
Long-Term Chironomid Emergence at a Karst Tufa Barrier in Plitvice Lakes National Park, Croatia

 , , ,
, , , 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Study Design
2.2. Assessment of Factors Influencing Chironomid Phenology in Plitvice Lakes
- (1)
- Month of peak emergence (month of occurrence of the median point of emergence, i.e., when 50% of individuals had emerged);
- (2)
- Duration of emergence (number of months between the start and end of emergence); and
- (3)
- The first appearance (the month in which the species emerged for the first time).
2.3. Microhabitat Preferences
2.4. Determination of the Magnitude of Chironomid Emergence
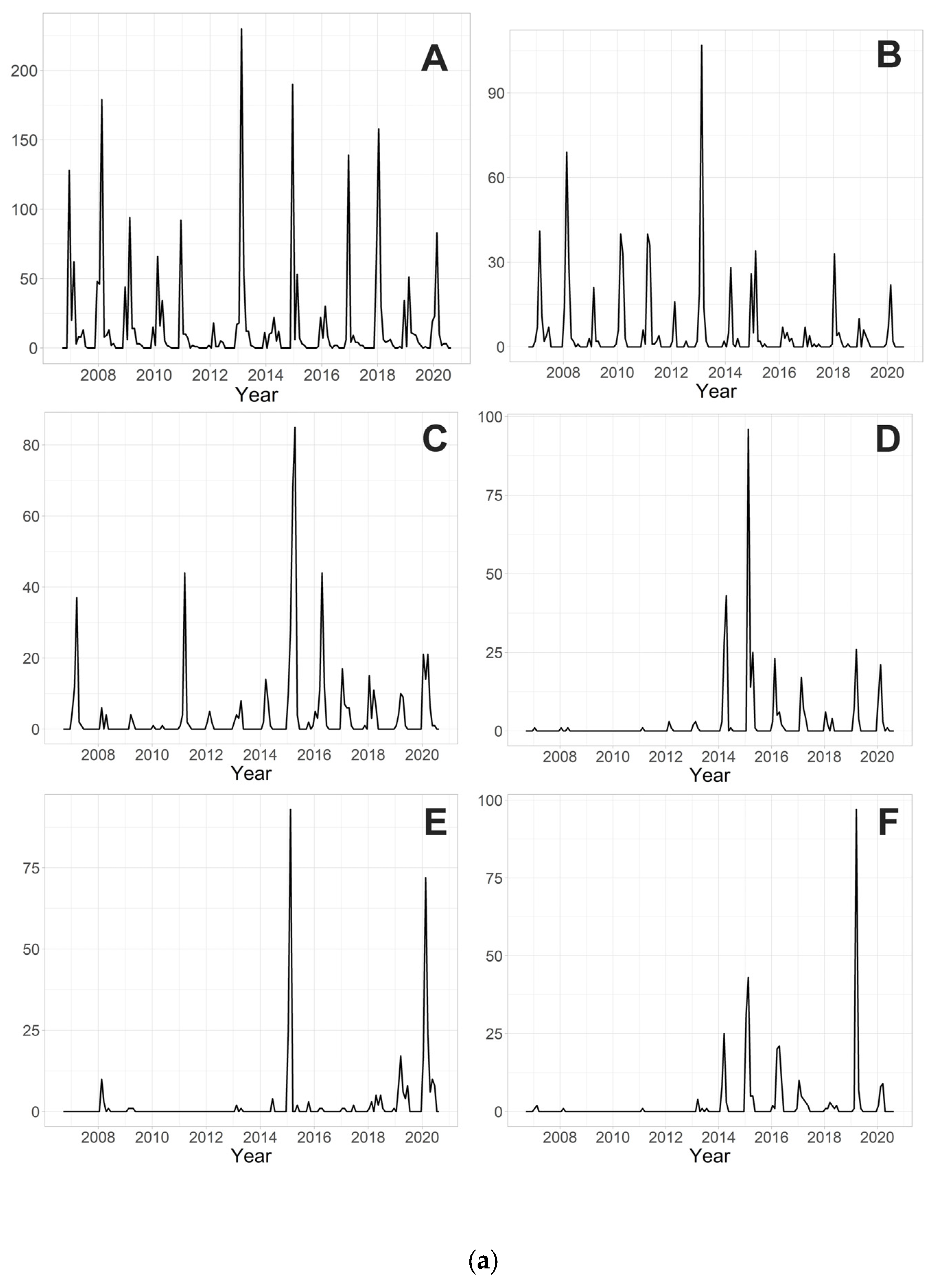
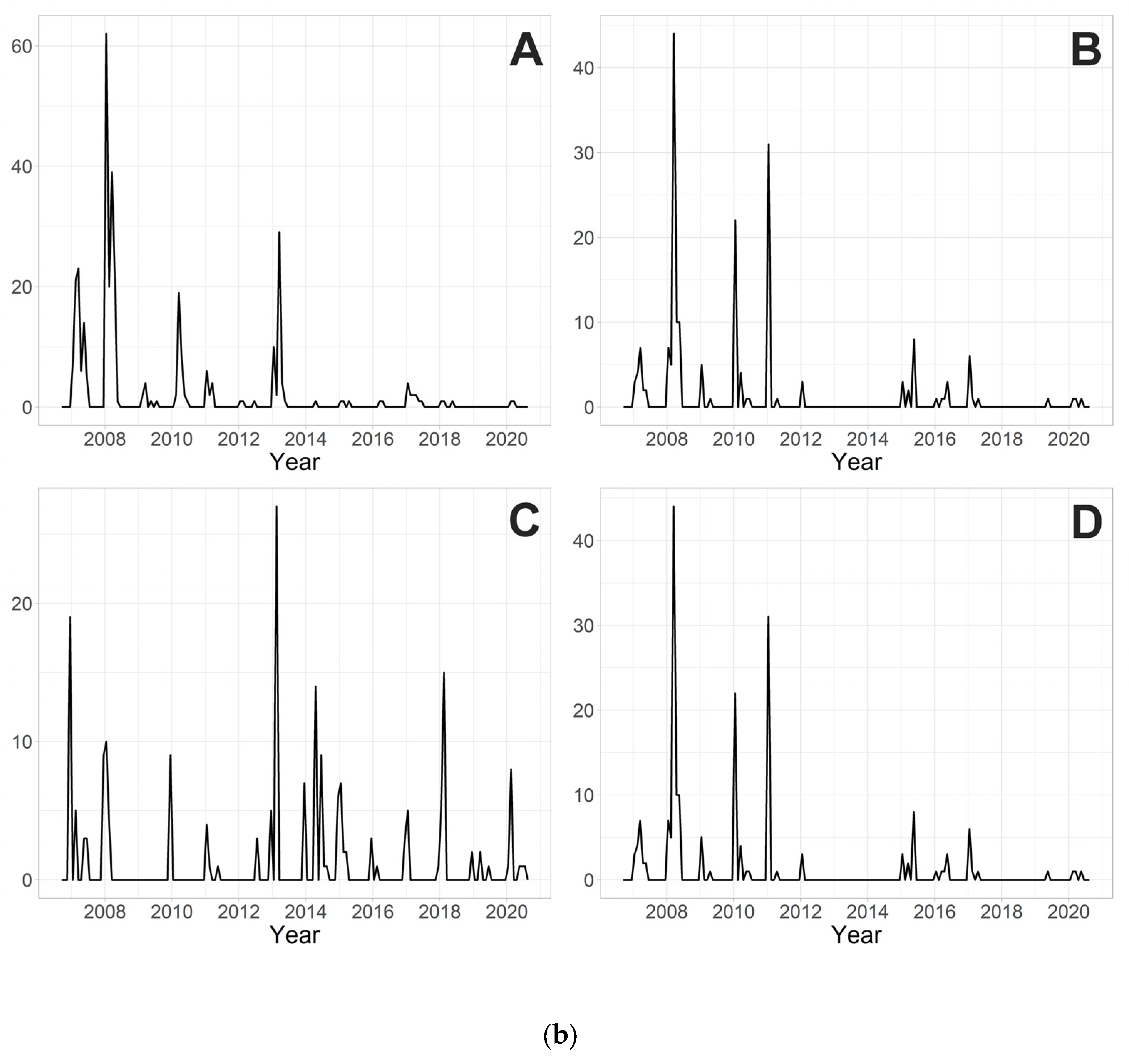
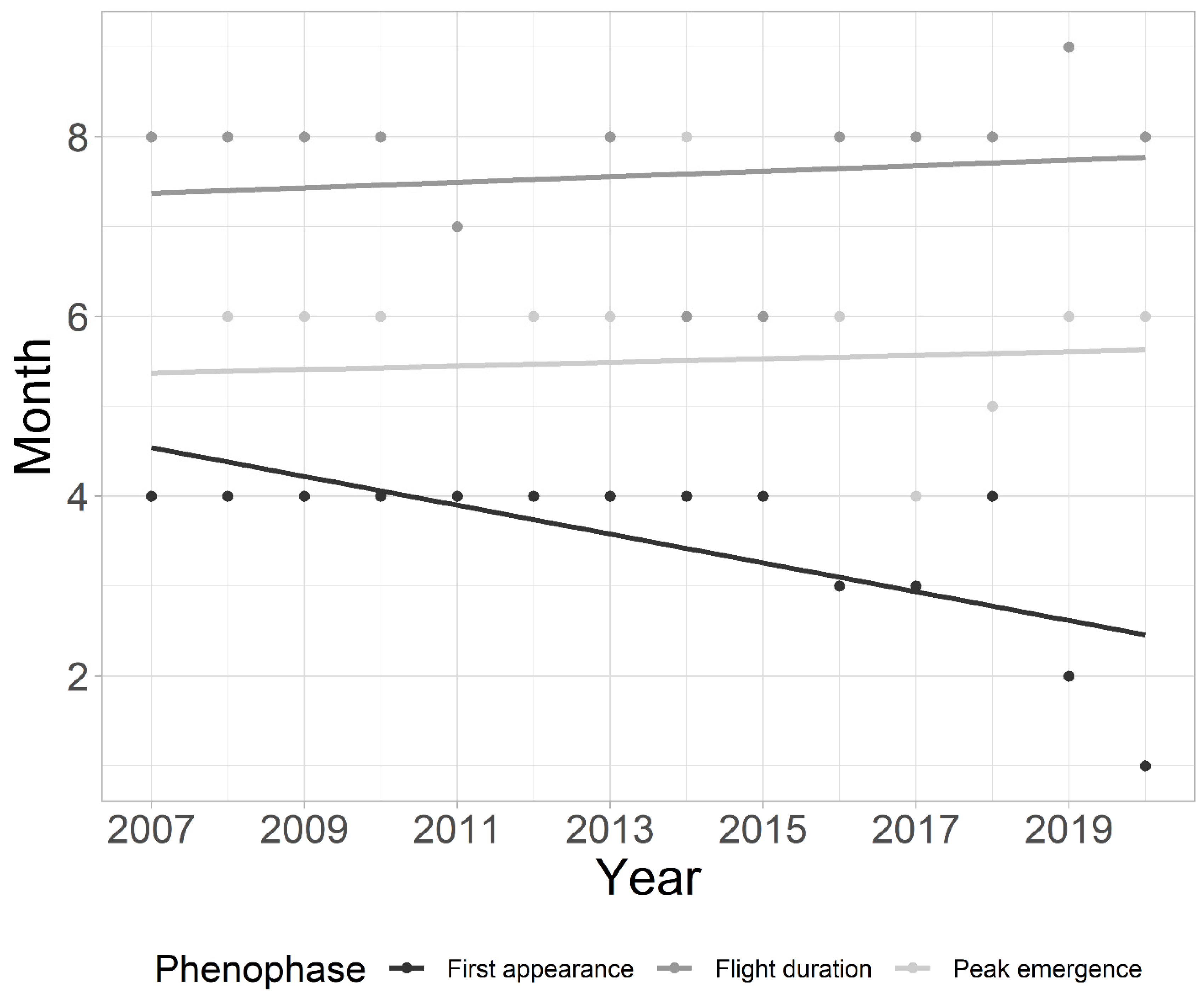
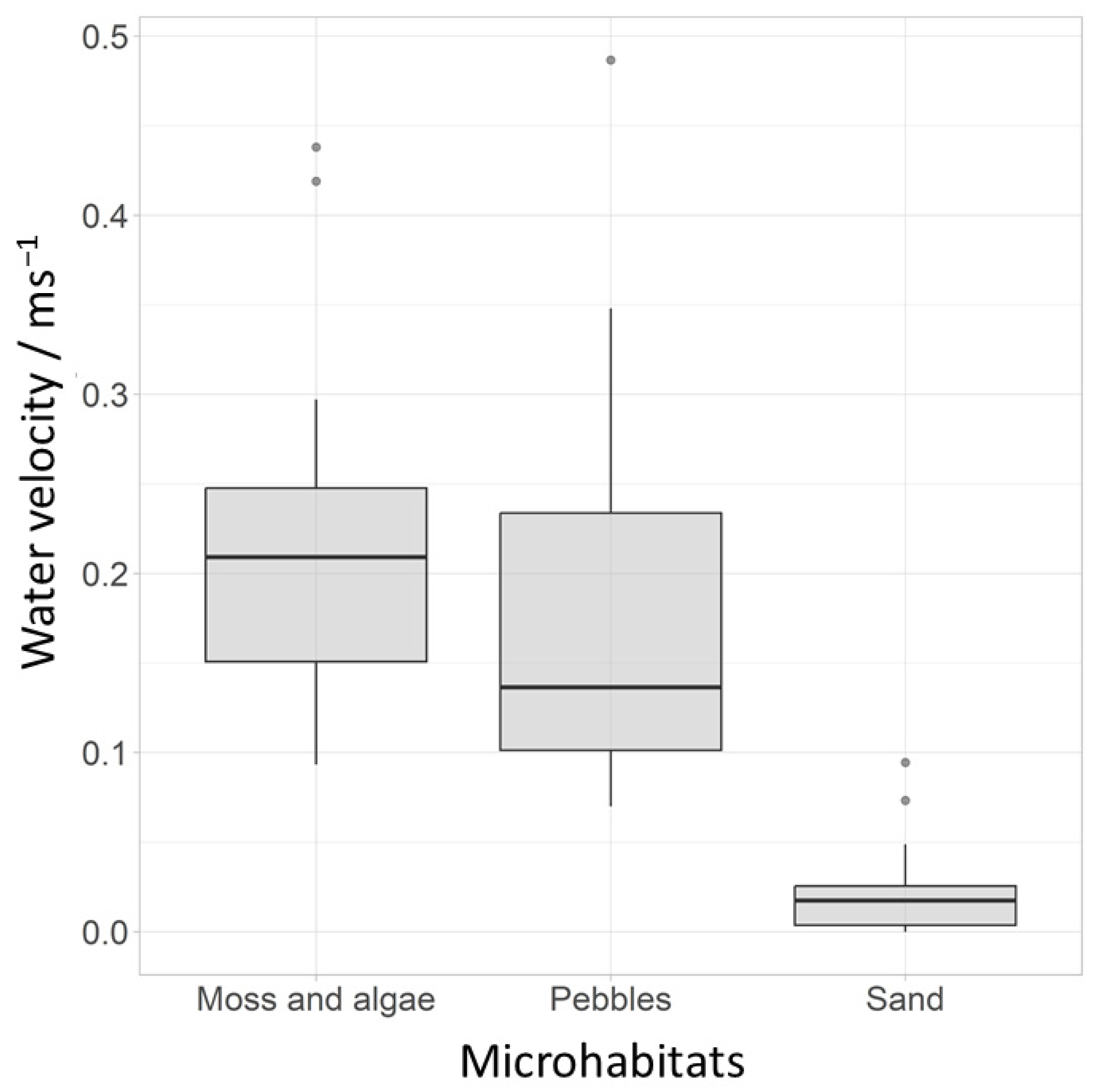
3. Results
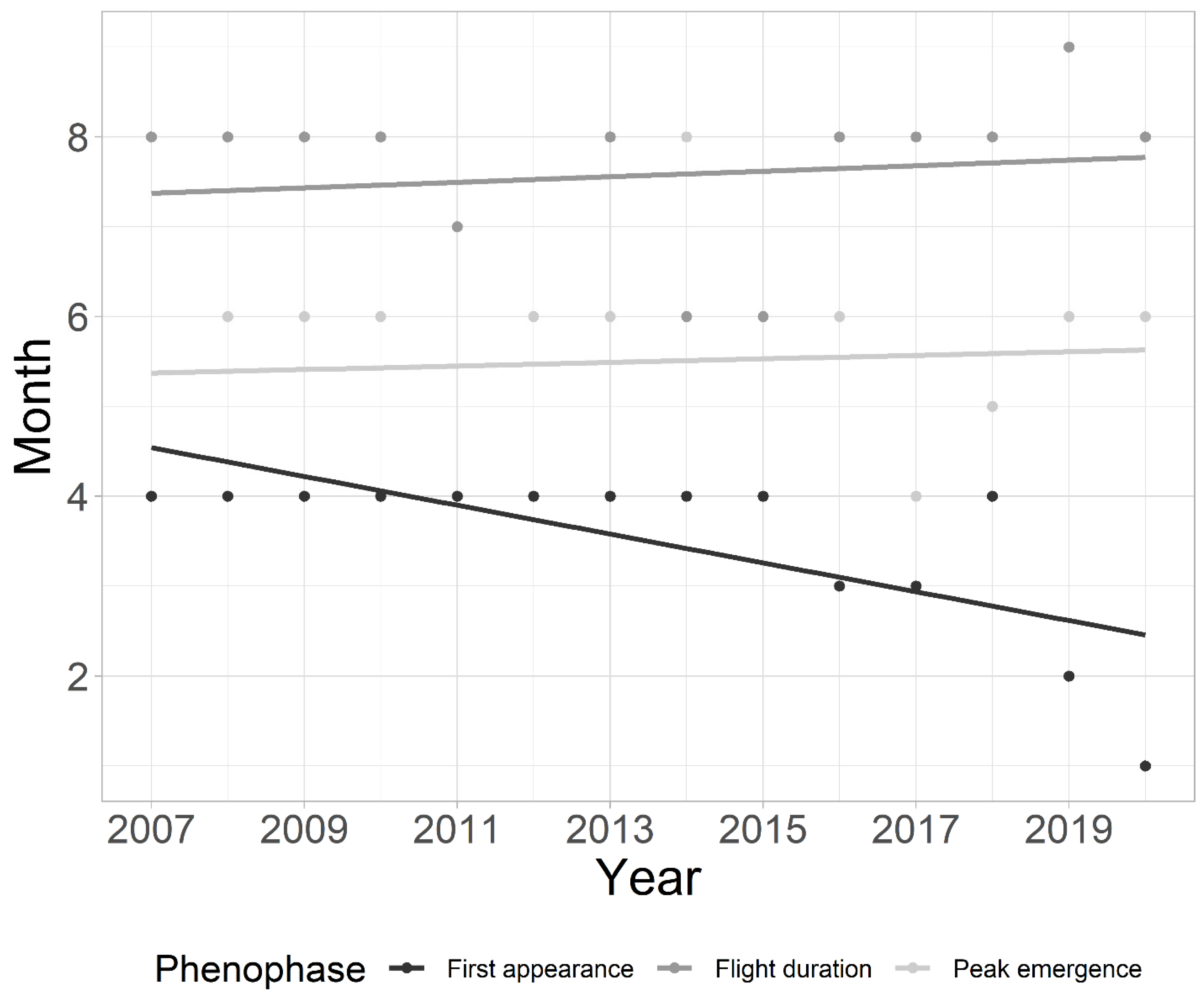
3.1. Assessment of Factors Influencing Chironomid Phenology in Plitvice Lakes
3.2. Microhabitat Preferences
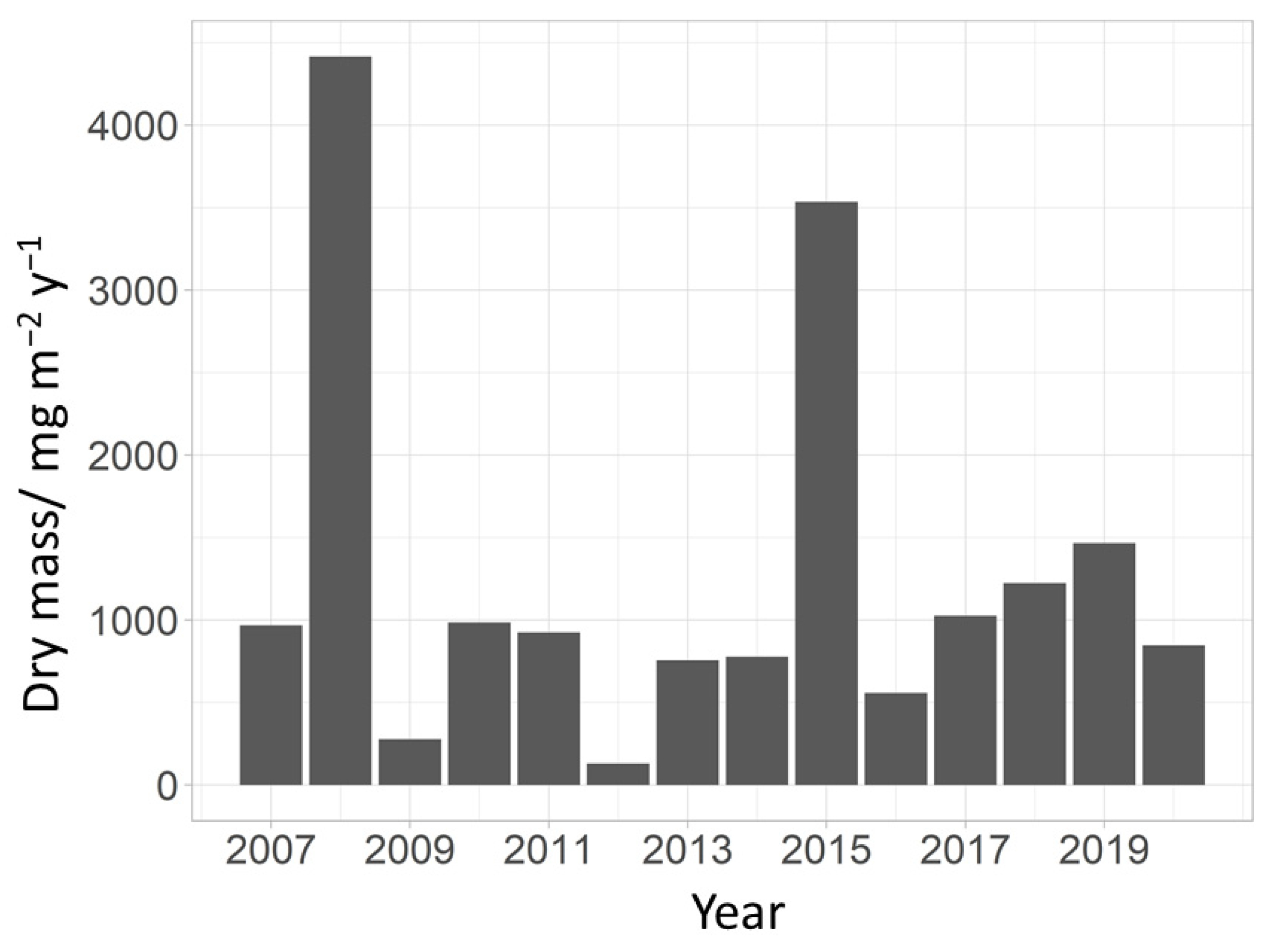
3.3. Determination of the Magnitude of Chironomid Emergence
4. Discussion
4.1. Chironomid Community at a Tufa Barrier
4.2. Factors Influencing Chironomid Phenology in Plitvice Lakes
4.3. Microhabitat Preferences
4.4. Magnitude of Chironomid Emergence
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lancaster, J.; Downes, B.J. Aquatic Entomology; Oxford University Press: Oxford, UK, 2013; 448p. [Google Scholar]
- Nakano, S.; Murakami, M. Reciprocal subsidies: Dynamic interdependence between terrestrial and aquatic food webs. Proc. Natl. Acad. Sci. USA 2001, 98, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Paetzold, A.; Tockner, K. Effects of riparian arthropod predation on the biomass and abundance of aquatic insect emergence. Freshw. Sci. 2005, 24, 395–402. [Google Scholar] [CrossRef]
- Dreyer, J.; Townsend, P.A.; Hook, J.C., III; Hoekman, D.; Vander Zanden, M.J.; Gratton, C. Quantifying aquatic insect deposition from lake to land. Ecology 2015, 96, 499–509. [Google Scholar] [CrossRef]
- Shipley, J.R.; Twining, C.W.; Mathieu-Resuge, M.; Parmar, T.P.; Kainz, M.; Martin-Creuzburg, D.; Weber, C.; Winkler, D.W.; Graham, C.H.; Matthews, B. Climate change shifts the timing of nutritional flux from aquatic insects. Curr. Biol. 2022, 32, 1342–1349. [Google Scholar] [CrossRef] [PubMed]
- Ivković, M.; Miliša, M.; Previšić, A.; Popijač, A.; Mihaljević, Z. Environmental control of emergence patterns: Case study of changes in hourly and daily emergence of aquatic insects at constant and variable water temperatures. Int. Rev. Hydrobiol. 2013, 98, 104–115. [Google Scholar] [CrossRef]
- Armitage, P.; Cranston, P.S.; Pinder, L.C.V. The Chironomidae: The Biology and Ecology of Non-Biting Midges; Chapman & Hall: London, UK, 1995; 572p. [Google Scholar]
- Coler, B.G.; Kondratieff, B.C. Emergence of Chironomidae (Diptera) from a delta-swamp receiving thermal effluent. Hydrobiologia 1989, 174, 67–77. [Google Scholar] [CrossRef]
- Baranov, V.; Lewandowski, J.; Romeijn, P.; Singer, G.; Krause, S. Effects of bioirrigation of non-biting midges (Diptera: Chironomidae) on lake sediment respiration. Sci. Rep. 2016, 6, 27329. [Google Scholar] [CrossRef] [PubMed]
- Dorić, V.; Pozojević, I.; Vučković, N.; Ivković, M.; Mihaljević, Z. Lentic chironomid performance in species-based bioassessment proving: High-level taxonomy is not a dead end in monitoring. Ecol. Indic. 2021, 121, 107041. [Google Scholar] [CrossRef]
- Walkowiak, M.; Paasivirta, L.; Meriläinen, J.J.; Arvola, L. Contrasting Patterns in Chironomid (Chironomidae) Communities of Shallow and Deep Boreal Lakes Since the 1960s. Ann. Zool. Fenn. 2016, 53, 35–51. [Google Scholar] [CrossRef]
- Lencioni, V.; Cranston, P.S.; Makarchenko, E. Recent Advances in the Study of Chironomidae: An Overview. J. Limnol. 2018, 77 (Suppl. 1), 1–6. [Google Scholar] [CrossRef]
- Oliver, D.R. Life history of the Chironomidae. Annu. Rev. Entomol. 1971, 16, 211–229. [Google Scholar] [CrossRef]
- Vallenduuk, H.J.; Moller Pillot, H.K.M. Chironomidae Larvae of the Netherlands and Adjacent Lowlands: General Ecology and Tanypodinae; KNNV Publishing: Zeist, The Netherlands, 2007; 140p. [Google Scholar]
- Pallottini, M.; Pagliarini, S.; Catasti, M.; Giontella, L.; La Porta, G.; Selvaggi, R.; Gaino, E.; Spacone, L.; Di Giulio, A.M.; Ali, A.; et al. Adult Chironomid (Chironomidae: Diptera) Positive Phototactic Behaviour—A Cue for Adult Population Management and Impact on Insect Biodiversity at Lake Trasimeno, Central Italy. Environments 2024, 11, 1. [Google Scholar] [CrossRef]
- Ivković, M.; Plant, A. Aquatic insects in the dinarides: Identifying hotspots of endemism and species richness shaped by geological and hydrological history using empididae (Diptera). Insect Conserv. Divers. 2015, 8, 302–312. [Google Scholar] [CrossRef]
- Vurnek, M.; Brozinčević, A.; Matoničkin Kepčija, R.; Frketić, T. Analyses of long-term trends in water quality data of the Plitvice Lakes National Park. Fundam. Appl. Limnol. 2021, 194, 155–169. [Google Scholar] [CrossRef]
- Šemnički, P.; Previšić, A.; Ivković, M.; Čmrlec, K.; Mihaljević, Z. Tufa Barriers from a Caddisfly’s Point of View: Streams or Lake Outlets? Int. Rev. Hydrobiol. 2012, 97, 465–484. [Google Scholar] [CrossRef]
- Vilenica, M.; Ivković, M.; Sartori, M.; Mihaljević, Z. Mayfly emergence along an oligotrophic dinaric karst hydrosystem: Spatial and temporal patterns, and species–environment relationship. Aquat. Ecol. 2017, 51, 417–433. [Google Scholar] [CrossRef]
- Vilenica, M.; Ivković, M. A decade-long study on mayfly emergence patterns. Mar. Freshw. Res. 2021, 72, 507–519. [Google Scholar] [CrossRef]
- Ridl, A.; Vilenica, M.; Ivković, M.; Popijač, A.; Sivec, I.; Miliša, M.; Mihaljević, M. Environmental drivers influencing stonefly assemblages along a longitudinal gradient in karst lotic habitats. J. Limnol. 2018, 77, 412–427. [Google Scholar] [CrossRef]
- Vilenica, M. Ecological traits of dragonfly (Odonata) assemblages along an oligotrophic Dinaric karst hydrosystem. Ann. Limnol. Int. J. Lim. 2017, 53, 377–389. [Google Scholar] [CrossRef]
- Vilenica, M.; Mičetić Stanković, V.; Mihaljević, Z.; Kučinić, M. Alderfly assemblages (Megaloptera, Sialidae) along an oligotrophic karst hydrosystem: Spatial patterns and species-environment relationship. Biologia 2018, 73, 493–503. [Google Scholar] [CrossRef]
- Ivković, M.; Weißmair, W. Faunistics and distribution of aquatic Neuroptera in Croatia. Nat. Croat. 2011, 20, 449–454. [Google Scholar]
- Baranov, V.; Ivković, M.; Willassen, E. First record of Diamesa thomasi Serra-Tosio, 1970, from Croatia. CHIRONOMUS J. Chironomidae Res. 2013, 26, 1623. [Google Scholar] [CrossRef]
- Ivković, M.; Mičetić Stanković, V.; Mihaljević, Z. Emergence patterns and microhabitat preference of aquatic dance flies (Empididae; clinocerinae and Hemerodromiinae) on a longitudinal gradient of barrage lake system. Limnologica 2012, 42, 43–49. [Google Scholar] [CrossRef]
- Ivković, M.; Kesić, M.; Mihaljević, Z.; Kúdela, M. Emergence patterns and ecological associations of some haematophagous blackfly species along an oligotrophic hydrosystem. Med. Vet. Entomol. 2014, 28, 94–102. [Google Scholar] [CrossRef]
- Ivković, M.; Miliša, M.; Baranov, V.; Mihaljević, Z. Environmental drivers of biotic traits and phenology patterns of diptera assemblages in karst springs: The role of canopy uncovered. Limnologica 2015, 54, 44–57. [Google Scholar] [CrossRef]
- Ivković, M.; Pont, A.C. Long-time emergence patterns of limnophora species (Diptera, Muscidae) in specific karst habitats: Tufa barriers. Limnologica 2016, 61, 29–35. [Google Scholar] [CrossRef]
- Ivković, M.; Dorić, V.; Baranov, V.; Mihaljević, Z.; Kolcsár, L.P.; Kvifte, G.M.; Nerudova, J.; Pont, A.C. Checklist of aquatic diptera (Insecta) of Plitvice Lakes National Park, Croatia, a UNESCO world heritage site. Zookeys 2020, 918, 99–142. [Google Scholar] [CrossRef]
- Dorić, V.; Ivković, M.; Baranov, V.; Pozojević, I.; Mihaljević, Z. Extreme freshwater discharge events exacerbated by climate change influence the structure and functional response of the chironomid community in a biodiversity hotspot. Sci. Total Environ. 2023, 879, 163110. [Google Scholar] [CrossRef] [PubMed]
- Pozojević, I.; Dorić, V.; Miliša, M.; Ternjej, I.; Ivković, M. Defining Patterns and Rates of Natural vs. Drought Driven Aquatic Community Variability Indicates the Ongoing Need for Long Term Ecological Research. Biology 2023, 12, 590. [Google Scholar] [CrossRef]
- Capezzuoli, E.; Gandin, A.; Pedley, M. Decoding tufa and travertine (freshwater carbonates) in the sedimentary record: The state of the art. Sedimentology 2013, 61, 1–21. [Google Scholar] [CrossRef]
- Radišić, M.; Rubinić, J.; Ružić, I.; Brozinčević, A. Hydrological system of the Plitvice Lakes—Trends and changes in water levels, inflows, and losses. Hydrology 2021, 8, 174. [Google Scholar] [CrossRef]
- Reiss, F.; Fittkau, E.F. Taxonomie und Ökologie europäisch verbreiteter tanytarsusarten (Chironomidae, Diptera). Arch. Hydrobiol. 1971, 40, 75–200. [Google Scholar]
- Hirvenoja, M. Revision der gattung Cricotopus van der wulp und ihrer verwandten (Diptera, Chironomidae). Ann. Zool. Fenn. 1973, 10, 1–363. [Google Scholar]
- Saether, O.A. A review of the genus Rheocricotopus Thienemann & Harnisch, 1932, with the description of three new species. Spixiana 1985, 11, 59–108. [Google Scholar]
- Wiederholm, T. Chironomidae of the Holarctic region—Keys and diagnoses: Part 3. Adult males. Entomol. Scand. Suppl. 1989, 34, 1–532. [Google Scholar]
- Spies, M. A contribution to the knowledge of holarctic parachironomus Lenz (Diptera: Chironomidae), with two new species and a provisional key to Nearctic adult males. Tijdschr. Voor Entomol. 2000, 143, 125–143. [Google Scholar] [CrossRef]
- Langton, P.H.; Pinder, L.C.V. Keys to the Adult Male Chironomidae of Britain and Ireland; Freshwater Biological Association: Newby Bridge, UK, 2007; Volume 1, 239p. [Google Scholar]
- Langton, P.H.; Pinder, L.C.V. Keys to the Adult Male Chironomidae of Britain and Ireland; Freshwater Biological Association: Newby Bridge, UK, 2007; Volume 2, 168p. [Google Scholar]
- Giłka, W. Klucze do oznaczania owadów Polski. Pol. J. Entomol. 2011, 177, 95. [Google Scholar]
- Ivković, M.; Miliša, M. (Eds.) Plitvice Lakes; Springer: Cham, Switzerland, 2023; pp. 243–274. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, Version 5.0; Microcomputer Power: Ithaca, NY, USA, 2012; 496p. [Google Scholar]
- IBM Corporation. 2020 IBM SPSS Statistics for Windows, Version 27.0; IBM Corporation: Armonk, NY, USA, 2020. [Google Scholar]
- Komsta, L.; Komsta, M.L. Package mblm. 2013. Available online: https://cran.r-project.org/web/packages/mblm/index.html (accessed on 15 October 2023).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; Available online: https://ggplot2.tidyverse.org (accessed on 15 October 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 15 October 2023).
- Moller Pillot, H.K.M. Chironomidae Larvae of The Netherlands and Adjacent Lowlands: Biology and Ecology of Aquatic Orthocladiinae; KNNV Publishing: Zeist, The Netherlands, 2013; 312p. [Google Scholar]
- Baranov, V.; Milošević, D.; Kurz, M.J.; Zarnetske, J.P.; Sabater, F.; Marti, E.; Robertson, A.; Brandt, T.; Sorolla, A.; Lewandowski, J.; et al. Helophyte impacts on the response of hyporheic invertebrate communities to inundation events in intermittent streams. Ecohydrology 2017, 10, e1857. [Google Scholar] [CrossRef]
- Andersen, T.; Cranston, P.S.; Epler, J.H. The larvae of chironomidae (Diptera) of the holarctic region–keys and diagnoses. Insect Syst. Evol. 2013, 66, 1–571. [Google Scholar]
- Jyväsjärvi, J.; Virtanen, R.; Ilmonen, J.; Paasivirta, L.; Muotka, T. Identifying taxonomic and functional surrogates for spring biodiversity conservation. Conserv. Biol. 2018, 32, 883–893. [Google Scholar] [CrossRef]
- Moller Pillot, H.K.M. Chironomidae Larvae of the Netherlands and Adjacent Lowlands: Biology and Ecology of the Chironomini; KNNV Publishing: Zeist, The Netherlands, 2009; 270p. [Google Scholar]
- Čmrlec, K.; Ivković, M.; Šemnički, P.; Mihaljević, Z. Emergence phenology and microhabitat distribution of aquatic diptera community at the outlets of barrage lakes: Effects of temperature, substrate and current velocity. Pol. J. Ecol. 2013, 61, 135–144. [Google Scholar]
- Finn, D.S.; Johnson, S.L.; Gerth, W.J.; Arismendi, I.; Li, J.L. Spatiotemporal patterns of emergence phenology reveal complex species-specific responses to temperature in aquatic insects. Divers. Distrib. 2021, 28, 1524–1541. [Google Scholar] [CrossRef]
- Ivanković, L.; Ivković, M.; Stanković, I. Perennial phenology patterns and ecological traits of Dixidae (Insecta, Diptera) in lotic habitats of a barrage lake system. Limnologica 2019, 76, 11–18. [Google Scholar] [CrossRef]
- Baranov, V.; Jourdan, J.; Pilotto, F.; Wagner, R.; Hasse, P. Complex and nonlinear climate-driven changes in freshwater insect communities over 42 years. Conserv. Biol. 2020, 34, 1241–1251. [Google Scholar] [CrossRef]
- Giller, P.S.; Malmqvist, B. The Biology of Streams and Rivers; Oxford University Press: Oxford, UK, 1998; 304p. [Google Scholar]
- Hamerlík, L.; Šporka, F.; Zaťovičová, Z. Macroinvertebrates of inlets and outlets of the Tatra Mountain lakes (Slovakia). Biologia 2006, 61, 167–179. [Google Scholar] [CrossRef]
- Miliša, M.; Habdija, I.; Primc-Habdija, B.; Radanović, I.; Matoničkin Kepčija, R. The Role of Flow Velocity in the Vertical Distribution of Particulate Organic Matter on Moss-covered Travertine Barriers of the Plitvice Lakes (Croatia). Hydrobiologia 2006, 553, 231–243. [Google Scholar] [CrossRef]
- Hanžek, N.; Gligora Udovič, M.; Kajan, K.; Borics, G.; Várbíró, G.; Stoeck, T.; Žutinić, P.; Orlić, S.; Stanković, I. Assessing ecological status in karstic lakes through the integration of phytoplankton functional groups, morphological approach and environmental DNA metabarcoding. Ecol. Indic. 2021, 131, 108166. [Google Scholar] [CrossRef]
- Gratton, C.; Donaldson, J.; Vander Zanden, M.J. Ecosystem Linkages Between Lakes and the Surrounding Terrestrial Landscape in Northeast Iceland. Ecosystems 2008, 11, 764–774. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Variable | Explains % | Contribution % | Pseudo-F | p | P (adj) |
|---|---|---|---|---|---|
| Temperature/°C | 5.1 | 24.2 | 6.5 | 0.002 | 0.024 |
| COD_KMnO4/mg O2 L−1 | 4 | 19 | 5.3 | 0.002 | 0.024 |
| Oxygen/mg O2 L−1 | 1.9 | 9.3 | 2.6 | 0.002 | 0.024 |
| Nitrites/mg N L−1 | 1.7 | 8.3 | 2.4 | 0.002 | 0.024 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dorić, V.; Pozojević, I.; Baranov, V.; Mihaljević, Z.; Ivković, M. Long-Term Chironomid Emergence at a Karst Tufa Barrier in Plitvice Lakes National Park, Croatia. Insects 2024, 15, 51. https://doi.org/10.3390/insects15010051
Dorić V, Pozojević I, Baranov V, Mihaljević Z, Ivković M. Long-Term Chironomid Emergence at a Karst Tufa Barrier in Plitvice Lakes National Park, Croatia. Insects. 2024; 15(1):51. https://doi.org/10.3390/insects15010051
Chicago/Turabian StyleDorić, Valentina, Ivana Pozojević, Viktor Baranov, Zlatko Mihaljević, and Marija Ivković. 2024. "Long-Term Chironomid Emergence at a Karst Tufa Barrier in Plitvice Lakes National Park, Croatia" Insects 15, no. 1: 51. https://doi.org/10.3390/insects15010051
APA StyleDorić, V., Pozojević, I., Baranov, V., Mihaljević, Z., & Ivković, M. (2024). Long-Term Chironomid Emergence at a Karst Tufa Barrier in Plitvice Lakes National Park, Croatia. Insects, 15(1), 51. https://doi.org/10.3390/insects15010051