


Heterosis for Resistance to Insect Herbivores in a 3-Line Hybrid Rice System

 ,
,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Insect Herbivores
2.3. Heterosis for Resistance in Greenhouse Experiments
2.3.1. Planthopper Responses to Antibiosis of Hybrid and Parental Lines
2.3.2. Planthopper Oviposition Responses to Hybrid and Parental Lines
2.3.3. Stemborer Responses to Antibiosis of Hybrid and Parental Lines
2.3.4. Stemborer Oviposition Responses to Hybrid and Parental Lines
2.4. Field Screening of Hybrid and Parental Lines
2.4.1. Biomass Accumulation and Yields of Hybrid and Inbred Lines in Field Plots
2.4.2. Monitoring of Herbivores and Damage in Field Plots
2.5. Data Analyses
3. Results
3.1. Heterosis and Heterobeltiosis for Resistance in Greenhouse Experiments
3.1.1. BPH Resistance
3.1.2. WBPH Resistance
3.1.3. YSB Resistance
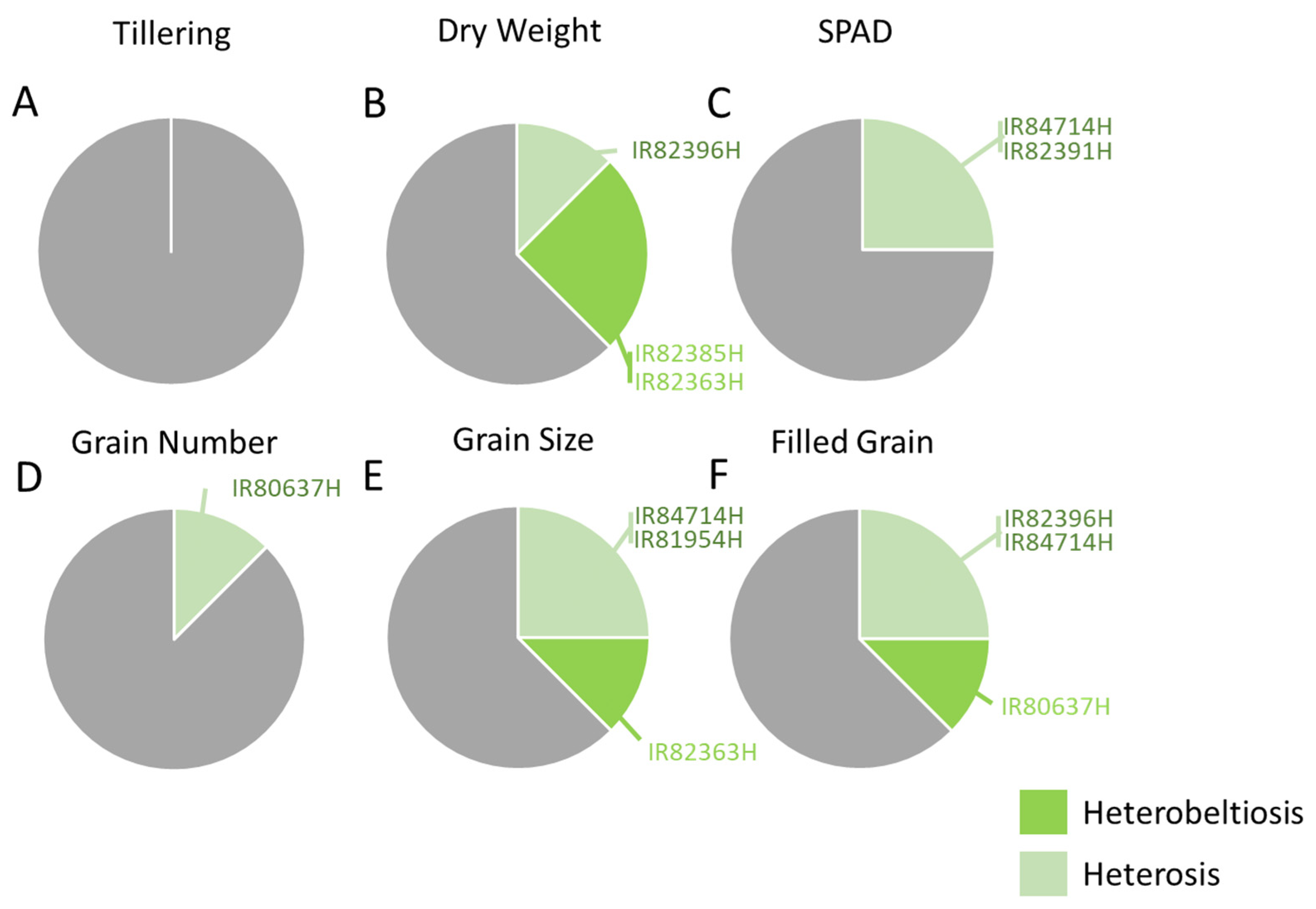
3.2. Plant Development and Herbivore Densities in Field Plots
3.2.1. Plant Biomass and Yields
3.2.2. Abundance of Planthoppers and Leafhoppers in Field Plots
3.2.3. Lepidopteran Herbivores in Field Plots
4. Discussion
4.1. Resistance to Planthoppers and Leafhoppers
4.2. Resistance to Leaf Chewers and Stemborers
4.3. Recommendations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bandumula, N. Rice production in Asia: Key to global food security. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2018, 88, 1323–1328. [Google Scholar] [CrossRef]
- Seck, P.A.; Diagne, A.; Mohanty, S.; Wopereis, M.C. Crops that feed the world 7: Rice. Food Secur. 2012, 4, 7–24. [Google Scholar] [CrossRef]
- Yuan, L.-P. Development of hybrid rice to ensure food security. Rice Sci. 2014, 21, 1. [Google Scholar] [CrossRef]
- Spielman, D.J.; Kolady, D.E.; Ward, P.S. The prospects for hybrid rice in India. Food Secur. 2013, 5, 651–665. [Google Scholar] [CrossRef]
- Virmani, S. Prospects of hybrid rice in the tropics and subtropics. In Hybrid Rice Technology: New Developments and Future Prospects; Virmani, S.S., Ed.; International Rice Research Institute: Los Baños, Philippines, 1994; pp. 7–19. [Google Scholar]
- Huang, M. The decreasing area of hybrid rice production in China: Causes and potential effects on Chinese rice self-sufficiency. Food Secur. 2022, 14, 267–272. [Google Scholar] [CrossRef]
- Huang, M. Hybrid breeding and cultivar diversity in rice production in China. Agric. Environ. Lett. 2022, 7, e20074. [Google Scholar] [CrossRef]
- Sogawa, K.; Liu, G.J.; Qiang, Q.; Heong, K.L.; Hardy, B. Prevalence of whitebacked planthoppers in Chinese hybrid rice and whitebacked planthopper resistance in Chinese japonica rice. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; Heong, K.L., Hardy, B., Eds.; International Rice Research Institute: Los Baños, Philippines, 2009; pp. 257–280. [Google Scholar]
- Sogawa, K.; Liu, G.-J.; Zhou, J.; Han, X.; You, C.-B. Causal analysis on the whitebacked planthopper prevalence in Chinese hybrid rice Shanyou 63. Chin. J. Rice Sci. 2003, 17, 95. [Google Scholar]
- Chen, J.-M.; Yu, X.-P.; Cheng, J.-A.; Lu, Z.-X.; Zheng, X.-S.; Xu, H.-X. Resistance screening and evaluation of newly-bred rice varieties (lines) to the rice brown planthopper, Nilaparvata lugens. Chin. J. Rice Sci. 2005, 19, 573. [Google Scholar]
- Zhu, Z.; Cheng, J.; Zuo, W.; Lin, X.; Guo, Y.; Jiang, Y.; Wu, X.; Teng, K.; Zhai, B.; Luo, J. Integrated management of rice stem borers in the Yangtze Delta, China. In Proceedings of the Area-Wide Control of Insect Pests: From Research to Field Implementation, Mexico City, Mexico, 29–30 November 2007; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, Germany, 2007; pp. 373–382. [Google Scholar]
- Tian, J.-R.; Li, X.-K. Managing diseases and insect pests of hybrid rice in China. In Hybrid Rice Technology: New Developments and Future Prospects; Virmani, S.S., Ed.; International Rice Research Institute: Los Baños, Philippines, 1994; pp. 115–122. [Google Scholar]
- Mew, T.W.; Wang, F.M.; Wu, J.T.; Lin, K.R.; Khush, G.S. Disease and insect resistance in hybrid rice. In Proceedings of the International Symposium on Hybrid Rice, Changsha, China, 6–10 October 1986; International Rice Research Institute: Manila, Philippines, 1988; pp. 189–200. [Google Scholar]
- Horgan, F.G.; Crisol, E. Hybrid rice and insect herbivores in Asia. Entomol. Exp. Appl. 2013, 148, 1–19. [Google Scholar] [CrossRef]
- Sogawa, K.; Liu, G.-J.; Shen, J.-H. A review on the hyper-susceptibility of Chinese hybrid rice to insect pests. Chin. J. Rice Sci. 2003, 17, 23. [Google Scholar]
- Virmani, S.; Aquino, R.; Khush, G. Heterosis breeding in rice (Oryza sativa L.). Theor. Appl. Genet. 1982, 63, 373–380. [Google Scholar] [CrossRef]
- Zhou, G.; Chen, Y.; Yao, W.; Zhang, C.; Xie, W.; Hua, J.; Xing, Y.; Xiao, J.; Zhang, Q. Genetic composition of yield heterosis in an elite rice hybrid. Proc. Natl. Acad. Sci. USA 2012, 109, 15847–15852. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Yang, S.; Gong, J.; Zhao, Y.; Feng, Q.; Gong, H.; Li, W.; Zhan, Q.; Cheng, B.; Xia, J. Genomic analysis of hybrid rice varieties reveals numerous superior alleles that contribute to heterosis. Nat. Commun. 2015, 6, 6258. [Google Scholar] [CrossRef]
- Hajjar, R.; Jarvis, D.I.; Gemmill-Herren, B. The utility of crop genetic diversity in maintaining ecosystem services. Agric. Ecosyst. Environ. 2008, 123, 261–270. [Google Scholar] [CrossRef]
- Jarvis, D.I.; Brown, A.H.D.; Imbruce, V.; Ochoa, J.; Sadiki, M.; Karamura, E.; Trutmann, P.; Finckh, M.R. Managing crop disease in traditional agroecosystems: Benefits and hazards of genetic diversity. In Managing Biodiversity in Agricultural Ecosystems; Jarvis, D.I., Ed.; Columbia University Press: New York, NY, USA, 2007; pp. 292–319. [Google Scholar]
- Chen, Q.; Zeng, G.; Hao, M.; Jiang, H.; Xiao, Y. Improvement of rice blast and brown planthopper resistance of PTGMS line C815S in two-line hybrid rice through marker-assisted selection. Mol. Breed. 2020, 40, 21. [Google Scholar] [CrossRef]
- Fan, F.; Li, N.; Chen, Y.; Liu, X.; Sun, H.; Wang, J.; He, G.; Zhu, Y.; Li, S. Development of elite BPH-resistant wide-spectrum restorer lines for three and two line hybrid rice. Front. Plant Sci. 2017, 8, 986. [Google Scholar] [CrossRef]
- Hu, J.; Cheng, M.; Gao, G.; Zhang, Q.; Xiao, J.; He, Y. Pyramiding and evaluation of three dominant brown planthopper resistance genes in the elite indica rice 9311 and its hybrids. Pest Manag. Sci. 2013, 69, 802–808. [Google Scholar] [CrossRef]
- Jiang, H.; Hu, J.; Li, Z.; Liu, J.; Gao, G.; Zhang, Q.; Xiao, J.; He, Y. Evaluation and breeding application of six brown planthopper resistance genes in rice maintainer line Jin 23B. Rice 2018, 11, 22. [Google Scholar] [CrossRef]
- Liu, M.; Fan, F.; He, S.; Guo, Y.; Chen, G.; Li, N.; Li, N.; Yuan, H.; Si, F.; Yang, F.; et al. Creation of elite rice with high-yield, superior-quality and high resistance to brown planthopper based on molecular design. Rice 2022, 15, 17. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, W.; Liu, H.; Zeng, Y.; Du, B.; Zhu, L.; He, G.; Chen, R. Marker assisted pyramiding of Bph6 and Bph9 into elite restorer line 93–11 and development of functional marker for Bph9. Rice 2017, 10, 51. [Google Scholar] [CrossRef]
- Wan, B.; Zha, Z.; Li, J.; Xia, M.; Du, X.; Lin, Y.; Yin, D. Development of elite rice restorer lines in the genetic background of R022 possessing tolerance to brown planthopper, stem borer, leaf folder and herbicide through marker-assisted breeding. Euphytica 2014, 195, 129–142. [Google Scholar] [CrossRef]
- Sun, Y.; Zhao, H.; Chen, Z.; Chen, H.; Li, B.; Wang, C.; Lin, X.; Cai, Y.; Zhou, D.; Ouyang, L.; et al. Comparison of the phenotypic performance, molecular diversity, and proteomics in transgenic rice. Plants 2023, 12, 156. [Google Scholar] [CrossRef]
- Ramesh, S.; Nagadhara, D.; Pasalu, I.C.; Kumari, A.P.; Sarma, N.P.; Reddy, V.D.; Rao, K.V. Development of stem borer resistant transgenic parental lines involved in the production of hybrid rice. J. Biotechnol. 2004, 111, 131–141. [Google Scholar] [CrossRef]
- Tu, J.; Zhang, G.; Datta, K.; Xu, C.; He, Y.; Zhang, Q.; Khush, G.S.; Datta, S.K. Field performance of transgenic elite commercial hybrid rice expressing Bacillus thuringiensis δ-endotoxin. Nat. Biotechnol. 2000, 18, 1101–1104. [Google Scholar] [CrossRef]
- Horgan, F.G.; Srinivasan, T.S.; Bentur, J.S.; Kumar, R.; Bhanu, K.V.; Sarao, P.S.; Chien, H.V.; Almazan, M.L.P.; Bernal, C.C.; Ramal, A.F.; et al. Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers: Implications for rice breeding and gene deployment. Agronomy 2017, 7, 62. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, C.; Chen, G.; Li, Y.; Yang, X.; Han, L.; Peng, Y. Downregulation of the CsABCC2 gene is associated with Cry1C resistance in the striped stem borer Chilo suppressalis. Pestic. Biochem. Physiol. 2022, 184, 105119. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Brévault, T.; Carrière, Y. Insect resistance to Bt crops: Lessons from the first billion acres. Nat. Biotechnol. 2013, 31, 510–521. [Google Scholar] [CrossRef]
- Kobayashi, T. Evolving ideas about genetics underlying insect virulence to plant resistance in rice-brown planthopper interactions. J. Insect Physiol. 2016, 84, 32–39. [Google Scholar] [CrossRef]
- Faiz, F.A.; Ijaz, M.; Awan, T.H.; Manzoor, Z.; Ahmed, M.; Wariach, N.M.; Zahid, M.A. Effect of wild abortive cytoplasm inducing male sterility on resistance/tolerance against brown plant hopper and white backed plant hopper in Basmati rice hybrids. J. Anim. Plant Sci. 2007, 17, 16–20. [Google Scholar]
- Horgan, F.G.; Crisol-Martínez, E.; Almazan, M.L.P.; Romena, A.; Ramal, A.F.; Ferrater, J.B.; Bernal, C.C. Susceptibility and tolerance in hybrid and pure-line rice varieties to herbivore attack: Biomass partitioning and resource-based compensation in response to damage. Ann. Appl. Biol. 2016, 169, 200–213. [Google Scholar] [CrossRef]
- Cohen, M.B.; Bernal, C.C.; Virmani, S.S. Do rice hybrids have heterosis for insect resistance? A study with Nilaparvata lugens (Hemiptera: Delphacidae) and Marasmia patnalis (Lepidoptera: Pyralidae). J. Econ. Entomol. 2003, 96, 1935–1941. [Google Scholar] [CrossRef]
- Crisol, E.; Almazan, M.L.P.; Jones, P.W.; Horgan, F.G. Planthopper-rice interactions: Unequal stresses on pure-line and hybrid rice under similar experimental conditions. Entomol. Exp. Appl. 2013, 147, 18–32. [Google Scholar] [CrossRef]
- Bottrell, D.G.; Schoenly, K.G. Resurrecting the ghost of green revolutions past: The brown planthopper as a recurring threat to high-yielding rice production in tropical Asia. J. Asia-Pac. Entomol. 2012, 15, 122–140. [Google Scholar] [CrossRef]
- Horgan, F.G. The structure of rice stemborer assemblages: A review of species’ distributions, host ranges, and interspecific interactions. Insects 2023, 14, 921. [Google Scholar] [CrossRef]
- Rumanti, I.; Purwoko, B.; Dewi, I.; Aswidinnoor, H.; Widyastuti, Y. Combining ability for yield and agronomic traits in hybrid rice derived from wild abortive, Gambiaca and Kalinga cytoplasmic male sterile lines. SABRAO J. Breed. Genet. 2017, 49, 69. [Google Scholar]
- Richharia, R.H.; Misro, B.; Rao, R.K. Sterility in the rice hybrids and its significance. Euphytica 1962, 11, 137–142. [Google Scholar] [CrossRef]
- Wan, J.; Yamaguchi, Y.; Kato, H.; Ikehashi, H. Two new loci for hybrid sterility in cultivated rice (Oryza sativa L.). Theor. Appl. Genet. 1996, 92, 183–190. [Google Scholar] [CrossRef]
- Yang, D.; Xiong, L.; Mou, T.; Mi, J. Improving the resistance of the rice PTGMS line Feng39S by pyramiding blast, bacterial blight, and brown planthopper resistance genes. Crop J. 2022, 10, 1187–1197. [Google Scholar] [CrossRef]
- Li, S.; Yang, D.; Zhu, Y. Characterization and use of male sterility in hybrid rice breeding. J. Integr. Plant Biol. 2007, 49, 791–804. [Google Scholar] [CrossRef]
- Xinggui, L.; Virmani, S.; Yang, R. Advances in two-lines hybrids rice breeding. In Proceedings of the Advances in Hybrid Rice Breeding: Proceedings of the 3rd International Symposium on Hybrid Rice, Hyderabad, India, 14–16 November 1996; Virmani, S.S., Siddiq, E.A., Muralidharan, K., Eds.; International Rice Research Institute: Los Baños, Philippines, 1998; pp. 89–98. [Google Scholar]
- Alam, S.N.; Cohen, M.B. Durability of brown planthopper, Nilaparvata lugens, resistance in rice variety IR64 in greenhouse selection studies. Entomol. Exp. Appl. 1998, 89, 71–78. [Google Scholar] [CrossRef]
- Khetnon, P.; Busarakam, K.; Sukhaket, W.; Niwaspragrit, C.; Kamolsukyeunyong, W.; Kamata, N.; Sanguansub, S. Mechanisms of trichomes and terpene compounds in indigenous and commercial Thai rice varieties against brown planthopper. Insects 2022, 13, 427. [Google Scholar] [CrossRef]
- Li, C.-P.; Wu, D.-H.; Huang, S.-H.; Meng, M.; Shih, H.-T.; Lai, M.-H.; Chen, L.-J.; Jena, K.K.; Hechanova, S.L.; Ke, T.-J. The Bph45 gene confers resistance against brown planthopper in rice by reducing the production of limonene. Int. J. Mol. Sci. 2023, 24, 1798. [Google Scholar] [CrossRef]
- Kamolsukyeunyong, W.; Sukhaket, W.; Pitija, K.; Thorngkham, P.; Mahatheeranont, S.; Toojinda, T.; Vanavichit, A. Rice sesquiterpene plays important roles in antixenosis against brown planthopper in rice. Plants 2021, 10, 1049. [Google Scholar] [CrossRef]
- Lin, Y.; Lin, X.; Ding, C.; Xia, M.; Xue, R.; Sun, Z.; Chen, D.; Zhu-Salzman, K.; Zeng, R.; Song, Y. Priming of rice defense against a sap-sucking insect pest brown planthopper by silicon. J. Pest Sci. 2022, 95, 1371–1385. [Google Scholar] [CrossRef]
- He, W.; Yang, M.; Li, Z.; Qiu, J.; Liu, F.; Qu, X.; Qiu, Y.; Li, R. High levels of silicon provided as a nutrient in hydroponic culture enhances rice plant resistance to brown planthopper. Crop Prot. 2015, 67, 20–25. [Google Scholar] [CrossRef]
- Wu, X.; Yu, Y.; Baerson, S.R.; Song, Y.; Liang, G.; Ding, C.; Niu, J.; Pan, Z.; Zeng, R. Interactions between nitrogen and silicon in rice and their effects on resistance toward the brown planthopper Nilaparvata lugens. Front. Plant Sci. 2017, 8, 28. [Google Scholar] [CrossRef]
- Yang, L.; Han, Y.; Li, P.; Wen, L.; Hou, M. Silicon amendment to rice plants impairs sucking behaviors and population growth in the phloem feeder Nilaparvata lugens (Hemiptera: Delphacidae). Sci. Rep. 2017, 7, 1101. [Google Scholar] [CrossRef]
- Shi, S.; Wang, H.; Nie, L.; Tan, D.; Zhou, C.; Zhang, Q.; Li, Y.; Du, B.; Guo, J.; Huang, J. Bph30 confers resistance to brown planthopper by fortifying sclerenchyma in rice leaf sheaths. Mol. Plant 2021, 14, 1714–1732. [Google Scholar] [CrossRef]
- Woodhead, S.; Padgham, D. The effect of plant surface characteristics on resistance of rice to the brown planthopper, Nilaparvata lugens. Entomol. Exp. Appl. 1988, 47, 15–22. [Google Scholar] [CrossRef]
- Lou, Y.-G.; Ma, B.; Cheng, J.-A. Attraction of the parasitoid Anagrus nilaparvatae to rice volatiles induced by the rice brown planthopper Nilaparvata lugens. J. Chem. Ecol. 2005, 31, 2357–2372. [Google Scholar] [CrossRef]
- Kartohardjono, A.; Heinrichs, E.A. Populations of the brown planthopper, Nilaparvata lugens (Stål) (Homoptera: Delphacidae), and its predators on rice varieties with different levels of resistance. Environ. Entomol. 1984, 13, 359–365. [Google Scholar] [CrossRef]
- Salim, M.; Heinrichs, E.A. Impact of varietal resistance in rice and predation on the mortality of Sogatella furcifera (Horváth) (Homoptera: Delphacidae). Crop Prot. 1986, 5, 395–399. [Google Scholar] [CrossRef]
- Horgan, F.G.; Romena, A.M.; Bernal, C.C.; Almazan, M.L.P.; Ramal, A.F. Stem borers revisited: Host resistance, tolerance, and vulnerability determine levels of field damage from a complex of Asian rice stemborers. Crop Prot. 2021, 142, 105513. [Google Scholar] [CrossRef]
- Li, J.; Yang, J.; Fei, P.; Song, J.; Li, D.; Ge, C.; Chen, W. Responses of rice leaf thickness, SPAD readings and chlorophyll a/b ratios to different nitrogen supply rates in paddy field. Field Crops Res. 2009, 114, 426–432. [Google Scholar]
- Yang, H.; Li, J.; Yang, J.; Wang, H.; Zou, J.; He, J. Effects of nitrogen application rate and leaf age on the distribution pattern of leaf SPAD readings in the rice canopy. PLoS ONE 2014, 9, e88421. [Google Scholar] [CrossRef]
- Andama, J.B.; Mujiono, K.; Hojo, Y.; Shinya, T.; Galis, I. Nonglandular silicified trichomes are essential for rice defense against chewing herbivores. Plant Cell Environ. 2020, 43, 2019–2032. [Google Scholar] [CrossRef]
- Kaur, I.; Kariyat, R.R. Eating barbed wire: Direct and indirect defensive roles of non-glandular trichomes. Plant Cell Environ. 2020, 43, 2015–2018. [Google Scholar] [CrossRef]
- Xiang, X.; Liu, S.; Li, H.; Danso Ofori, A.; Yi, X.; Zheng, A. Defense strategies of rice in response to the attack of the herbivorous insect, Chilo suppressalis. Int. J. Mol. Sci. 2023, 24, 14361. [Google Scholar] [CrossRef]
- Liu, X.; Li, J.; Xu, L.; Wang, Q.; Lou, Y. Expressing OsMPK4 impairs plant growth but enhances the resistance of rice to the striped stem borer Chilo suppressalis. Int. J. Mol. Sci. 2018, 19, 1182. [Google Scholar] [CrossRef]
- Alam, S.; Cohen, M. Detection and analysis of QTLs for resistance to the brown planthopper, Nilaparvata lugens, in a double-haploid rice population. Theor. Appl. Genet. 1998, 97, 1370–1379. [Google Scholar] [CrossRef]
- Cohen, M.B.; Alam, S.N.; Medina, E.B.; Bernal, C.C. Brown planthopper, Nilaparvata lugens, resistance in rice cultivar IR64: Mechanism and role in successful N. lugens management in Central Luzon, Philippines. Entomol. Exp. Appl. 1997, 85, 221–229. [Google Scholar] [CrossRef]
- Villegas, J.M.; Way, M.O.; Pearson, R.A.; Stout, M.J. Integrating soil silicon amendment into management programs for insect pests of drill-seeded rice. Plants 2017, 6, 33. [Google Scholar] [CrossRef]
- Villegas, J.M.; Wilson, B.E.; Way, M.O.; Gore, J.; Stout, M.J. Tolerance to rice water weevil, Lissorhoptrus oryzophilus Kuschel (Coleoptera: Curculionidae), infestations among hybrid and inbred rice cultivars in the Southern U.S. Crop Prot. 2021, 139, 105368. [Google Scholar] [CrossRef]
- Bernaola, L.; Stout, M.J. The effect of mycorrhizal seed treatments on rice growth, yield, and tolerance to insect herbivores. J. Pest Sci. 2021, 94, 375–392. [Google Scholar] [CrossRef]
- Stout, M.; Rice, W.; Ring, D. The influence of plant age on tolerance of rice to injury by the rice water weevil, Lissorhoptrus oryzophilus (Coleoptera: Curculionidae). Bull. Entomol. Res. 2002, 92, 177–184. [Google Scholar] [CrossRef]
- Stout, M.J.; McCarter, K.; Villegas, J.M.; Wilson, B.E. Natural incidence of stem borer damage in U.S. rice varieties. Crop Prot. 2024, 177, 106565. [Google Scholar] [CrossRef]
- Rubia, E.; Heong, K.; Zalucki, M.; Gonzales, B.; Norton, G. Mechanisms of compensation of rice plants to yellow stem borer Scirpophaga incertulas (Walker) injury. Crop Prot. 1996, 15, 335–340. [Google Scholar] [CrossRef]
- Rubia-Sanchez, E.; Heong, K.; Zalucki, M.; Norton, G. White stem borer damage and grain yield in irrigated rice in West Java, Indonesia. Crop Prot. 1997, 16, 665–671. [Google Scholar] [CrossRef]



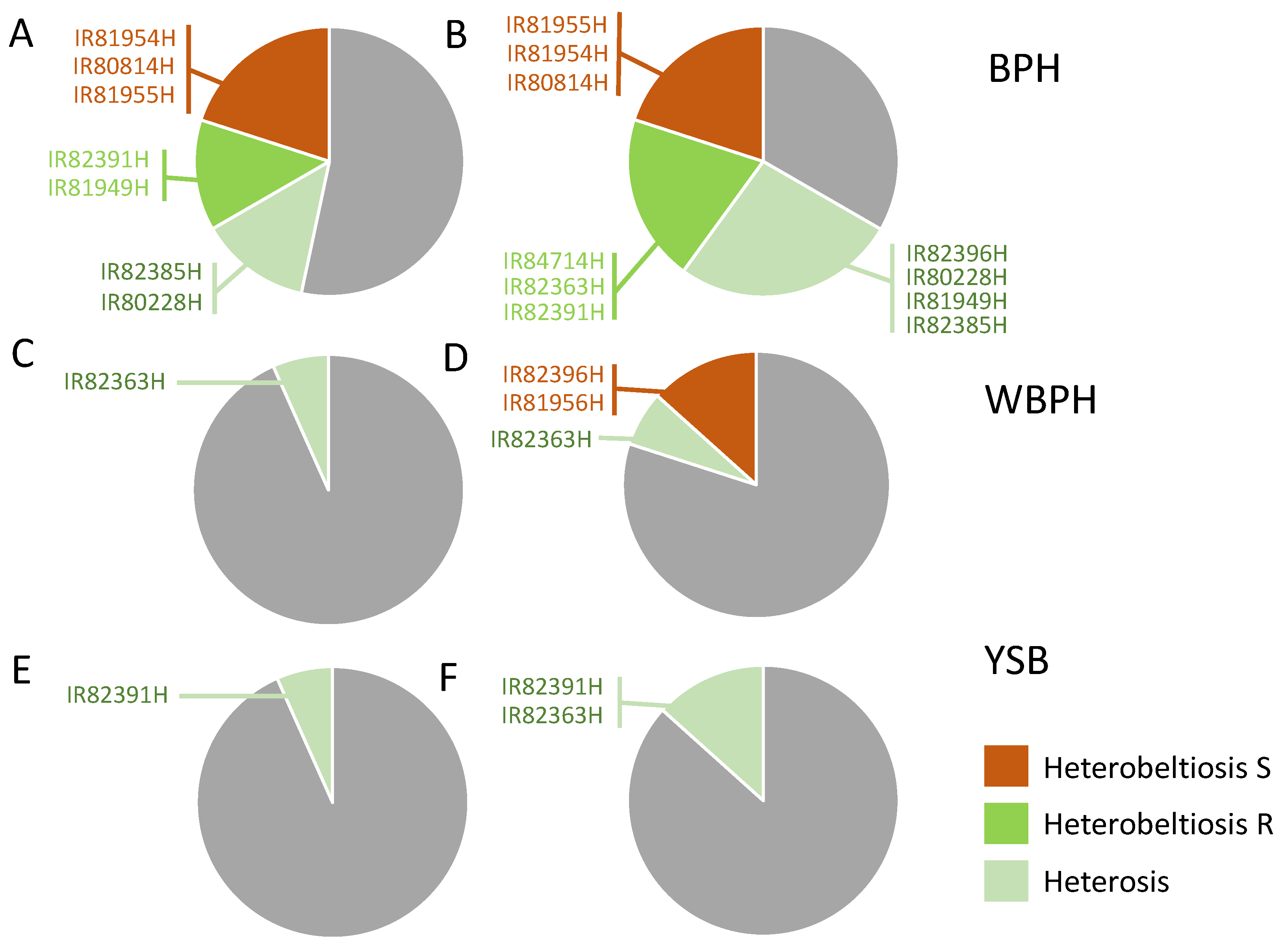{kind=link}
{kind=link}
| Accession | Harvest Time (DAT) | Yield (Tonnes Hectare−1) 1 | Production of Unfilled Grain (Tonnes Hectare−1) 1 | 1000 Filled Grain Weight (mg) 1 |
|---|---|---|---|---|
| IR82396H | 98 | 6.43 (0.12) [ht] | 0.67 (0.05) | 21.12 (0.19) a |
| IR82391H | 93 | 5.73 (0.30) [ht] | 0.85 (0.03) | 20.90 (0.23) a [ht] |
| IR84714H | 93 | 7.02 (0.10) [hb+] | 0.55 (0.36) | 22.67 (0.30) ab |
| IR85471H | 93 | 6.24 (0.18) [hb+] | 0.70 (0.10) | 22.25 (0.49) ab |
| IR81954H | 98 | 5.68 (0.20) [hb+] | 0.83 (0.57) | 25.25 (0.14) a [hb+] |
| IR80637H | 98 | 5.20 (0.33) | 0.71 (0.34) | 22.47 (0.29) ab |
| IR82385H | 112 | 6.38 (0.10) [ht] | 0.73 (0.43) [ht] | 22.88 (0.78) ab |
| IR82363H | 93 | 6.30 (0.22) | 0.70 (0.54) | 23.50 (0.25) ab [ht] |
| F-Accession | 1.414 ns | 0.497 ns | 2.520 * |
| Accession | Sampling Time (DAT) | BPH (Number Sample−1) 1 | WBPH (Number Sample−1) 1 | Virus Vectors (Number Sample−1) 1 | Other Hemiptera (Number Sample−1) 1 |
|---|---|---|---|---|---|
| IR82396H | 21 | 0.20 (0.09) | 2.80 (0.63) | 3.40 (0.91) | 0.00 (0.00) |
| 56 | 15.20 (2.38) | 16.20 (2.99) | 43.00 (8.57) | 1.00 (0.45) | |
| 84 | 0.80 (0.26) | 3.20 (0.49) | 9.60 (1.08) [ht] | 0.40 (0.18) | |
| IR82391H | 21 | 0.40 (0.18) | 3.60 (0.87) | 2.80 (0.79) | 0.00 (0.00) |
| 56 | 11.60 (2.19) | 27.40 (4.53) | 26.40 (4.57) | 1.20 (0.38) | |
| 84 | 2.40 (0.30) | 4.00 (0.87) | 13.80 (1.87) | 0.60 (0.27) | |
| IR84714H | 21 | 1.00 (0.20) | 5.60 (1.87) | 2.80 (0.60) | 0.00 (0.00) |
| 56 | 14.40 (2.55) | 12.80 (2.51) | 14.40 (2.55) | 0.80 (0.27) | |
| 84 | 0.80 (0.22) | 8.40 (2.54) | 7.00 (1.46) | 0.40 (0.18) | |
| IR85471H | 21 | 1.00 (0.24) | 4.20 (1.00) | 2.40 (0.50) | 0.20 (0.09) |
| 56 | 27.20 (7.65) | 26.20 (6.37) | 34.00 (9.92) | 0.20 (6.09) | |
| 84 | 0.80 (0.17) | 7.00 (0.81) | 8.20 (1.57) | 0.40 (0.18) | |
| IR81954H | 21 | 2.40 (0.87) | 7.80 (0.64) | 2.40 (0.59) | 0.00 (0.00) |
| 56 | 25.00 (3.97) | 43.60 (7.79) | 33.20 (6.35) | 0.20 (0.09) | |
| 84 | 1.20 (0.26) | 7.00 (2.50) | 3.80 (0.87) [ht] | 0.00 (0.00) | |
| IR80637H | 21 | 0.40 (0.18) | 5.40 (1.14) | 2.60 (0.57) | 0.00 (0.00) |
| 56 | 27.20 (3.70) | 37.40 (5.80) | 16.60 (3.97) | 1.00 (0.38) | |
| 84 | 0.60 (0.11) | 1.60 (0.61) | 2.80 (0.68) | 0.40 (0.18) | |
| IR82385H | 21 | 0.20 (0.09) | 6.20 (1.41) | 3.20 (1.02) | 0.20 (0.09) |
| 56 | 40.40 (5.70) | 29.20 (3.73) | 38.80 (8.01) | 0.60 (0.27) | |
| 84 | 1.40 (0.23) | 6.60 (1.00) | 7.80 (1.29) | 1.00 (0.31) | |
| IR82363H | 21 | 1.00 (0.24) | 6.00 (0.87) | 6.60 (1.40) | 0.00 (0.00) |
| 56 | 23.20 (3.37) | 34.80 (5.01) | 24.40 (4.47) | 0.00 (0.00) | |
| 84 | 1.40 (0.39) | 3.20 (0.89) | 9.20 (1.34) | 0.20 (0.09) | |
| F-Time | 41.726 *** | 35.708 *** | 5.993 *** | 27.674 *** | |
| F Time × Accession | 0.954 ns | 0.907 ns | 0.584 ns | 0.672 ns | |
| F-Accession | 0.887 ns | 0.810 ns | 1.338 ns | 0.748 ns | |
| Accession | Sampling Time (DAT) | C. medinalis (Number Plant−1) 1,2,4 | R. atimeta (Number Plant−1) 1,2,4 | PSB (Number Plant−1) 1,3,4 | SSB (Number Plant−1) 1,3,4 | YSB (Number Plant−1) 1,3,4 | Whitehead (Proportion Plant−1) 1,3,4 |
|---|---|---|---|---|---|---|---|
| IR82396H | 84 | 0.89 (0.08) | 0.67 (0.14) | ||||
| 98 | 0.55 (0.04) ab [ht] | 0.22 (0.03) | 0.60 (0.18) | 0.40 (0.18) | 1.60 (0.39) ab | 0.15 (0.02) ab | |
| IR82391H | 84 | 0.50 (0.07) | 0.52 (0.12) | ||||
| 93 | 0.30 (0.05) a [ht] | 0.83 (0.12) | 0.00 (0.00) | 0.00 (0.00) | 0.00 (0.00) a [ht] | 0.01 (0.00) a [ht] | |
| IR84714H | 84 | 0.52 (0.04) | 0.38 (0.06) | ||||
| 93 | 0.76 (0.10) ab | 0.84 (1.00) | 0.00 (1.00) | 0.00 (1.00) | 0.00 (0.00) a | 0.00 (0.00) a [ht] | |
| IR85471H | 84 | 0.88 (0.08) | 0.32 (0.06) | ||||
| 93 | 0.89 (0.04) b [ht] | 0.69 (0.05) | 0.20 (0.09) | 0.00 (0.00) | 0.40 (0.11) a | 0.04 (0.01) a [ht] | |
| IR81954H | 84 | 0.66 (0.11) | 0.22 (1.06) | ||||
| 98 | 0.49 (0.14) ab | 0.45 (1.06) [hb−] | 0.50 (0.14) | 0.25(0.13) | 3.50 (0.66) ab | 0.14 (0.02) ab | |
| IR80637H | 84 | 0.61 (0.11) | 0.23 (1.06) | ||||
| 98 | 0.62 (0.05) ab | 0.42 (0.08) | 0.60 (0.18) | 0.00 (0.00) 5 | 2.80 (0.52) ab | 0.26 (0.03) b | |
| IR82385H | 84 | 0.63 (0.05) | 0.55 (0.12) | ||||
| 112 | 0.39 (0.02) ab | 0.25 (0.05) | 0.60 (0.11) | 0.80 (0.17) | 5.00 (0.73) b [hb−] | 0.25 (0.02) b [hb−] | |
| IR82363H | 84 | 0.77 (0.06) | 0.47 (0.00) | ||||
| 93 | 0.55 (0.05) ab | 0.70 (0.00) [hb−] | 0.00 (0.00) | 0.40 (0.18) | 0.20 (0.09) a [ht] | 0.03 (0.01) a [ht] | |
| F-Time | 0.105 ns | 0.041 ns | |||||
| F-Time × Accession | 0.630 ns | 3.497 *** | |||||
| F-Accession | 2.896 * | 1.067 ns | 1.370 ns | 1.323 ns | 4.539 *** | 6.880 *** | |
| F-Tillers | 7.866 ** | 7.635 ** | |||||
| F-Height | 4.470 * | 9.083 *** | |||||
| F-Leafiness | 0.303 ns | 15.369 *** | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horgan, F.G.; Bernal, C.C.; Ramal, A.F.; Almazan, M.L.P.; Mundaca, E.A.; Crisol-Martínez, E. Heterosis for Resistance to Insect Herbivores in a 3-Line Hybrid Rice System. Insects 2024, 15, 164. https://doi.org/10.3390/insects15030164
Horgan FG, Bernal CC, Ramal AF, Almazan MLP, Mundaca EA, Crisol-Martínez E. Heterosis for Resistance to Insect Herbivores in a 3-Line Hybrid Rice System. Insects. 2024; 15(3):164. https://doi.org/10.3390/insects15030164
Chicago/Turabian StyleHorgan, Finbarr G., Carmencita C. Bernal, Angelee F. Ramal, Maria Liberty P. Almazan, Enrique A. Mundaca, and Eduardo Crisol-Martínez. 2024. "Heterosis for Resistance to Insect Herbivores in a 3-Line Hybrid Rice System" Insects 15, no. 3: 164. https://doi.org/10.3390/insects15030164
APA StyleHorgan, F. G., Bernal, C. C., Ramal, A. F., Almazan, M. L. P., Mundaca, E. A., & Crisol-Martínez, E. (2024). Heterosis for Resistance to Insect Herbivores in a 3-Line Hybrid Rice System. Insects, 15(3), 164. https://doi.org/10.3390/insects15030164