
Comparing Different Sticky Traps to Monitor the Occurrence of Philaenus spumarius and Neophilaenus campestris, Vectors of Xylella fastidiosa, in Different Crops

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Evaluation of the Different Colored Sticky Traps in Olive Groves and Almond Orchards
2.2. Evaluation of the Efficiency of the Different Yellow Sticky Traps in the Olive Groves, and in the Cherry and Almond Orchards
2.3. Monitoring the Vectors on Different Vegetation Compartments
2.4. Testing the Traps in Sites Characterized by Different Spittlebug Density Populations
2.5. Data Analyses
3. Results
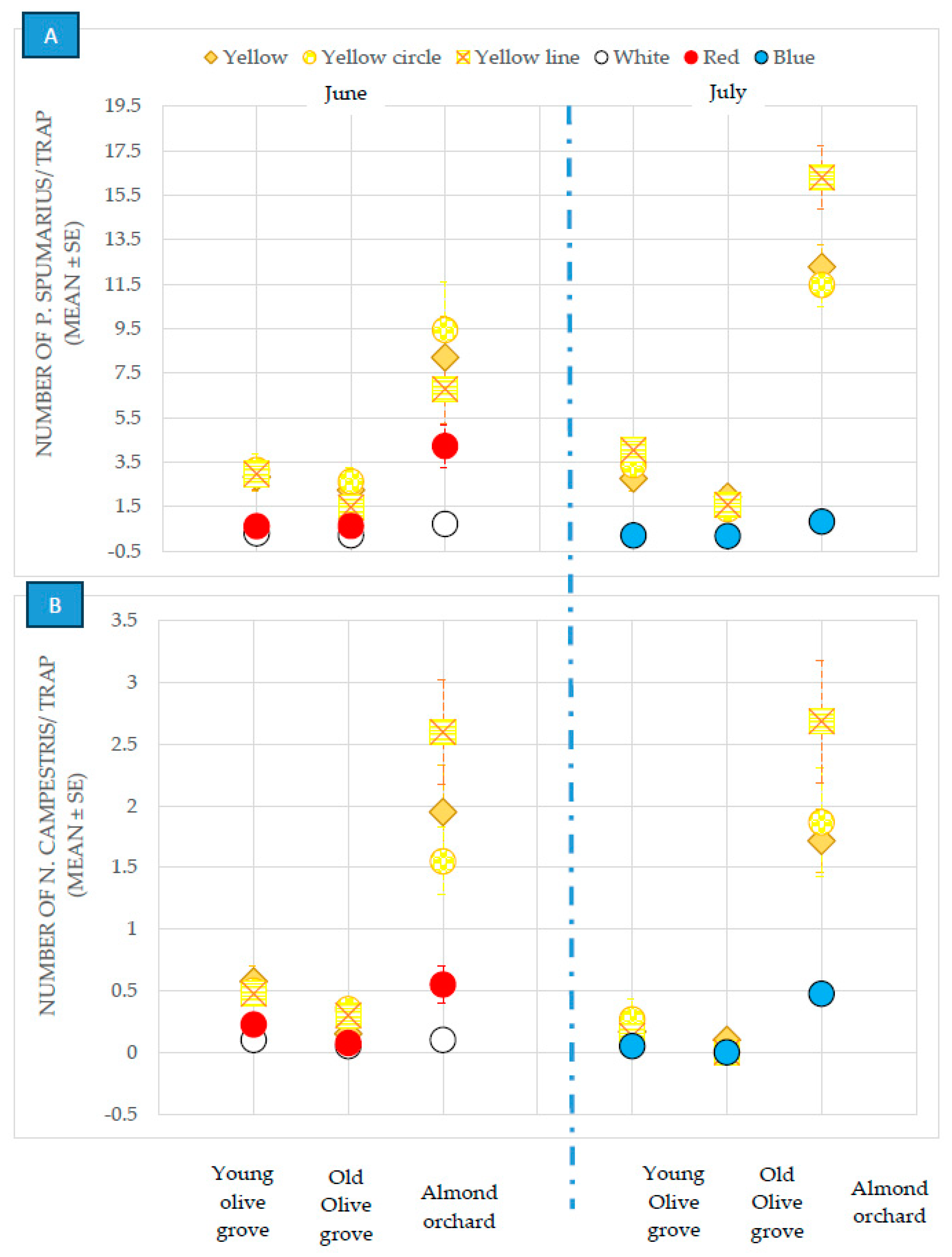
3.1. Evaluation of the Different Colored Sticky Traps in the Olive Groves and Almond Orchards
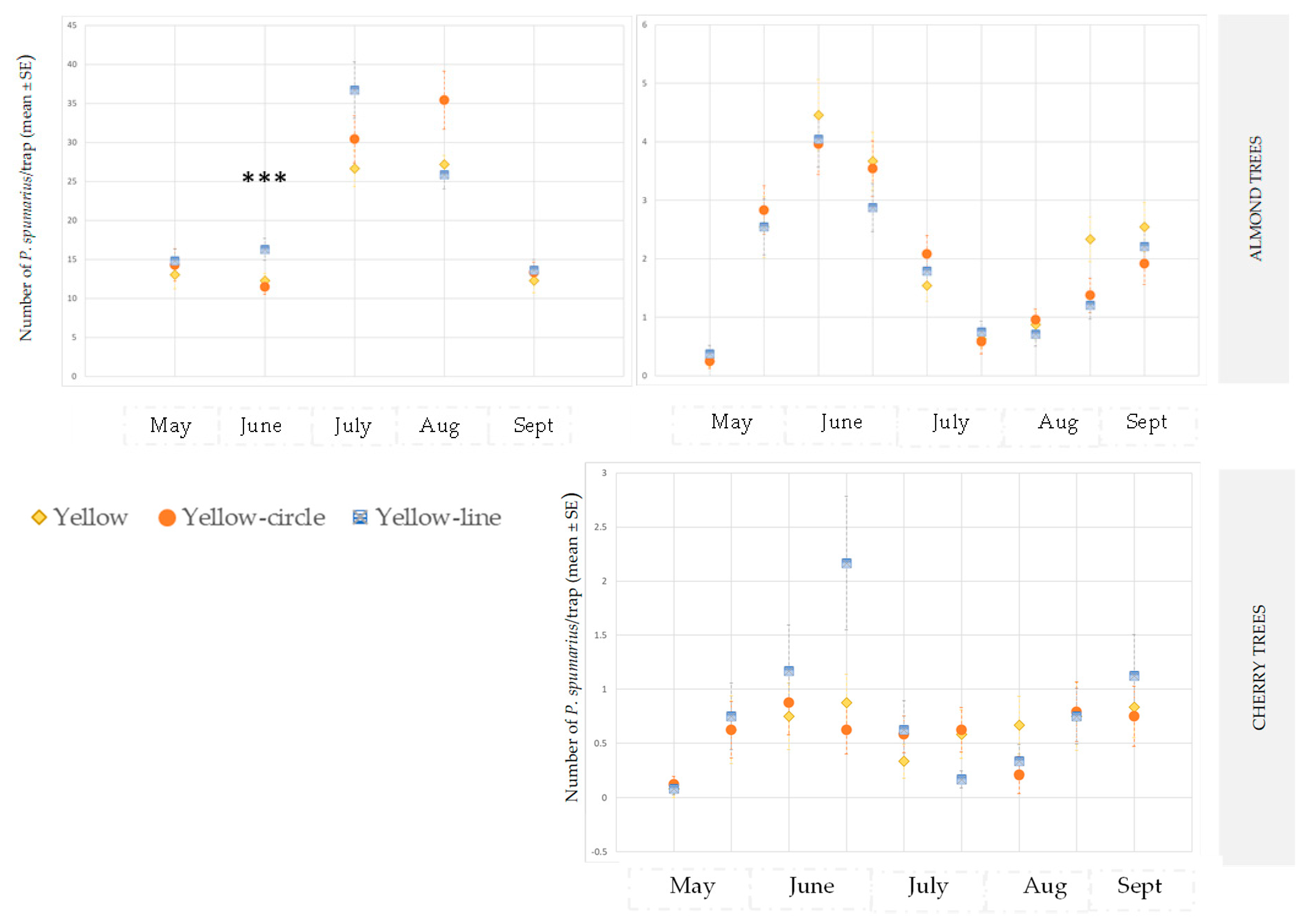
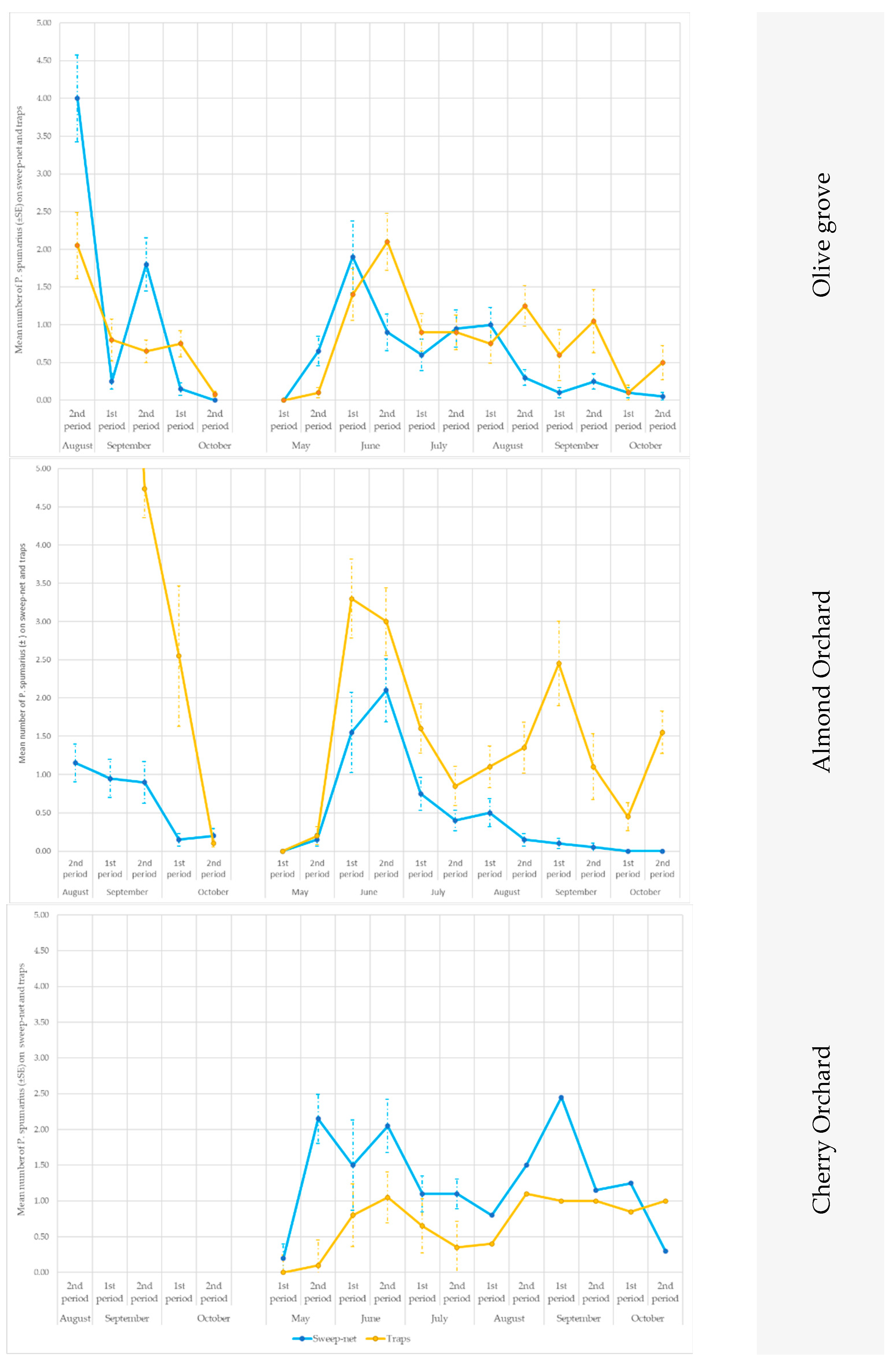
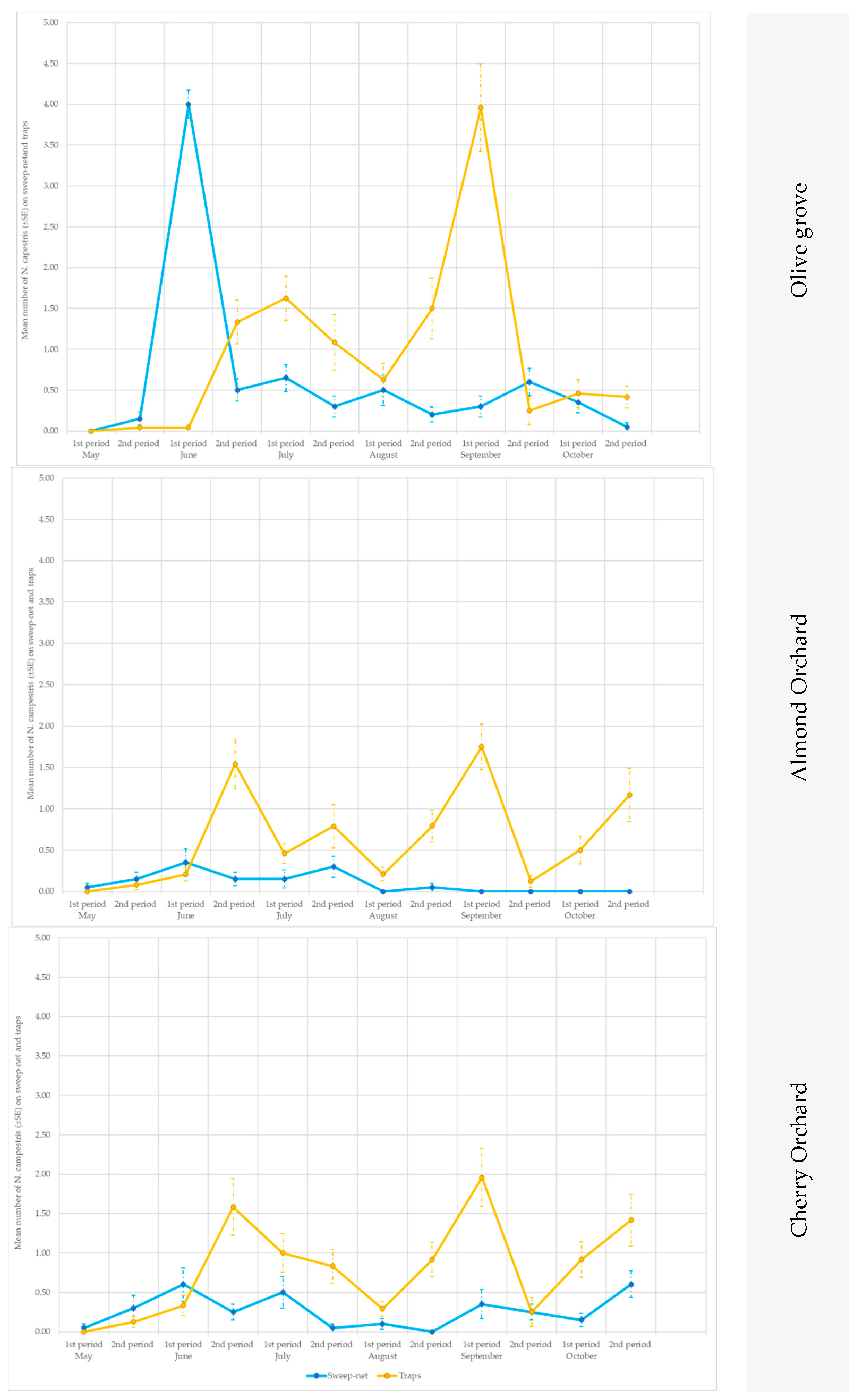
3.2. Evaluation of the Efficiency of the Different Yellow Sticky Traps in the Olive Groves, and in the Cherry and Almond Orchards
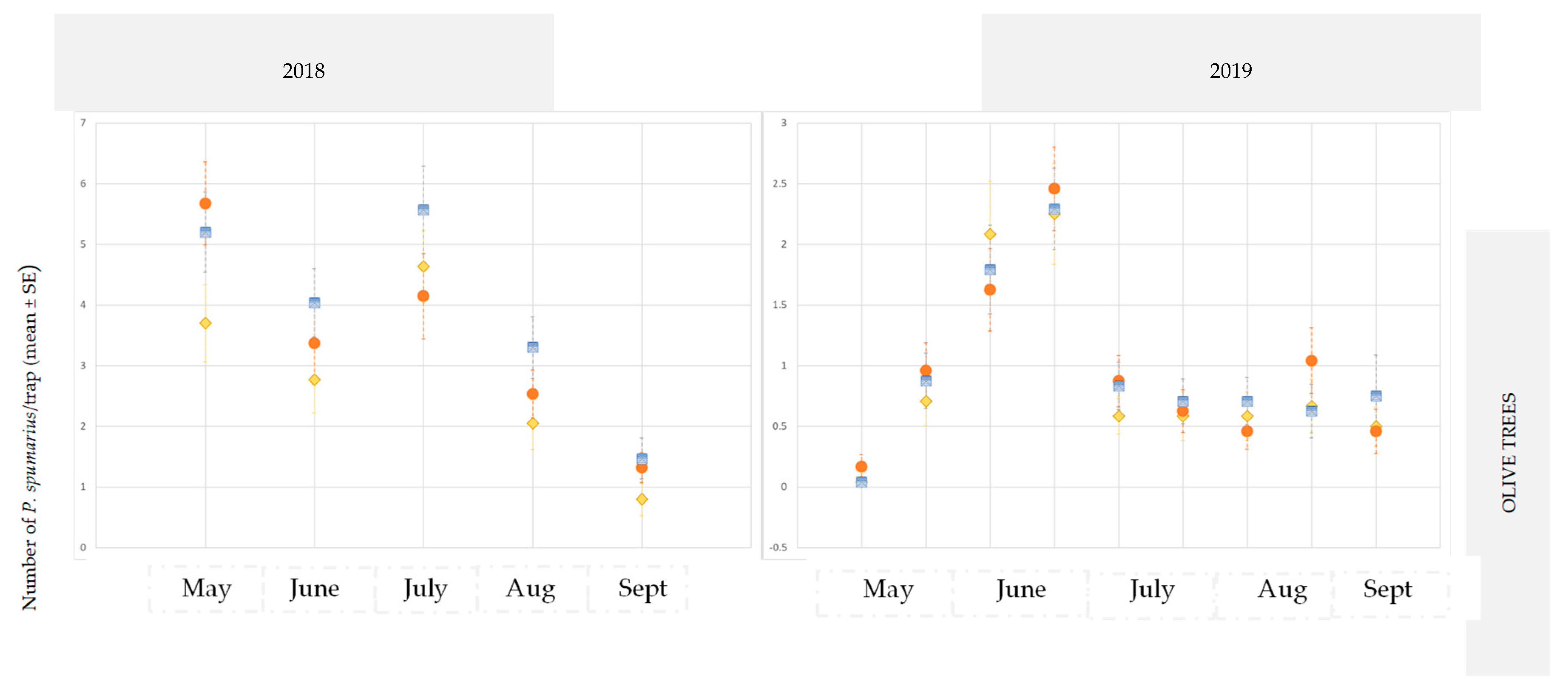
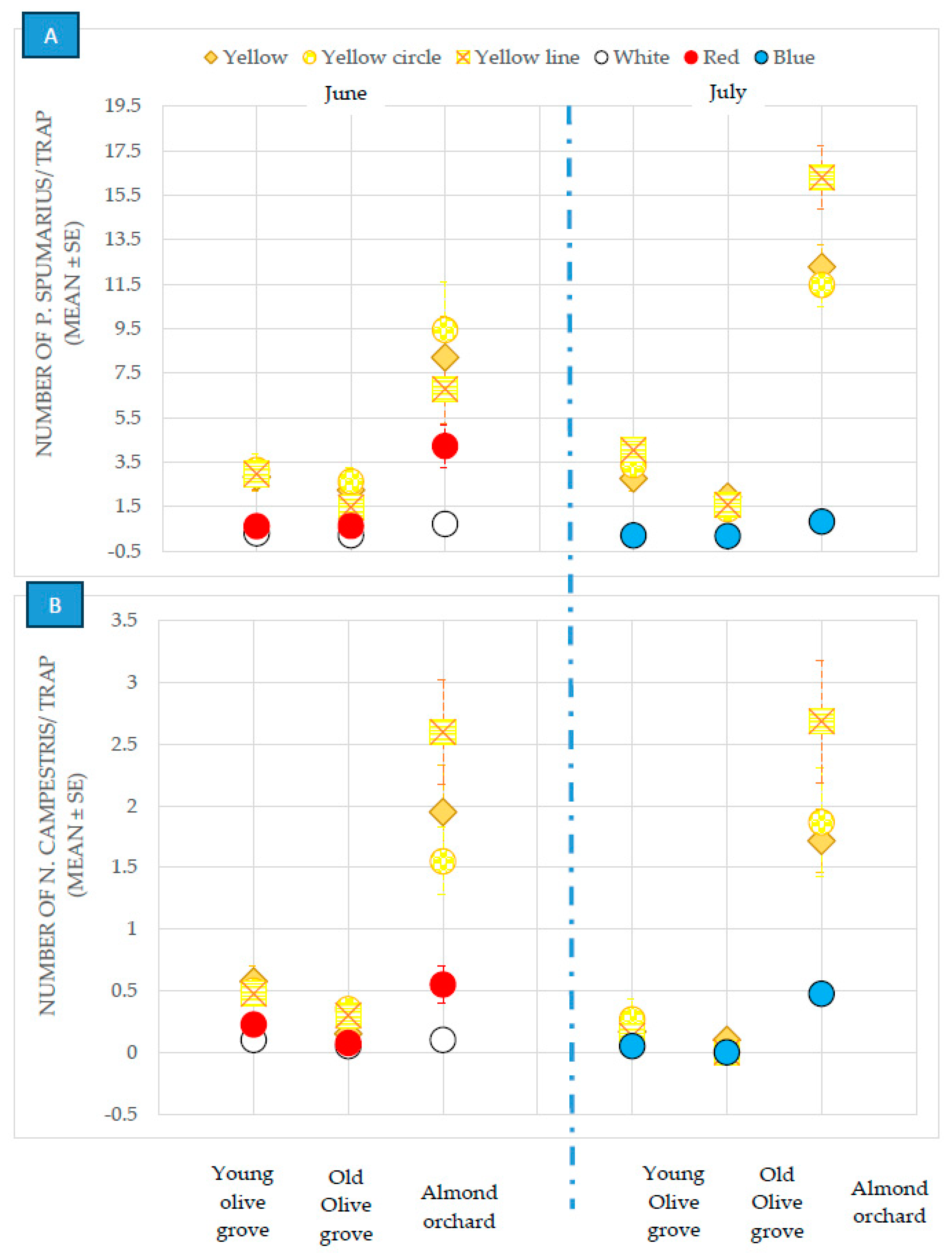
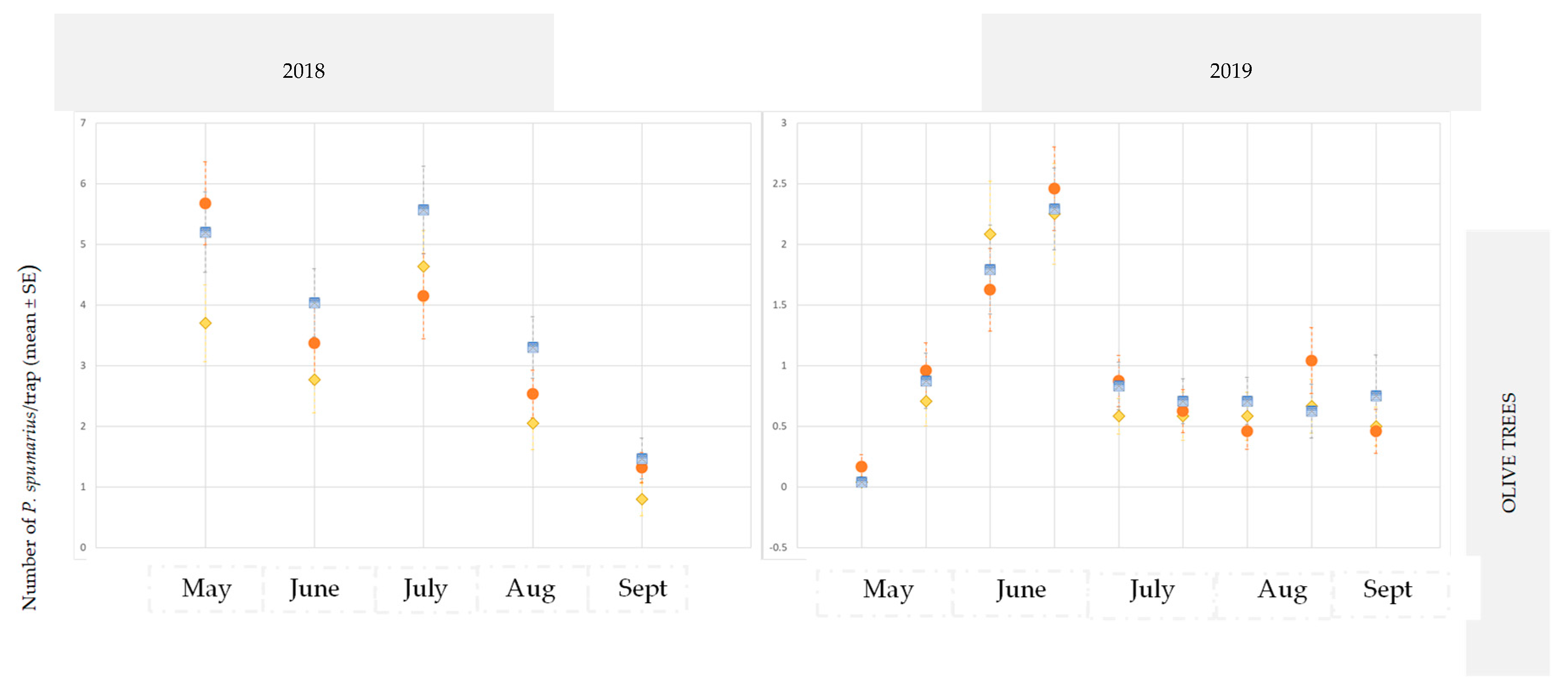
3.2.1. Olive Groves
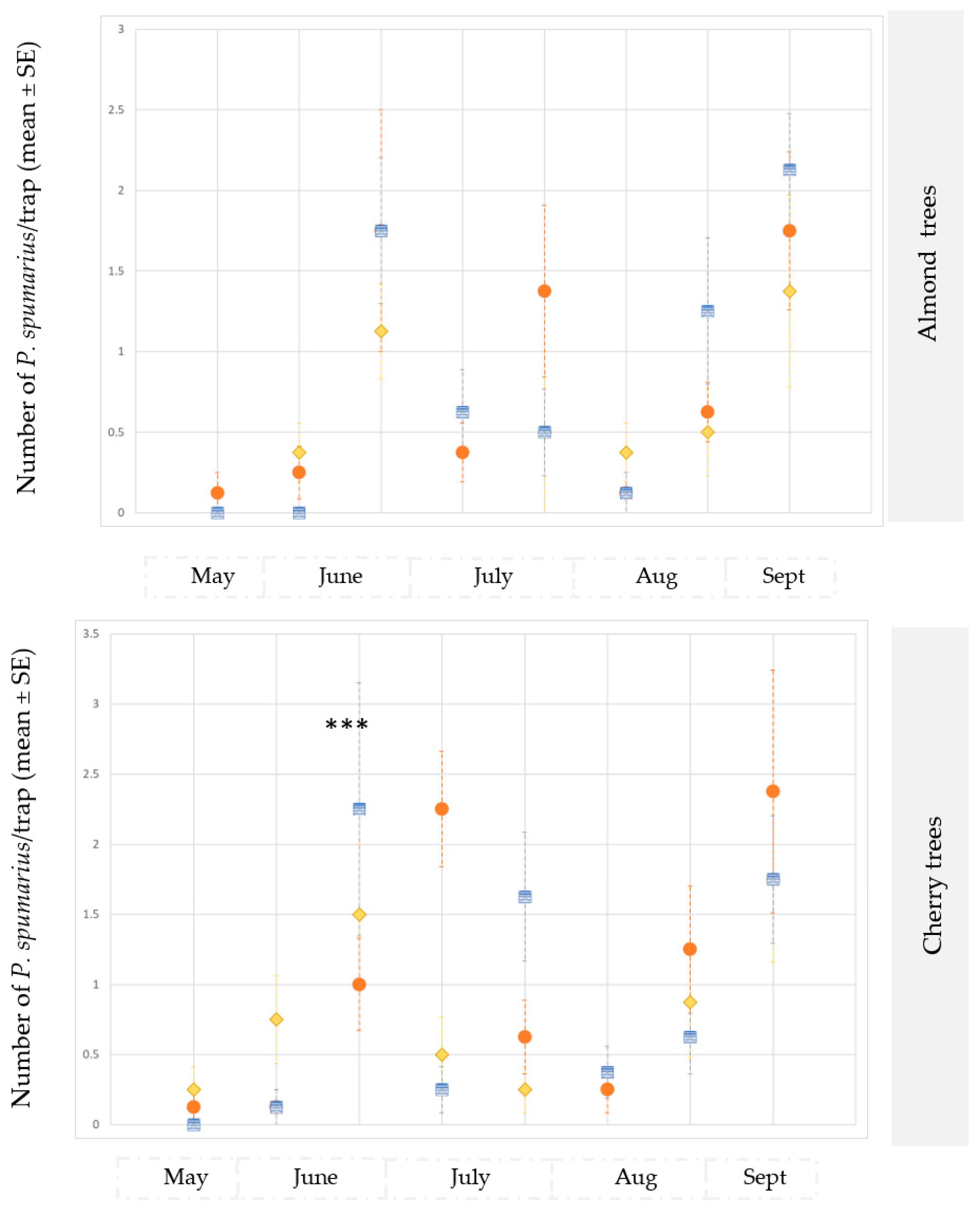
3.2.2. Almond and Cherry Orchards
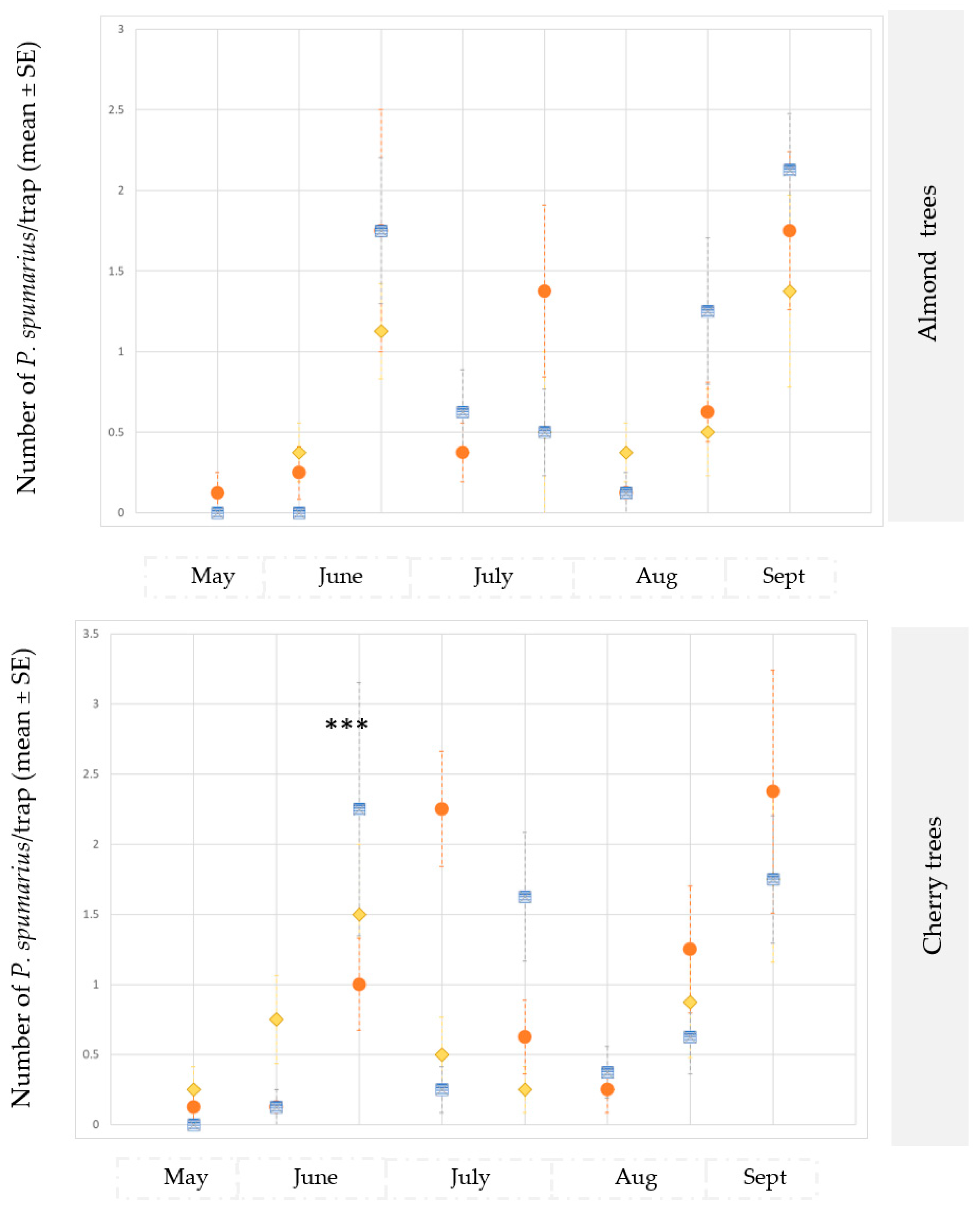
3.2.3. Occurrence of Non-Target Insect Species on the Traps
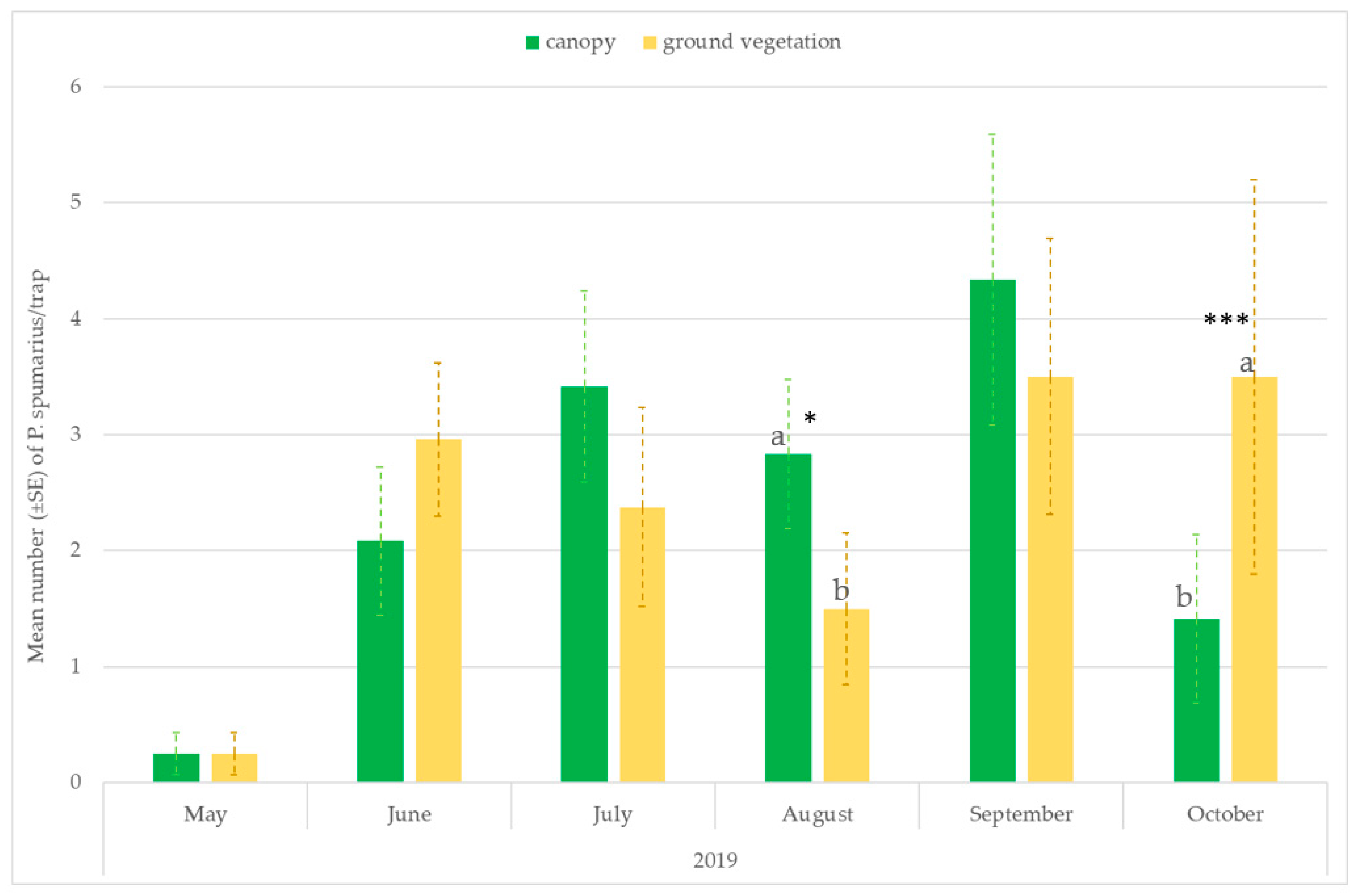
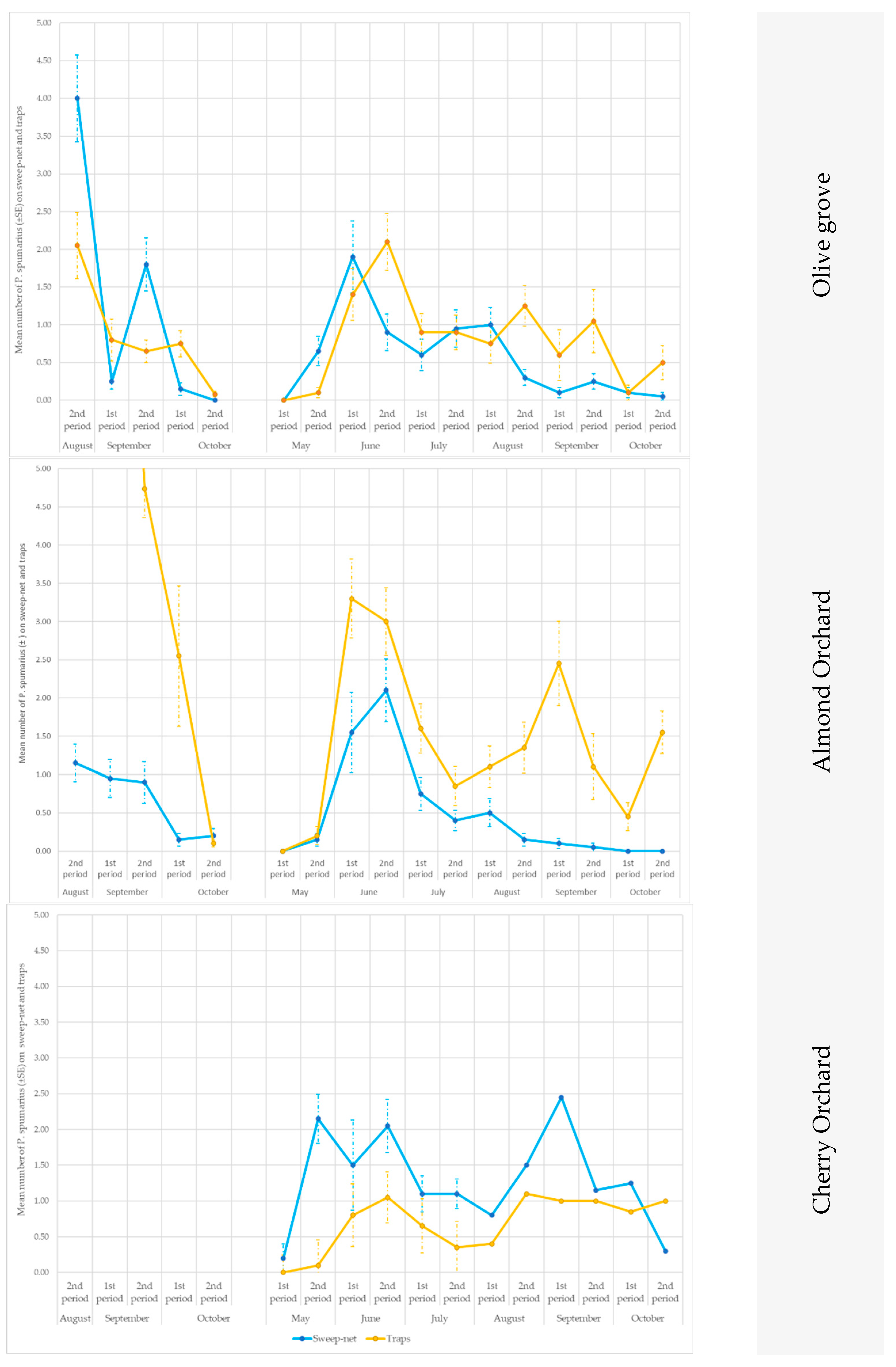
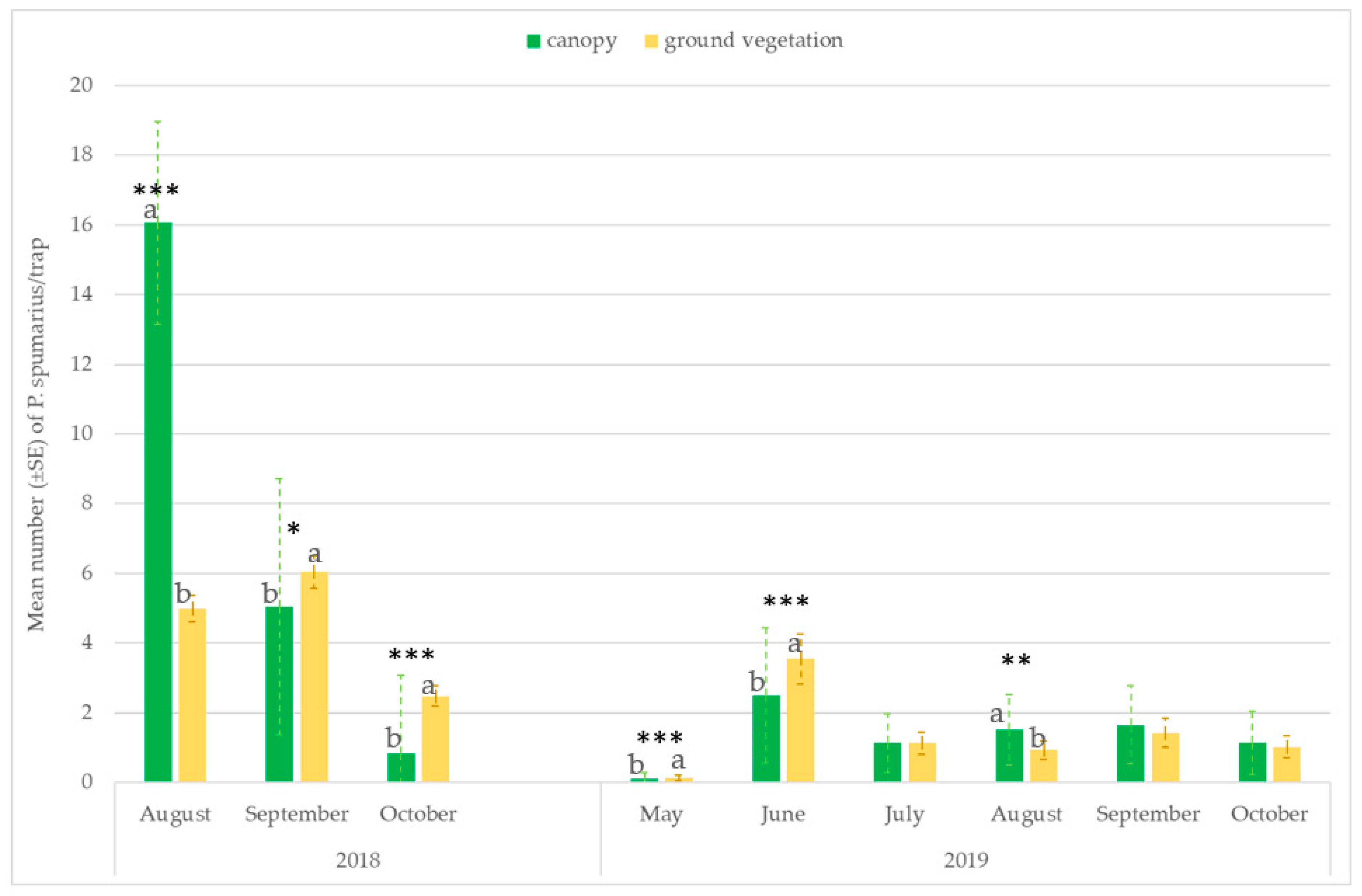
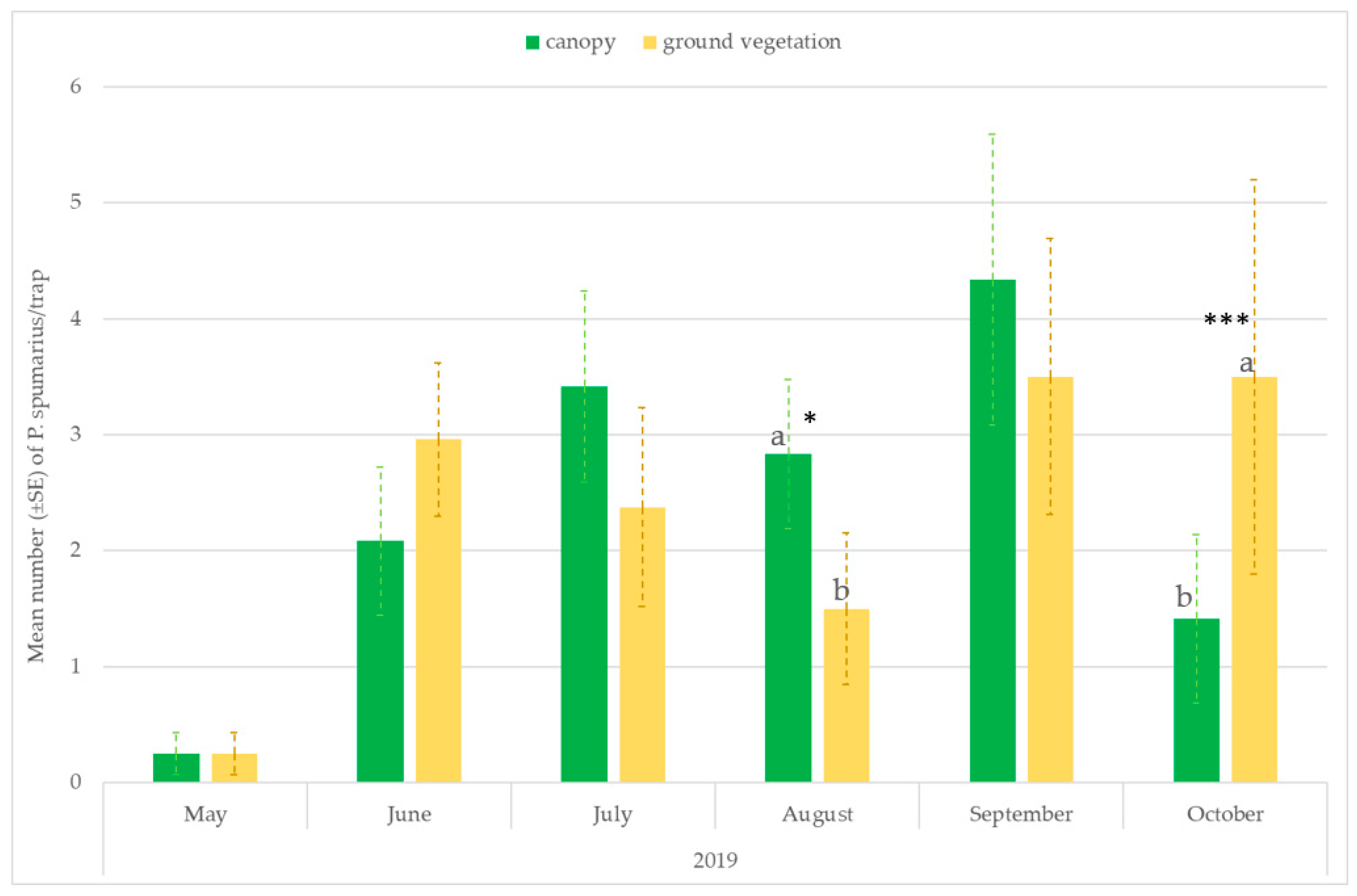
3.3. Monitoring Vectors in the Different Vegetation Compartments
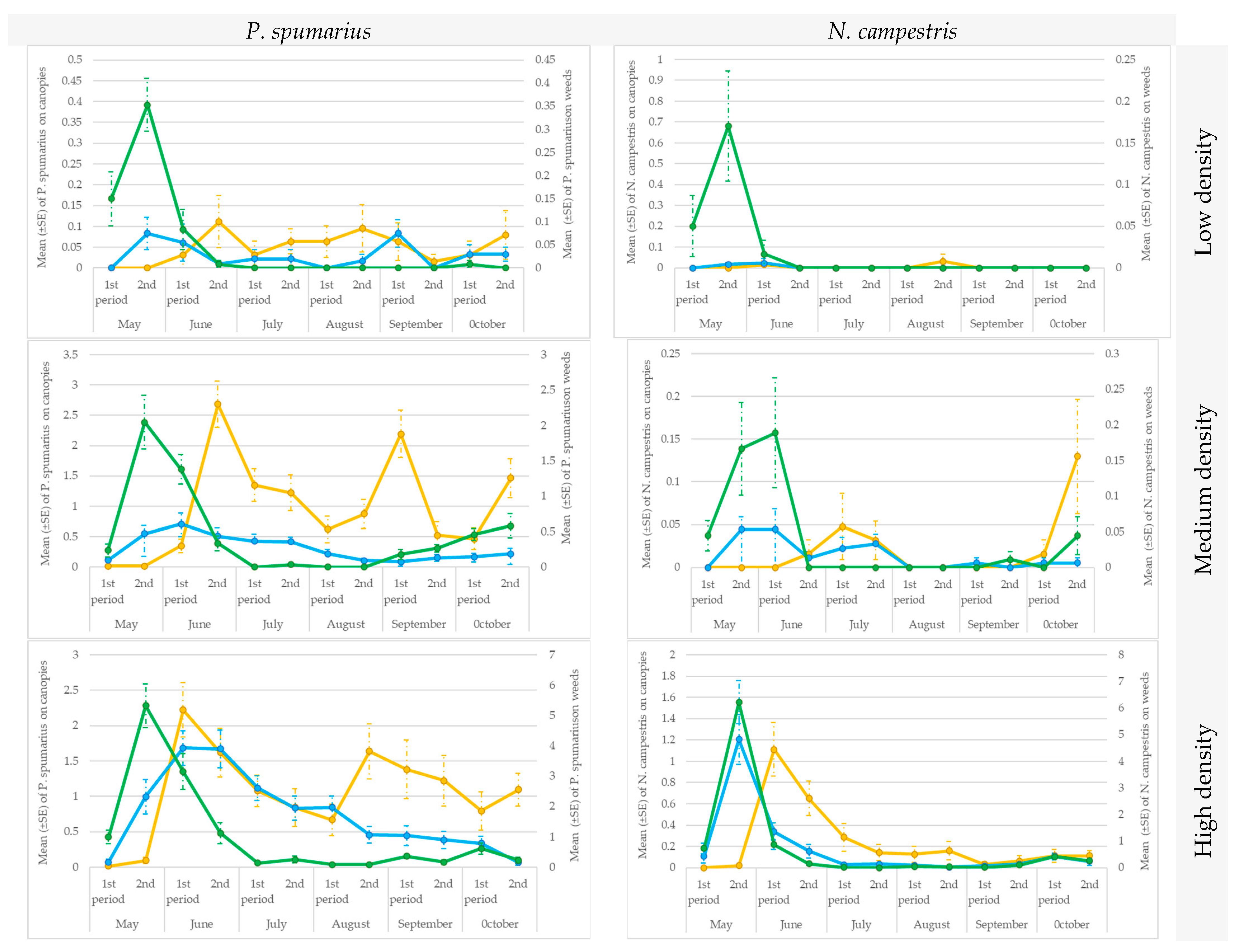
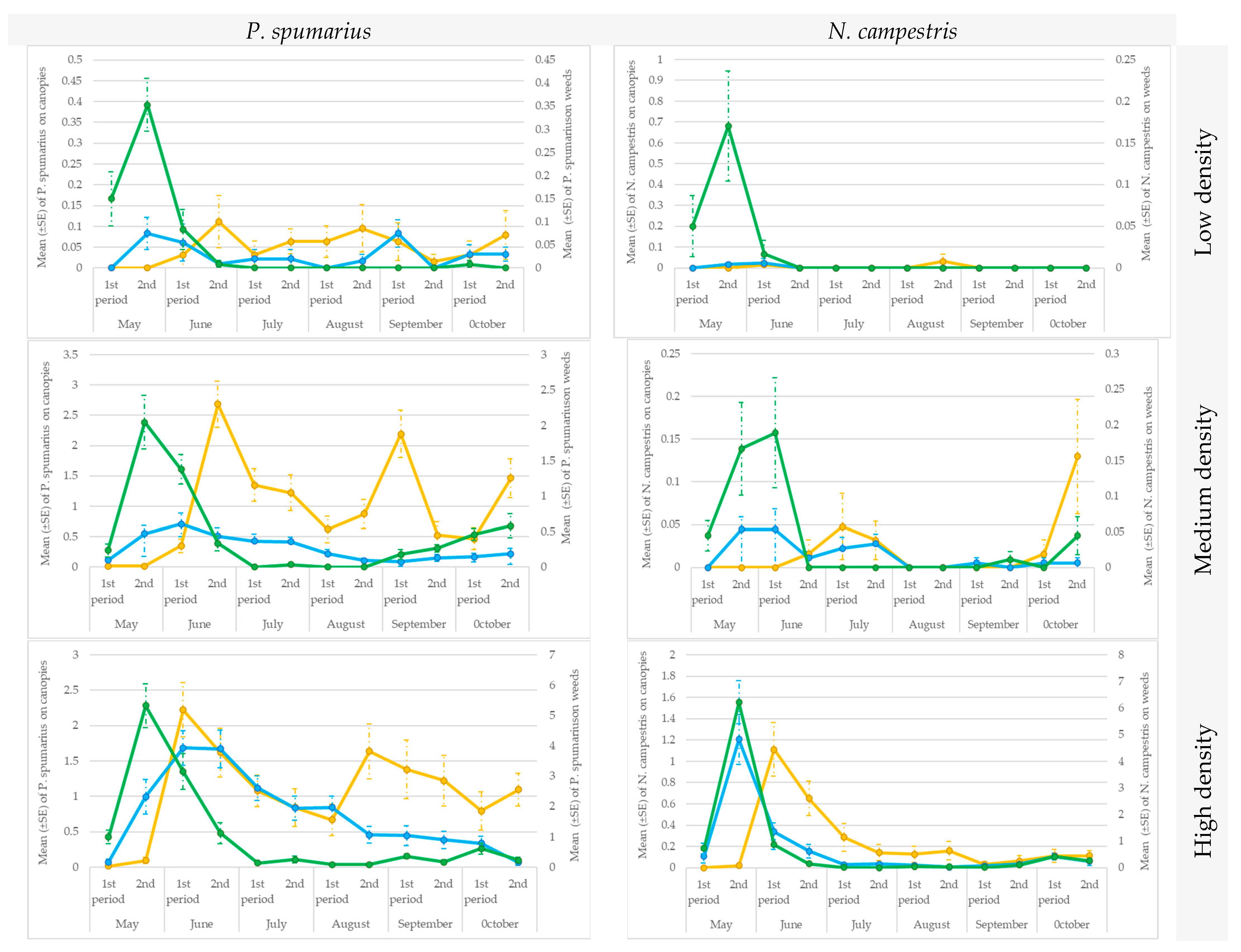
3.4. Testing the Traps in the Sites Characterized by Different Spittlebug Density Populations
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saponari, M.; Boscia, D.; Nigro, F.; Martelli, G.P. Identification of DNA sequences related to Xylella fastidiosa in oleander, almond and olive trees exhibiting leaf scorch symptoms in Apulia (Southern Italy). J. Plant Pathol. 2013, 95, 688. [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health). Scientific Opinion on the risks to plant health posed by Xylella fastidiosa in the EU territory, with the identification and evaluation of risk reduction options. EFSA J. 2015, 13, 3989. [Google Scholar] [CrossRef]
- EFSA PLH Panel (EFSA Panel on Plant Health). Updated pest categorization of Xylella fastidiosa. EFSA J. 2018, 16, 5357. [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health). Update of the Scientific Opinion on the risks to plant health posed by Xylella fastidiosa in the EU territory. EFSA J. 2019, 17, e05665. [Google Scholar]
- Giampetruzzi, A.; Loconsole, G.; Zicca, S.; Boscia, D.; Balestra, G.M.; Saponari, M. Draft Genome Sequence Resource of Xylella fastidiosa Strain Alm_Lz_1 Associated with a New Outbreak in Lazio, Italy. Phytopathology 2023, 113, 108–111. [Google Scholar] [CrossRef]
- Bodino, N.; Cavalieri, V.; Dongiovanni, C.; Saponari, M.; Bosco, D. Bioecological traits of spittlebugs and their implications on the epidemiology and control of Xylella fastidiosa epidemic in Apulia (Southern Italy). Phytopathology 2023. Online ahead of print. [Google Scholar] [CrossRef]
- Cornara, D.; Morente, M.; Markheiser, A.; Bodino, N.; Tsai, C.W.; Fereres, A.; Redak, R.A.; Perring, T.M.; Lopes, J.R.S. An overview on the worldwide vectors of Xylella fastidiosa. Entomol. Gen. 2019, 39, 157–181. [Google Scholar] [CrossRef]
- Desprez-Loustau, M.L.; Balci, Y.; Cornara, D.; Gonthier, P.; Robin, C.; Jacques, M.A. Is Xylella fastidiosa a serious threat to European forests? Forestry 2021, 94, 1–17. [Google Scholar] [CrossRef]
- Rösch, V.; Marques, E.; Miralles-Núñez, A.; Zahniser, J.; Wilson, M. Draeculacephala robinsoni Hamilton, 1967 (Hemiptera: Auchenorryncha: Cicadellidae), a newly introduced species and genus in Europe with comments on its identification. Zootaxa 2022, 5113, 439–448. [Google Scholar] [CrossRef]
- Cornara, D.; Saponari, M.; Zeilinger, A.R.; de Stradis, A.; Boscia, D.; Loconsole, G.; Bosco, D.; Martelli, G.P.; Almeida, R.P.P.; Porcelli, F. Spittlebugs as vectors of Xylella fastidiosa in olive orchards in Italy. J. Pest Sci. 2017, 90, 521–530. [Google Scholar] [CrossRef]
- Cruaud, A.; Gonzalez, A.; Godefroid, M.; Nidelet, S.; Streito, J.; Thuillier, J.; Rossi, J.P.; Santoni, S.; Rasplus, J. Using insects to detect, monitor and predict the distribution of Xylella fastidiosa: A case study in Corsica. Sci. Rep. 2018, 8, 15628. [Google Scholar] [CrossRef]
- Olmo, D.; Nieto, A.; Adrover, F.; Urbano, A.; Beidas, O.; Juan, A.; Noales, M.; Lόpez, M.M.; Navarro, I.; Monteverde, A.; et al. First detection of Xylella fastidiosa infecting cherry (Prunus avium) and Polygala myrtifolia plants, in Mallorca Island, Spain. Plant Dis. 2017, 101, 1820. [Google Scholar] [CrossRef]
- Saponari, M.; Loconsole, G.; Cornara, D.; Yokomi, R.K.; Stradis, A.D.; Boscia, D.; Bosco, D.; Martelli, G.P.; Krugner, R.; Porcelli, F. Infectivity and transmission of Xylella fastidiosa by Philaenus spumarius (Hemiptera: Aphrophoridae) in Apulia, Italy. J. Econ. Entomol. 2014, 107, 1316–1319. [Google Scholar] [CrossRef]
- Cavalieri, V.; Altamura, G.; Fumarola, G.; Di Carolo, M.; Saponari, M.; Cornara, D.; Bosco, D.; Dongiovanni, C. Transmission of Xylella fastidiosa Subspecies pauca Sequence Type 53 by different Insects Species. Insects 2019, 10, 324. [Google Scholar] [CrossRef]
- Ben Moussa, I.E.; Mazzoni, V.; Valentini, F.; Yasseen, T.; Lorusso, D.; Speranza, S.; Digiaro, M.; Varvaro, L.; Krugher, R.; D’Onghia, A.M. Seasonal fluctuations of sap-feeding insect species infected by Xylella fastidiosa in Apulia olive groves of southern Italy. J. Econ. Entomol. 2016, 109, 1512–1518. [Google Scholar] [CrossRef]
- Cornara, D.; Cavalieri, V.; Dongiovanni, C.; Altamura, G.; Palmisano, F.; Bosco, D.; Porcelli, F.; Almeida, R.P.P.; Saponari, M. Transmission of Xylella fastidiosa by naturally infected Philaenus spumarius (Hemiptera, Aphrophoridae) to different host plants. J. Appl. Entomol. 2017, 141, 80–87. [Google Scholar] [CrossRef]
- Bodino, N.; Cavalieri, V.; Dongiovanni, C.; Plazio, E.; Saladini, M.A.; Volani, S.; Simonetto, A.; Fumarola, G.; Di Carolo, M.; Porcelli, F.; et al. Phenology, seasonal abundance and stage-structure of spittlebug (Hemiptera: Aphrophoridae) populations in olive groves in Italy. Sci. Rep. 2019, 9, 17725. [Google Scholar] [CrossRef]
- Cornara, D.; Bosco, D.; Fereres, A. Philaenus spumarius: When an old acquaintance becomes a new threat to European agriculture. J. Pest Sci. 2018, 91, 957–972. [Google Scholar] [CrossRef]
- Antonatos, S.; Papachristos, D.; Kapantaidaki, D.; Lytra, I.C.; Varikou, K.; Evangelou, V.; Milonas, P. Presence of Cicadomorpha in olive orchards of Greece with special reference to Xylella fastidiosa vectors. J. Appl. Entom. 2019, 144, 1–11. [Google Scholar] [CrossRef]
- Di Serio, F.; Bodino, N.; Cavalieri, V.; Demichelis, S.; Di Carolo, M.; Dongiovanni, C.; Fumarola, G.; Gilioli, G.; Guerrieri, E.; Picciotti, U.; et al. Collection of data and information on biology and control of vectors of Xylella fastidiosa. EFSA Support. Publ. 2019, 16, 1628E. [Google Scholar] [CrossRef]
- Vos, S.; Camilleri, M.; Diakaki, M.; Lázaro, E.; Parnell, S.; Schenk, M.; Schrader, G.; Vicent, A. Pest survey card on Xylella fastidiosa. EFSA Support. Publ. 2019, 16, 1667E. [Google Scholar]
- Purcell, A.H.; Gravena, S.; Donadio, L.C. Sharpshooter in citrus crops. In Citrus-Integrated Management of Insect and Mite Pests. Bebedouro, Estaçao Experimental de Citricultura; 1994; pp. 213–229. [Google Scholar]
- Morente, M.; Cornara, D.; Plaza, M.; Durán, J.M.; Capiscol, C.; Trillo, R.; Ruiz, M.; Ruz, C.; Sanjuan, S.; Pereira, J.A.; et al. Distribution and relative abundance of insect vectors of Xylella fastidiosa in olive groves of the Iberian Peninsula. Insects 2018, 9, 175. [Google Scholar] [CrossRef]
- Weaver, C.R.; King, D.R. Meadow spittlebug Philaenus leucophthalmus (L.). Ohio Agric. Exp. Stn. Res. Bull. 1954, 741, 1–99. [Google Scholar]
- Lavigne, B. Biology of Philaenus leucophthalmus (L.), in Massachusetts. J. Econ. Entomol. 1959, 52, 904–907. [Google Scholar] [CrossRef]
- Wilson, M.C.; Shade, R.E. Relative attractiveness of various luminescent colors to the cereal leaf beetle and the meadow spittle bug. J. Econ. Entomol. 1967, 60, 578–580. [Google Scholar] [CrossRef]
- Novotny, V. Vertical-distribution of leafhoppers (Hemiptera, Auchenorrhyncha) within a meadow community. Acta Entomol. Bohemoslov. 1992, 89, 13–20. [Google Scholar]
- Pavan, F. Occurrence on elm and phenology of Auchenorrhyncha potential vectors of the phytoplasma associated with elm yellows disease. Boll. di Zool. Agr. Bachic. 2000, 32, 59–68. [Google Scholar]
- Bleicher, K.; Orosz, A.; Cross, J.; Markό, V. Survey of leafhoppers, planthoppers and froghoppers (Auchenorrhyncha) In apple orchards in South-East England. Acta Phytopathol. Entomol. Hung. 2010, 45, 93–105. [Google Scholar] [CrossRef]
- Krysan, J.L.; Horton, D.R. Seasonality of catch of pear psylla Cacopsylla pyricola (Homoptera: Psyllidae) on yellow traps. Environ. Entomol. 1991, 20, 626–634. [Google Scholar] [CrossRef]
- Sètamou, M.; Sanchez, A.; Saldana, R.R.; Patt, J.M.; Summy, R. Visual responses of adult Asian citrus psyllid (Hemiptera: Liviidae) to colored sticky traps on citrus trees. J. Insect Behav. 2014, 27, 540–553. [Google Scholar] [CrossRef]
- Berlinger, M.J. A yellow sticky trap for whiteflies: Trialeurodes vaporarorium and Bemisia tabaci (Aleurodidae). Entomol. Exp. Appl. 1980, 27, 98–102. [Google Scholar] [CrossRef]
- Gerling, D.; Horowitz, A.R. Yellow traps for evaluation the population levels and dispersal patterns of Bemisia tabaci (Gennadius) (Homoptera: Aleyrodidae). Ann. Entomol. Soc. Am. 1984, 77, 753–759. [Google Scholar] [CrossRef]
- Aldini, R.N.; Ciampitti, M.; Cravedi, P. Monitoring the leafhopper Scaphoideus titanus Ball and the planthopper Hyalesthes obsoletus Signoret in Northern Italy. IOBC WPRS Bull. 2003, 26, 233–236. [Google Scholar]
- Matsukura, K.; Yoshida, K.; Matsumura, M. Efficient monitoring of maize orange leafhopper, Cicadulina bipuncata (Hemiptera: Cicadellidae), and small brown planthopper, Laodelphax striatellus (Hemiptera: Delphacidae), in forage maize fields using yellow sticky traps. Appl. Entomol. Zool. 2011, 46, 585–591. [Google Scholar] [CrossRef]
- Bodino, N.; Cavalieri, V.; Dongiovanni, C.; Saladini, M.A.; Simonetto, A.; Volani, S.; Plazio, E.; Altamura, G.; Tauro, D.; Gilioli, G.; et al. Spittlebugs of Mediterranean olive groves: Host-plant exploitation throughout the year. Insects 2020, 11, 130. [Google Scholar] [CrossRef]
- Biedermann, R.; Niedringhaus, R. The Plant-and Leafhoppers of Germany-Identification Key to All Species; WABV Fründ: Bremen, Germany, 2009; pp. 1–409. [Google Scholar]
- Della Giustina, W. Homoptéres Cicadellidae; Faune de France: Paris, France, 1989; pp. 1–350. [Google Scholar]
- Holzinger, W.; Kammerlander, I.; Nickel, H. The Auchenorrhyncha of Central Europe. Die Zikaden Mitteleuropas: Fulgoromorpha, Cicadomorpha excl. Cicadellidae; Brill Leiden: Boston, MA, USA, 2003; pp. 1–673. [Google Scholar]
- Ribaut, H. Homoptères Auchénorhynques II (Jassidae). In Faune de France; Lechevalier: Paris, France, 1952; Volume 57, pp. 1–477. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Tukey, J. Comparing Individual Means in the Analysis of Variance. Biometrics 1949, 5, 99–114. [Google Scholar] [CrossRef]
- Bullas-Appleton, E.S.; Otis, G.; Gillard, G.; Schaafsma, A. Potato leafhopper (Homoptera: Cicadellidae) varietal preferences in edible beans in relation to visual and olfactory cues. Environ. Entomol. 2004, 33, 1381–1388. [Google Scholar] [CrossRef]
- Mazzoni, V.; Trona, F.; Ioriatti, C.; Lucchi, A.; Eriksson, A.; Anfora, G. Attractiveness of different colours to Scaphoideus titanus Ball (Hemiptera: Cicadellidae) adults. IOBC Bull. 2011, 67, 281–284. [Google Scholar]
- Patt, J.M.; Sétamou, M. Olfactory and visual stimuli affecting host plant detection in Homolodisca coagulata (Hemiptera: Cicadellidae). Environ. Entomol. 2007, 36, 142–150. [Google Scholar] [CrossRef]
- Todd, J.L.; Harris, M.O.; Nault, L.R. Importance of color stimuli in host-finding by Dalbulus leafhoppers. Entomol. Exp. Appl. 1990, 53, 245–255. [Google Scholar] [CrossRef]
- Todd, J.L.; Phelan, P.L.; Nault, L.R. Interaction between visual and olfactory stimuli during host-finding by leafhopper, Dalbulus maidis (Homoptera: Cicadellidae). J. Chem. Ecol. 1990, 16, 2121–2133. [Google Scholar] [CrossRef]
- van der Kooi, C.J.; Stavenga, D.G.; Arikawa, K.; Belušic, G.; Kelber, A. Evolution of Insect Color Vision: From Spectral Sensitivity to Visual Ecology. Annu. Rev. Entomol. 2021, 66, 435–461. [Google Scholar] [CrossRef]
- Nilakhe, S.S.; Buainain, C.M. Observations on movement of spittlebug adults. Pesq. Agropec. Bras. 1988, 23, 123–134. [Google Scholar]
- Johnson, M.P.; Freytag, P.H. Metallic wood borers (Coleoptera: Buprestidae) and spittlebugs (Homoptera: Cercopidae) on Pin Oak in Kentucky. J. Kans. Entomol. Soc. 2001, 74, 28–31. [Google Scholar]
- Avramov, Z.; Ivanova, I.; Laginova, M. Screening for phytoplasma presence in leafhoppers and planthoppers collected in Bulgarian vineyards. Bull. Insectology 2011, 64, 115–116. [Google Scholar]
- Zhang, J.; Lashomb, J.; Gould, A.; Hamilton, G. Cicadomorpha Insects Associated with Bacterial Leaf Scorch Infected Oak in Central New Jersey. Environ. Entomol. 2011, 40, 1131–1143. [Google Scholar] [CrossRef]
- Dellapé, G.; Bouvet, J.P.; Paradell, S.L. Diversity of Cicadomorpha (Hemiptera: Auchenorrhyncha) in Citrus Orchards in Northeastern Argentina. Florida Entomol. 2013, 96, 1125–1134. [Google Scholar] [CrossRef]
- Elbeaino, T.; Yaseen, T.; Valentini, F.; Moussa, I.E.B.; Mazzoni, V.; Onghia, A.M. Identification of three potential insect vectors of Xylella fastidiosa in southern Italy. Phytopathol. Mediterr. 2014, 53, 328–332. [Google Scholar]
- Yaseen, T.; Drago, S.; Valentini, F.; Elbeaino, T.; Stampone, G.; Digiaro, M.; D’Onghia, A.M. On-site detection of Xylella fastidiosa in host plants and in “spy insects” using the real-time loop-mediated isothermal amplification method. Phytopathol. Mediterr. 2015, 54, 488–496. [Google Scholar]
- Santoiemma, G.; Tamburini, G.; Sanna, F.; Mori, N.; Marini, L. Landscape composition predicts the distribution of Philaenus spumarius, vector of Xylella fastidiosa, in olive groves. J. Pest Sci. 2019, 92, 1101–1109. [Google Scholar] [CrossRef]
- Sanna, F.; Mori, N.; Santoiemma, G.; D’Ascenzo, D.; Scotillo, M.A.; Marini, L. Ground cover management in olive groves reduces populations of Philaenus spumarius (Hemiptera: Aphrophoridae), vector of Xylella fastidiosa. J. Econ. Entomol. 2021, 114, 1716–1721. [Google Scholar] [CrossRef] [PubMed]
- López-Mercadal, J.; Delgado, S.; Mercadal, P.; Seguí, G.; Lalucat, J.; Busquets, A.; Gomila, M.; Lester, K.; Kenyon, D.M.; Ruiz-Pérez, M.; et al. Collection of data and information in Balearic Islands on biology of vectors and potential vectors of Xylella fastidiosa (GP/EFSA/ALPHA/017/01). EFSA Support. Publ. 2021, 18, 6925E. [Google Scholar] [CrossRef]
- Beal, D.J.; Cooper, M.; Daugherty, M.P.; Purcell, A.H.; Almeida, R.P. Seasonal abundance and infectivity of Philaenus spumarius (Hemiptera: Aphrophoridae), a vector of Xylella fastidiosa in California vineyards. Environ. Entomol. 2021, 50, 467–476. [Google Scholar] [CrossRef]
- Daugherty, M.P.; Lopes, J.; Almeida, R.P. Vector within-host feeding preference mediates transmission of a heterogeneously distributed pathogen. Ecol. Entomol. 2010, 35, 360–366. [Google Scholar] [CrossRef]
- Mazzoni, V. Contribution to the knowledge of the Auchenorrhyncha (Hemiptera Fulgoromorpha and Cicadomorpha) of Tuscany (Italy). Redia 2005, 88, 85–102. [Google Scholar]
- Lopes, J.R.; Landa, B.B.; Fereres, A. A survey of potential insect vectors of the plant pathogenic bacterium Xylella fastidiosa in three regions of Spain. Span. J. Agric. Res. 2014, 12, 795–800. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traps | Mean Number of Spittlebugs (±SE) | |||||
| June 2018 | ||||||
| P. spumarius | N. campestris | |||||
| 20 Year-Old Olive Grove | Century-Old Olive Grove | Almond Orchard | 20 Year-Old Olive Grove | Century-Old Olive Grove | Almond Orchard | |
| White | 0.2 ± 0.06 b | 0.1 ± 0.04 b | 1.1 ± 0.16 b | 0.1 ± 0.05 b | 0.05 ± 0.03 a | 0.2 ± 0.06 c |
| Red | 0.75 ± 0.13 b | 0.6 ± 0.14 b | 3.4 ± 0.94 b | 0.3 ± 0.09 ab | 0.08 ± 0.04 a | 0.7 ± 0.15 bc |
| Yellow | 3.7 ± 0.63 a | 2.6 ± 0.50 a | 13 ± 1.83 a | 0.6 ± 0.12 a | 0.15 ± 0.06 a | 2.2 ± 0.38 a |
| Yellow with a circle-pattern | 5.7 ± 0.72 a | 3.3 ± 0.60 a | 14.3 ± 2.16 a | 0.5 ± 0.10 a | 0.35 ± 0.09 a | 1.6 ± 0.27 ab |
| Yellow with a line-pattern | 5.2 ± 0.68 a | 2.3 ± 0.33 a | 14.8 ± 1.55 a | 0.5 ± 0.09 a | 0.30 ± 0.08 a | 2.6 ± 0.42 a |
| F-value | 31.27 | 17.14 | 19.02 | 5.46 | 1.22 | 15.2 |
| p-value | <0.00001 | <0.00001 | <0.00001 | 0.0006 | 0.2500 | <0.00001 |
| Traps | July 2018 | |||||
| P. spumarius | N. campestris | |||||
| 20 Year-Old Olive Grove | Century-Old Olive Grove | Almond Orchard | 20 Year-Old Olive Grove | Century-Old Olive Grove | Almond Orchard | |
| Blue | 0.2 ± 0.05 b | 0.2 ± 0.06 b | 0.8 ± 0.12 c | 0.05 ± 0.03 a | 0 ± 0 a | 0.5 ± 0.10 b |
| Yellow | 2.8 ± 0.54 a | 1.9 ± 0.43 a | 12.3 ± 0.96 ab | 0.2 ± 0.08 a | 0.1 ± 0.06 a | 1.8 ± 0.26 ab |
| Yellow with a circle-pattern | 3.4 ± 0.69 a | 1.4 ± 0.29 ab | 11.5 ± 0.96 b | 0.3 ± 0.16 a | 0 ± 0 a | 1.9 ± 0.44 ab |
| Yellow with a line-pattern | 4.0 ± 0.56 a | 1.6 ± 0.29 ab | 16.3 ± 1.40 a | 0.1 ± 0.07 a | 0 ± 0 a | 2.8 ± 0.50 a |
| F-value | 10.58 | 5.74 | 40.84 | 0.33 | 2.11 | 5.09 |
| p-value | <0.00001 | 0.0017 | <0.00001 | 0.8008 | 0.1089 | 0.0034 |
| Traps | 26 July 2018 | ||
| 20 Year-Old Olive Grove | Century-Old Olive Grove | Almond Orchard | |
| Yellow | 20.59 b | 7.90 a | 10.23 a |
| Yellow with a circle-pattern | 27.53 ab | 4.93 a | 4.40 a |
| Yellow with a line-pattern | 35.96 a | 5.72 a | 7.36 a |
| F-value | 4.85 | 1.81 | 0.99 |
| p-value | 0.013 | 0.1779 | 0.3806 |
| Traps | 31 May 2019 | ||
| 20 Year-Old Olive Grove | Almond Orchards | Cherry Orchard | |
| Yellow | 13.92 a | 1.67 b | 0.46 a |
| Yellow with a circle-pattern | 10.25 a | 3.04 ab | 0.46 a |
| Yellow with a line-pattern | 9.63 a | 4.04 a | 0.29 a |
| F-value | 1.21 | 4.25 | 0.25 |
| p-value | 0.3093 | 0.0202 | 0.7830 |
| Host Plant | May | June | July | August | September | October |
|---|---|---|---|---|---|---|
| Site A—2018 | ||||||
| Olive trees | - | - | - | 2.0 ± 0.23 b | 1.7 ± 0.18 b | 1.9 ± 0.25 b |
| Almond trees | - | - | - | 19.0 ± 1.78 a | 20.4 ± 1.18 a | 4.8 ± 0.63 a |
| F-value | 411.27 | 538.18 | 53.65 | |||
| p-value | <0.00001 | <0.00001 | <0.00001 | |||
| Site A—2019 | ||||||
| Olive trees | 0.9 ± 0.19 b | 4.2 ± 0.58 b | 1.4 ± 0.22 b | 1.4 ± 0.27 b | 1.1 ± 0.37 b | 0.6 ± 0.15 b |
| Almond trees | 2.9 ± 0.39 a | 7.5 ± 0.63 a | 2.5 ± 0.28 a | 2.5 ± 0.30 a | 3.5 ± 0.45 a | 1.7 ± 0.23 a |
| Cherry trees | 0.8 ± 0.26 b | 2.2 ± 0.53 c | 1.0 ± 0.25 b | 1.2 ± 0.36 b | 1.5 ± 0.42 b | 1.4 ± 0.33 a |
| F-value | 42.45 | 49.23 | 19.43 | 11.92 | 10.63 | 11.05 |
| p-value | <0.00001 | <0.00001 | <0.00001 | <0.00001 | <0.00001 | <0.00001 |
| Site B—2019 | ||||||
| Olive trees | 0.04 ± 0.05 a | 1.4 ± 0.38 a | 2.7 ± 0.63 a | 2.1 ± 0.65 a | 3.8 ± 0.83 a | 0.9 ± 0.35 b |
| Almond trees | 0.08 ± 0.08 a | 1.8 ± 0.43 a | 1.3 ± 0.42 b | 1.0 ± 0.29 b | 1.9 ± 0.36 b | 1.7 ± 0.54 ab |
| Cherry trees | 0.12 ± 0.09 a | 1.9 ± 0.52 a | 1.8 ± 0.42 ab | 1.3 ± 0.35 ab | 2.3 ± 0.54 b | 2.3 ± 0.70 a |
| F-value | 0.53 | 0.79 | 5.42 | 3.64 | 5.51 | 5.79 |
| p-value | 0.59 | 0.46 | 0.0071 | 0.0329 | 0.0066 | 0.0052 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dongiovanni, C.; Di Carolo, M.; Fumarola, G.; Tauro, D.; Tedone, B.; Ancona, S.; Palmisano, V.; Carrieri, M.; Cavalieri, V. Comparing Different Sticky Traps to Monitor the Occurrence of Philaenus spumarius and Neophilaenus campestris, Vectors of Xylella fastidiosa, in Different Crops. Insects 2023, 14, 777. https://doi.org/10.3390/insects14090777
Dongiovanni C, Di Carolo M, Fumarola G, Tauro D, Tedone B, Ancona S, Palmisano V, Carrieri M, Cavalieri V. Comparing Different Sticky Traps to Monitor the Occurrence of Philaenus spumarius and Neophilaenus campestris, Vectors of Xylella fastidiosa, in Different Crops. Insects. 2023; 14(9):777. https://doi.org/10.3390/insects14090777
Chicago/Turabian StyleDongiovanni, Crescenza, Michele Di Carolo, Giulio Fumarola, Daniele Tauro, Biagio Tedone, Simona Ancona, Valentina Palmisano, Mauro Carrieri, and Vincenzo Cavalieri. 2023. "Comparing Different Sticky Traps to Monitor the Occurrence of Philaenus spumarius and Neophilaenus campestris, Vectors of Xylella fastidiosa, in Different Crops" Insects 14, no. 9: 777. https://doi.org/10.3390/insects14090777
APA StyleDongiovanni, C., Di Carolo, M., Fumarola, G., Tauro, D., Tedone, B., Ancona, S., Palmisano, V., Carrieri, M., & Cavalieri, V. (2023). Comparing Different Sticky Traps to Monitor the Occurrence of Philaenus spumarius and Neophilaenus campestris, Vectors of Xylella fastidiosa, in Different Crops. Insects, 14(9), 777. https://doi.org/10.3390/insects14090777