
Ovary Dissection Is a Sensitive Measure of Sterility in Anopheles gambiae Exposed to the Insect Growth Regulator Pyriproxyfen
 , ,
, ,  , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
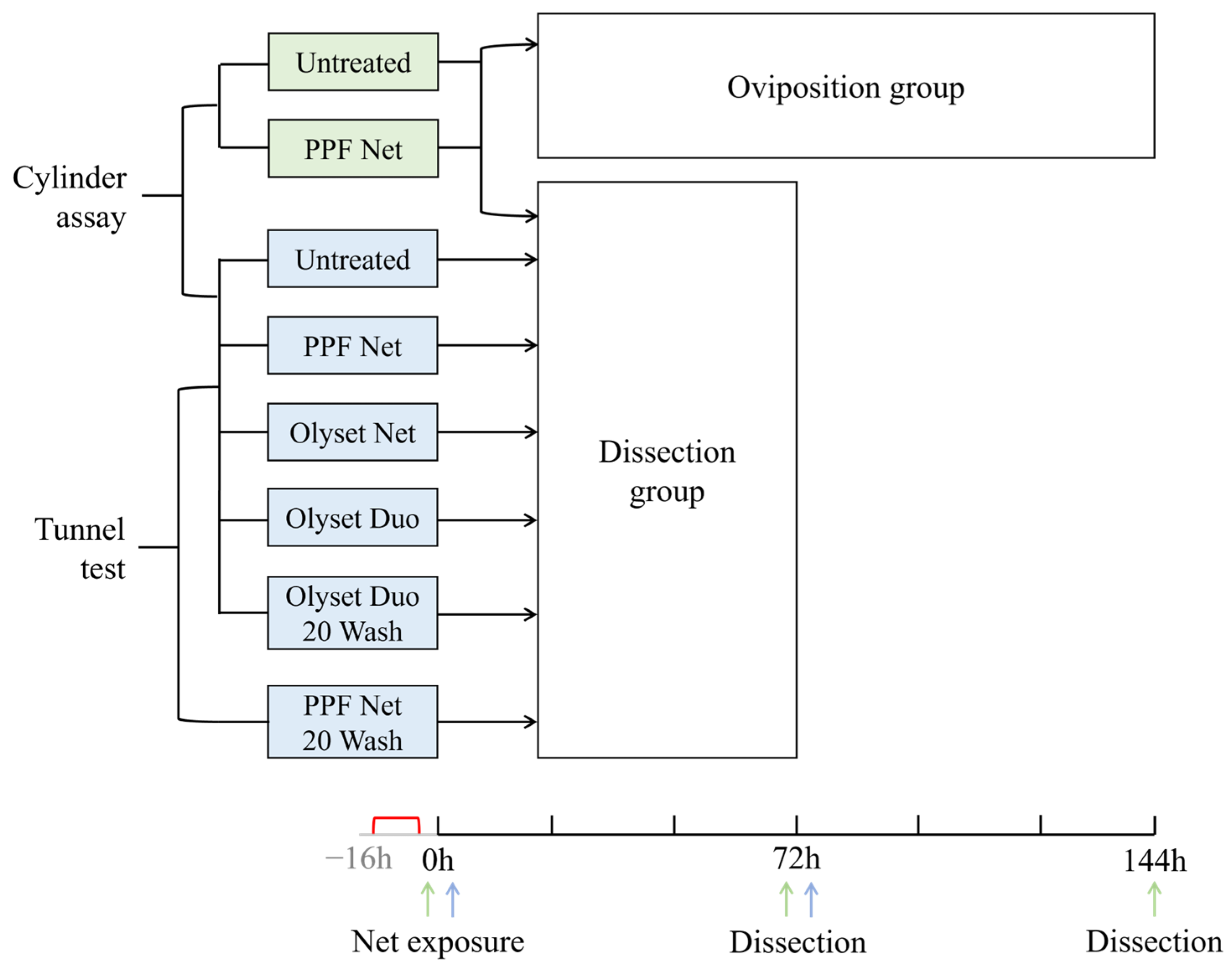
2. Materials and Methods
2.1. Mosquitoes
2.2. Test LLINs
- (1)
- Olyset® Duo (2% permethrin and 1% PPF), unwashed;
- (2)
- Olyset® Duo (2% permethrin and 1% PPF) with 20 washes (20W);
- (3)
- Olyset® Net (2% permethrin), unwashed;
- (4)
- Long-lasting PPF Net (1% PPF), unwashed;
- (5)
- Long-lasting PPF Net (1% PPF), 20W.
2.3. Bioassay Procedures
2.4. Oviposition Method
2.5. Dissection Procedure
2.6. Outcomes
2.7. Data Analysis
3. Results
3.1. Comparison of Oviposition and Dissection Methods
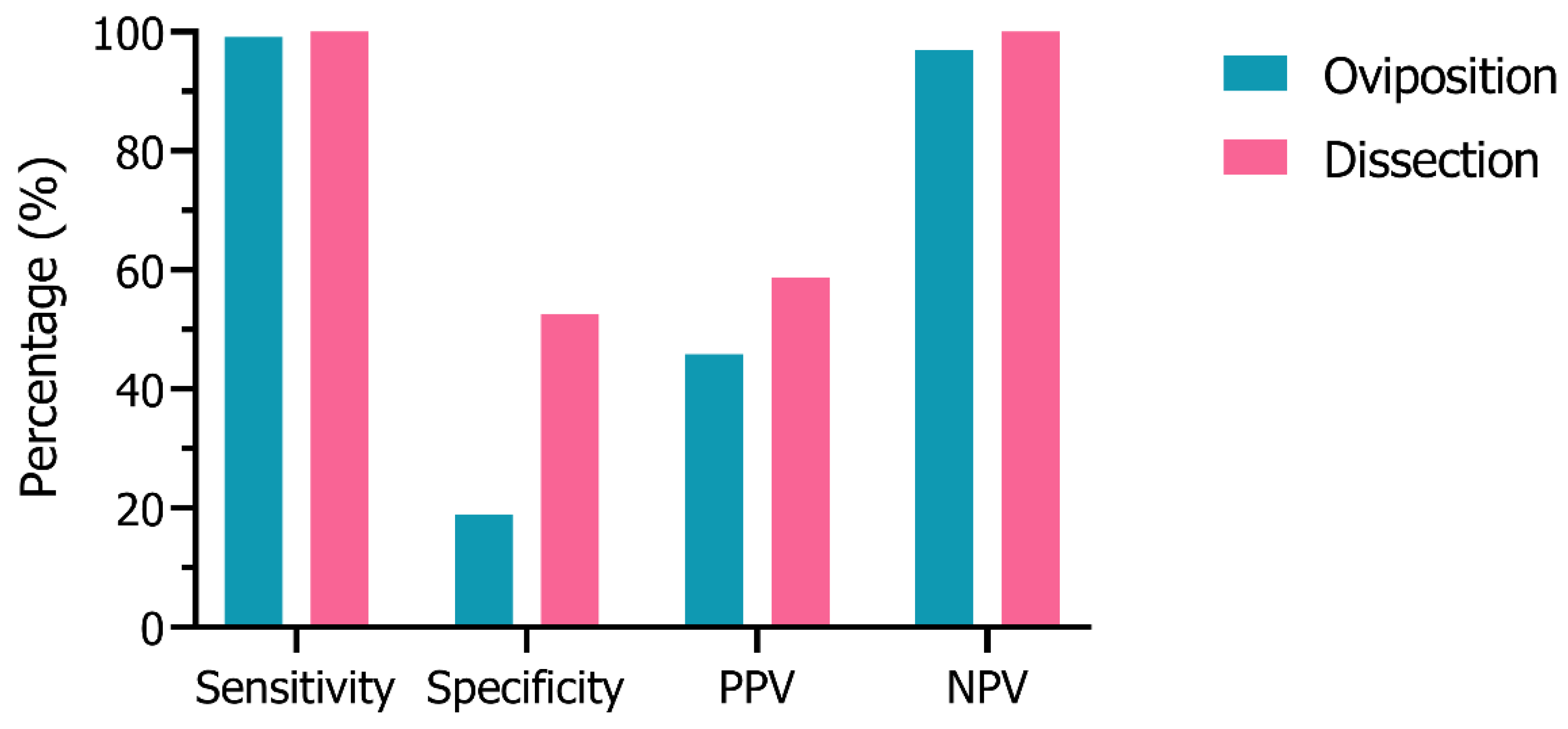
3.2. Sensitivity & Specificity Analysis
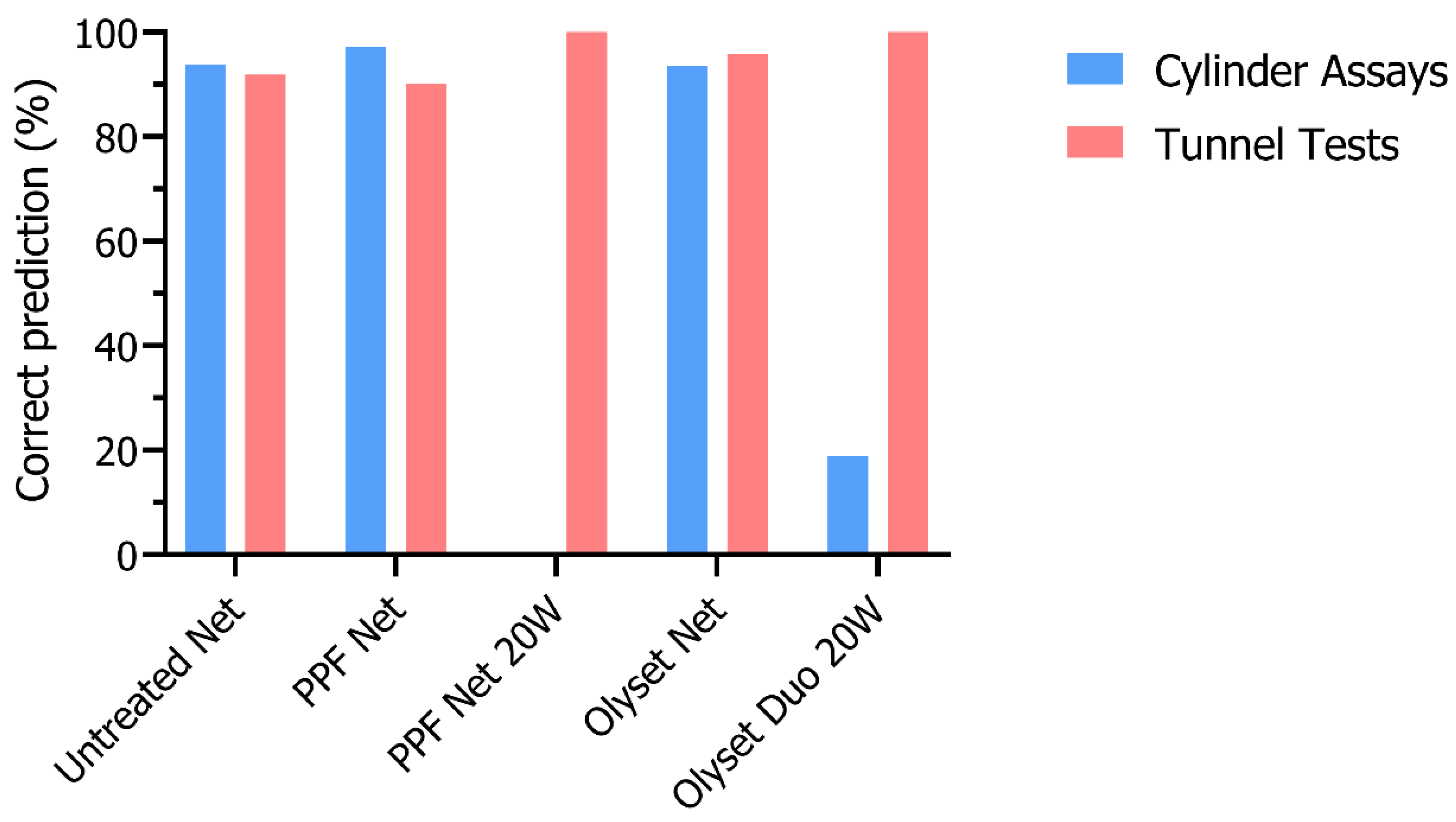
3.3. Dissections after PPF Exposure in Cylinder Assays and Tunnel Tests
3.4. Blinded Dissections
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- WHO. World Malaria Report 2017; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Koama, B.; Namountougou, M.; Sanou, R.; Ndo, S.; Ouattara, A.; Dabiré, R.K.; Malone, D.; Diabaté, A. The sterilizing effect of pyriproxyfen on the malaria vector Anopheles gambiae: Physiological impact on ovaries development. Malar. J. 2015, 14, 101. [Google Scholar] [CrossRef] [PubMed]
- White, M.T.; Lwetoijera, D.; Marshall, J.; Caron-Lormier, G.; Bohan, D.A.; Denholm, I.; Devine, G.J. Negative cross resistance mediated by co-treated bed nets: A potential means of restoring pyrethroid-susceptibility to malaria vectors. PLoS ONE 2014, 9, e95640. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Standard Operating Procedure for Evaluating the Sterilizing Properties of Pyriproxyfen in Adult Female Mosquitoes in WHO Bottle Bioassays; World Health Organization: Geneva, Switzerland, 2022. [Google Scholar]
- WHO. Guidelines for Laboratory and Field-Testing of Long-Lasting Insecticidal Nets, 2013th ed; World Health Organization Press: Geneva, Switzerland, 2013. [Google Scholar]
- Jaffer, A.; Protopopoff, N.; Mosha, F.W.; Malone, D.; Rowland, M.W.; Oxborough, R.M. Evaluating the sterilizing effect of pyriproxyfen treated mosquito nets against Anopheles gambiae at different blood-feeding intervals. Acta Trop. 2015, 150, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Ngufor, C.; N’Guessan, R.; Fagbohoun, J.; Odjo, A.; Malone, D.; Akogbeto, M.; Rowland, M. Olyset Duo® (a pyriproxyfen and permethrin mixture net): An experimental hut trial against pyre-throid resistant Anopheles gambiae and Culex quinquefasciatus in southern Benin. PLoS ONE 2014, 9, e93603. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, K.; Nakada, K.; Ishiwatari, T.; Miyaguchi, J.I.; Shono, Y.; Lucas, J.R.; Mito, N. Efficacy of Pyriproxyfen-Treated Nets in Sterilizing and Shortening the Longevity of Anopheles gambiae (Diptera: Culicidae). J. Med. Entomol. 2012, 49, 1052–1058. [Google Scholar] [CrossRef] [PubMed]
- Harris, C.; Lwetoijera, D.W.; Dongus, S.; Matowo, N.S.; Lorenz, L.M.; Devine, G.J.; Majambere, S. Sterilising effects of pyriproxyfen on Anopheles arabiensis and its potential use in malaria control. Parasites Vectors 2013, 6, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Grisales, N.; Lees, R.S.; Maas, J.; Morgan, J.C.; Wangrawa, D.W.; Guelbeogo, W.M.; N’fale, S.; Lindsay, S.W.; McCall, P.J.; Ranson, H. Pyriproxyfen-treated bed nets reduce reproductive fitness and longevity of pyrethroid-resistant Anopheles gambiae under laboratory and field conditions. Malar. J. 2021, 20, 273. [Google Scholar] [CrossRef] [PubMed]
- Toé, K.H.; Mechan, F.; Tangena, J.-A.A.; Morris, M.; Solino, J.; Tchicaya, E.F.S.; Traoré, A.; Ismail, H.; Maas, J.; Lissenden, N.; et al. Assessing the impact of the addition of pyriproxyfen on the durability of permethrin-treated bed nets in Burkina Faso: A compound-randomized controlled trial. Malar. J. 2019, 18, 355. [Google Scholar] [CrossRef] [PubMed]
- Koffi, A.A. Efficacy of Olyset ® Duo, a permethrin and pyriproxyfen mixture net against wild pyrethroid-resistant Anopheles gambiae s.s. from Côte d’Ivoire: An experimental hut trial. Parasite 2015, 22, 28. [Google Scholar] [CrossRef] [PubMed]
- Kawada, H.; Dida, G.O.; Ohashi, K.; Kawashima, E.; Sonye, G.; Njenga, S.M.; Mwandawiro, C.; Minakawa, N. A Small-Scale Field Trial of Pyriproxyfen-Impregnated Bed Nets against Pyrethroid-Resistant Anopheles gambiae s.s. in Western Kenya. PLoS ONE 2014, 9, e111195. [Google Scholar] [CrossRef] [PubMed]
- Yunta, C.; Grisales, N.; Nász, S.; Hemmings, K.; Pignatelli, P.; Voice, M.; Ranson, H.; Paine, M.J. Pyriproxyfen is metabolized by P450s associated with pyrethroid resistance in An. gambiae. Insect Biochem. Mol. Biol. 2016, 78, 50–57. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Division of Malaria and other Parasitic Diseases. In Manual on Practical Entomology in Malaria; Part II Methods and Techniques; World Health Organization: Geneva, Switzerland, 1975. [Google Scholar]
- Baldini, F.; Gabrieli, P.; South, A.; Valim, C.; Mancini, F.; Catteruccia, F. The Interaction between a Sexually Trans-ferred Steroid Hormone and a Female Protein Regulates Oogenesis in the Malaria Mosquito Anopheles gambiae. PLoS Biol. 2013, 11, e1001695. [Google Scholar] [CrossRef] [PubMed]
- Gulia-Nuss, M.; Eum, J.-H.; Strand, M.R.; Brown, M.R. Ovary ecdysteroidogenic hormone activates egg maturation in the mosquito, Georgecraigius atropalpus, after adult eclosion or a blood meal. J. Exp. Biol. 2012, 215 Pt 21, 3758–3767. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hernández-Martínez, S.; Unnithan, G.C.; Feyereisen, R.; Noriega, F.G. Activity of the corpora allata of adult female Aedes aegypti: Effects of mating and feeding. Insect Biochem. Mol. Biol. 2003, 33, 1307–1315. [Google Scholar] [CrossRef] [PubMed]
- Detinova, T. On the Influence of Glands of internal Secretion upon the Ripening of the Gonads and the imaginai Diapause in Anopheles maculipennis. Zool. Zhurnal. 1945, 5, 291–298. [Google Scholar]
- Gillies, M.T. The duration of the gonotrophic cycle in Anopheles gambiae and Anopheles funestus, with a note on the efficiency of hand catching. East Afr. Med. J. 1953, 30, 129–135. [Google Scholar]
- Rowland, M. Behaviour and fitness of γHCH/dieldrin resistant and susceptible female Anopheles gambiae and An.stephensi mosquitoes in the absence of insecticide. Med. Vet. Entomol. 1991, 5, 193–206. [Google Scholar] [CrossRef]
- Fernandes, L.; Briegel, H. Reproductive physiology of Anopheles gambiae and Anopheles atroparvus. J. Vector Ecol. 2005, 30, 11–26. [Google Scholar] [PubMed]
- Takken, W.; Klowden, M.J.; Chambers, G.M. Effect of body size on host seeking and blood meal utilization in Anopheles gambiae sensu stricto (Diptera: Culicidae): The disadvantage of being small. J. Med. Èntomol. 1998, 35, 639–645. [Google Scholar] [CrossRef]
- Detinova, T. Age-Grouping Methods in Diptera of Medical Importance with Special Reference to Some Vectors of Malaria; World Health Organization: Geneva, Switzerland, 1962. [Google Scholar]
- Clements, A.N. The Biology of Mosquitoes: Development, Nutrition and Reproduction; Chapman & Hall: London, UK, 1992. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Outcome | Oviposition Group | Dissection Group |
|---|---|---|
| % Fecund females | Proportion of females that laid eggs over the total number of blood-fed females alive at 3 days post-exposure. | Proportion of dissected females with mature eggs over the total number of blood-fed females dissected at 3 days post-exposure. |
| % Induced sterility (proxy for oviposition inhibition) | Reduction in the proportion of egg-laying females caused by PPF exposure. | Reduction in the proportion of dissected females with mature eggs caused by PPF exposure. |
| Fecundity | Mean number of eggs laid per female, calculated as the total number of eggs laid divided by the total number of blood-fed females. | Mean number of mature eggs per female, calculated as the total number of mature eggs divided by the total number of blood-fed females. |
| % Reduction in fecundity | Reduction in the proportion of eggs laid per female caused by PPF exposure. | Reduction in the proportion of mature eggs dissected per female caused by PPF exposure. |
| Fertility | Proportion of laid eggs that hatched into larvae. | - |
| Reproductive rate | Mean number of larvae that hatched per blood-fed female alive at 3 days post-exposure. | - |
| % Reduction in reproductive rate | Reduction in the proportion of larvae that hatched per female caused by PPF exposure. | - |
| PPF Group | Untreated Group | Calculation | |
|---|---|---|---|
| Sterile | A | B | PPV = A/(A + B) |
| Fecund | C | D | NPV = D/(C + D) |
| Calculation | Sensitivity = A/(A + C) | Specificity = D/(B + D) |
| Oviposition | Dissection | |||
|---|---|---|---|---|
| Untreated | PPF | Untreated | PPF | |
| N Survivors at 3 days post-exposure | 164 | 114 | 80 | 54 |
| N Survivors at 6 days post-exposure | 99 | 41 | - | - |
| N Dissected females with eggs | - | - | 59 | 36 |
| N Fecund females | 31 | 1 | 42 | 0 |
| % Fecund females (95% CI) | 17.0 (12.2–23.3) | 0.7 (0.1–4.9) | 71.2 (58.0–81.5) | 0.0 |
| % Induced sterility | - | 96.8 | - | 100.0 |
| N Underdeveloped (stages II–IV) eggs | 0 (not laid) | 0 (not laid) | 918 | 3510 |
| N Developed (stage V) eggs | 1778 | 75 | 2393 | 0.0 |
| N Mean eggs per female | 10.8 | 1.5 | 41.4 | 65.0 |
| Fecundity: N Mean mature eggs per female | 10.8 | 1.5 | 29.9 | 0.0 |
| % Reduction in fecundity | - | 86.1 | - | 100.0 |
| Cylinder Assay | Tunnel Test | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Untreated | PPF | Olyset® Net | Olyset® Duo 20W | Untreated | PPF | PPF 20W | Olyset® Net | Olyset® Duo 20W | |
| N Exposed | 174 | 189 | 181 | 129 | 205 | 196 | 50 | 164 | 115 |
| N Blood-fed (tunnel tests) | - | - | - | - | 124 | 132 | 50 | 38 | 34 |
| % Mortality 24 h (95% CI) | 5.2 (2.7–9.7) | 11.6 (7.8–17.1) | 9.4 (5.9–14.6) | 9.3 (5.3–15.7) | 13.7 (8.7–21.0) | 9.4 (5.4–15.8) | 18.2 (9.3–32.6) | 8.6 (2.7–23.8) | 8.8 (2.8–24.4) |
| N Females dissected | 142 | 113 | 145 | 93 | 90 | 98 | 28 | 25 | 30 |
| % Females with eggs, all stages (95% CI) | 81.0 (73.6–86.7) | 93.8 (87.5–97.0) | 89.0 (82.7–93.1) | 90.3 (82.4–94.9) | 83.3 (74.1–89.7) | 95.9 (89.5–98.5) | 96.4 (77.8–99.5) | 96.0 (75.5–99.5) | 86.7 (68.9–95.0) |
| N Females with mature eggs | 105 | 2 | 116 | 60 | 66 | 7 | 0 | 23 | 9 |
| % Females with mature eggs (95% CI) | 89.7 (82.7–94.1) | 1.9 * (0.5–7.3) | 89.9 (83.4–94.1) | 71.4 * (60.8–80.1) | 88.0 (78.4–93.7) | 7.4 * (3.6–14.9) | 0.0 * | 95.8 (74.7–99.4) | 34.6 * (18.8–54.8) |
| % Induced sterility | - | 97.9 | 0.0 | 20.4 | - | 91.6 | 100.0 | 0.0 | 60.7 |
| N Eggs, all stages (95% CI) | 8422 (7658–9186) | 8399 (7965–8832) | 9705 (8897–10,512) | 6598 (5848–7347) | 5926 (5347–6505) | 7582 (7094–8070) | 1974 (1764–2183) | 2176 (1928–2424) | 1590 (1310–1869) |
| N Mature eggs dissected (n, 95% CI) | 8127 (7391–8863) | 68 (0–159) | 9285 (8511–10,059) | 5202 (4542–5862) | 5392 (4828–5956) | 699 (481–916) | 0 | 2065 (1818–2311) | 648 (421–875) |
| N Mean eggs dissected, all stages (SD) | 75.2 (36.7) | 81.5 (21.7) | 77.6 (36.8) | 78.5 (41.6) | 79.0 (34.0) | 80.7 (25.5) | 73.1 (20.5) | 90.7 (25.7) | 61.1 (27.8) |
| N Mean mature eggs dissected (SD) | 77.4 (36.5) | 34.0 * (32.5) | 80.0 (36.5) | 86.7 * (43.3) | 81.7 (35.0) | 99.8 * (41.5) | 0.0 | 89.8 * (25.9) | 72.0 * (38.2) |
| Fecundity: N Mean mature eggs per female | 57.2 | 0.6 | 64.0 | 55.9 | 59.9 | 7.1 | 0.0 | 82.6 | 21.6 |
| % Reduction in fecundity | - | 99.0 | 0.0 | 2.3 | - | 88.1 | 100.0 | 0.0 | 63.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soto, A.; Rowland, M.; Messenger, L.A.; Kirby, M.; Mosha, F.W.; Manjurano, A.; Protopopoff, N. Ovary Dissection Is a Sensitive Measure of Sterility in Anopheles gambiae Exposed to the Insect Growth Regulator Pyriproxyfen. Insects 2023, 14, 552. https://doi.org/10.3390/insects14060552
Soto A, Rowland M, Messenger LA, Kirby M, Mosha FW, Manjurano A, Protopopoff N. Ovary Dissection Is a Sensitive Measure of Sterility in Anopheles gambiae Exposed to the Insect Growth Regulator Pyriproxyfen. Insects. 2023; 14(6):552. https://doi.org/10.3390/insects14060552
Chicago/Turabian StyleSoto, Alina, Mark Rowland, Louisa A. Messenger, Mathew Kirby, Franklin W. Mosha, Alphaxard Manjurano, and Natacha Protopopoff. 2023. "Ovary Dissection Is a Sensitive Measure of Sterility in Anopheles gambiae Exposed to the Insect Growth Regulator Pyriproxyfen" Insects 14, no. 6: 552. https://doi.org/10.3390/insects14060552
APA StyleSoto, A., Rowland, M., Messenger, L. A., Kirby, M., Mosha, F. W., Manjurano, A., & Protopopoff, N. (2023). Ovary Dissection Is a Sensitive Measure of Sterility in Anopheles gambiae Exposed to the Insect Growth Regulator Pyriproxyfen. Insects, 14(6), 552. https://doi.org/10.3390/insects14060552