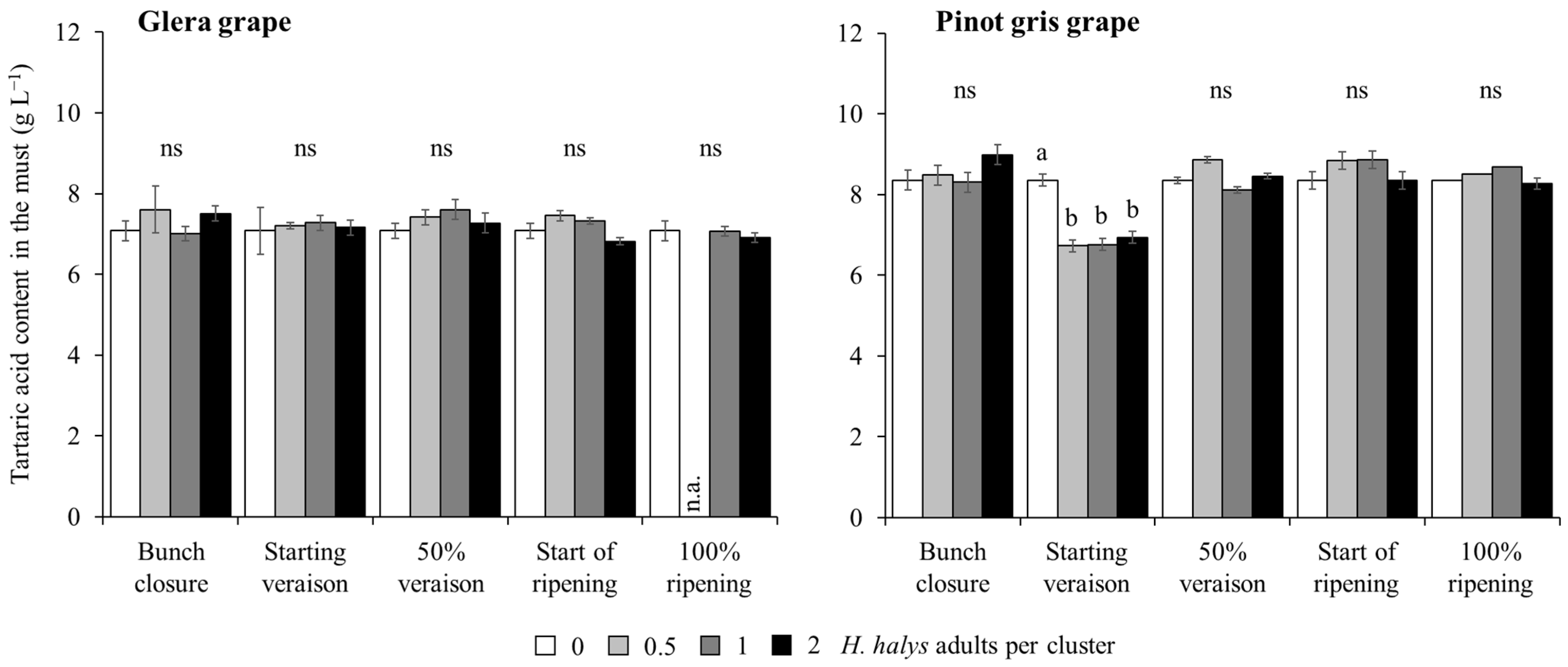
| Tartaric acid (white cultivars) | Cultivar (Cv) | 1, 195 | 182.05 | <0.0001 |
| Phenological stage (Phen) | 4, 195 | 16.03 | <0.0001 | |
| H. halys density (Dens) | 3, 195 | 1.73 | 0.1612 | |
| Cv × Phen | 4, 195 | 14.02 | <0.0001 | |
| Cv × Dens | 3, 195 | 3.23 | 0.0235 | |
| Phen × Dens | 12, 195 | 3.39 | 0.0002 | |
| Cv × Phen × Dens | 12, 195 | 1.95 | 0.0354 | |
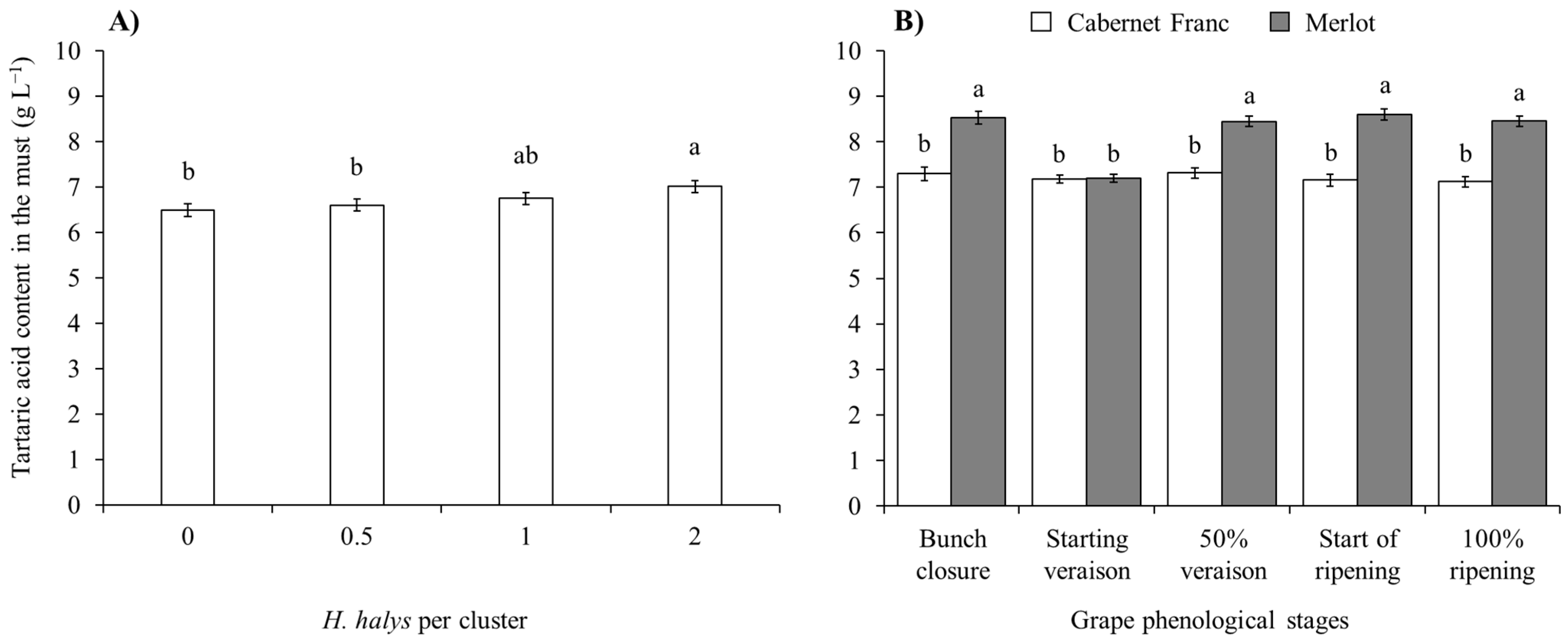
| Tartaric acid (red cultivars) | Cv | 1, 200 | 199.76 | <0.0001 |
| Phen | 4, 200 | 2.39 | 0.0523 | |
| Dens | 3, 200 | 5.95 | 0.0007 | |
| Cv × Phen | 4, 200 | 3.72 | 0.0061 | |
| Cv × Dens | 3, 200 | 0.75 | 0.5237 | |
| Phen × Dens | 12, 200 | 0.80 | 0.6509 | |
| Cv × Phen × Dens | 12, 200 | 1.01 | 0.4393 | |
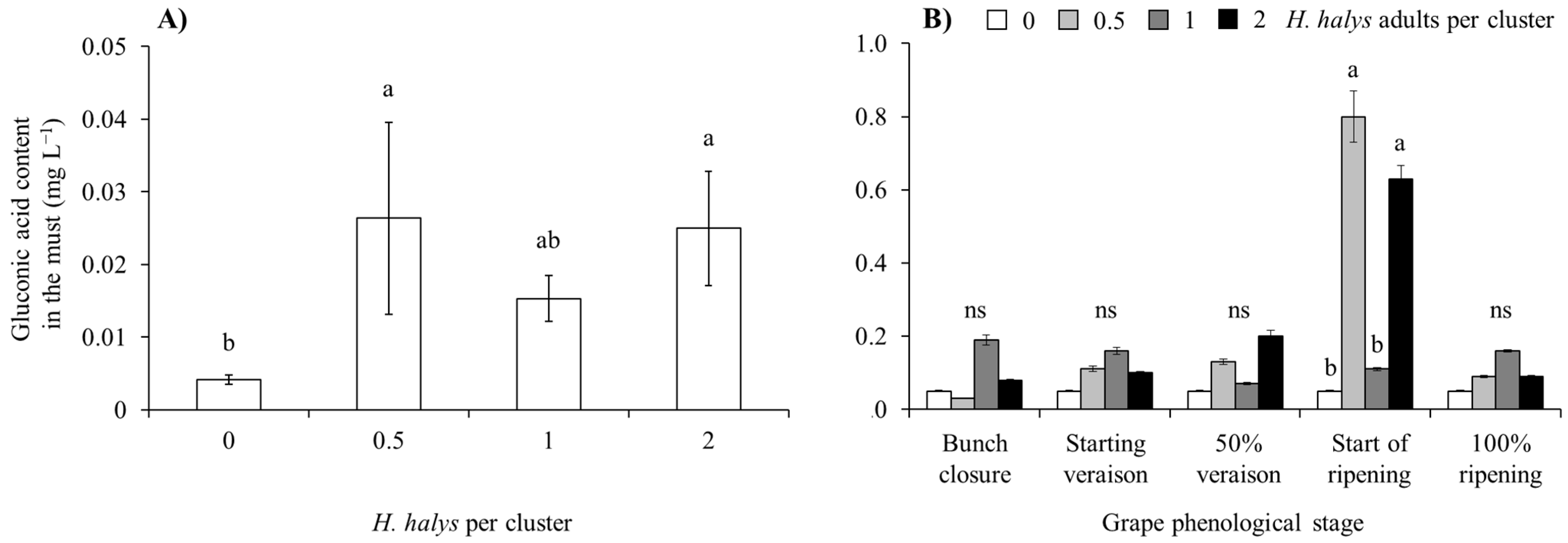
| Gluconic acid (white cultivars) | Cv | 1, 153 | 1.37 | 0.2439 |
| Phen | 4, 153 | 5.71 | 0.0003 | |
| Dens | 3, 153 | 3.63 | 0.0145 | |
| Cv × Phen | 4, 153 | 1.51 | 0.2007 | |
| Cv × Dens | 3, 153 | 0.71 | 0.5482 | |
| Phen × Dens | 12, 153 | 2.19 | 0.0149 | |
| Cv × Phen × Dens | 12, 153 | 0.70 | 0.7449 | |
| Gluconic acid (red cultivars) | Cv | 1, 157 | 0.44 | 0.5105 |
| Phen | 4, 157 | 3.38 | 0.0110 | |
| Dens | 3, 157 | 4.86 | 0.0029 | |
| Cv × Phen | 4, 157 | 1.77 | 0.1373 | |
| Cv × Dens | 3, 157 | 1.82 | 0.1454 | |
| Phen × Dens | 12, 157 | 1.47 | 0.1416 | |
| Cv × Phen × Dens | 12, 157 | 0.74 | 0.7109 |
| Source of Variation | df | F Value | p Value |
|---|---|---|---|
| Treatment | 5, 51.2 | 8.73 | <0.0001 |
| Sampling date (D) | 3, 117 | 81.41 | <0.0001 |
| Treatment × D | 15, 125 | 3.59 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scaccini, D.; Fornasiero, D.; Tirello, P.; Vincenzi, S.; Cecchetto, M.; Allgjata, I.; Duso, C.; Pozzebon, A. Seasonal Dynamics and Damage of Halyomorpha halys in Italian Vineyards. Insects 2024, 15, 378. https://doi.org/10.3390/insects15060378
Scaccini D, Fornasiero D, Tirello P, Vincenzi S, Cecchetto M, Allgjata I, Duso C, Pozzebon A. Seasonal Dynamics and Damage of Halyomorpha halys in Italian Vineyards. Insects. 2024; 15(6):378. https://doi.org/10.3390/insects15060378
Chicago/Turabian StyleScaccini, Davide, Diego Fornasiero, Paola Tirello, Simone Vincenzi, Massimo Cecchetto, Ilirjan Allgjata, Carlo Duso, and Alberto Pozzebon. 2024. "Seasonal Dynamics and Damage of Halyomorpha halys in Italian Vineyards" Insects 15, no. 6: 378. https://doi.org/10.3390/insects15060378
APA StyleScaccini, D., Fornasiero, D., Tirello, P., Vincenzi, S., Cecchetto, M., Allgjata, I., Duso, C., & Pozzebon, A. (2024). Seasonal Dynamics and Damage of Halyomorpha halys in Italian Vineyards. Insects, 15(6), 378. https://doi.org/10.3390/insects15060378