
The Distribution Pattern and Species Richness of Scorpionflies (Mecoptera: Panorpidae)

 and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Resources
2.2. Identifying Bioclimatic Variables
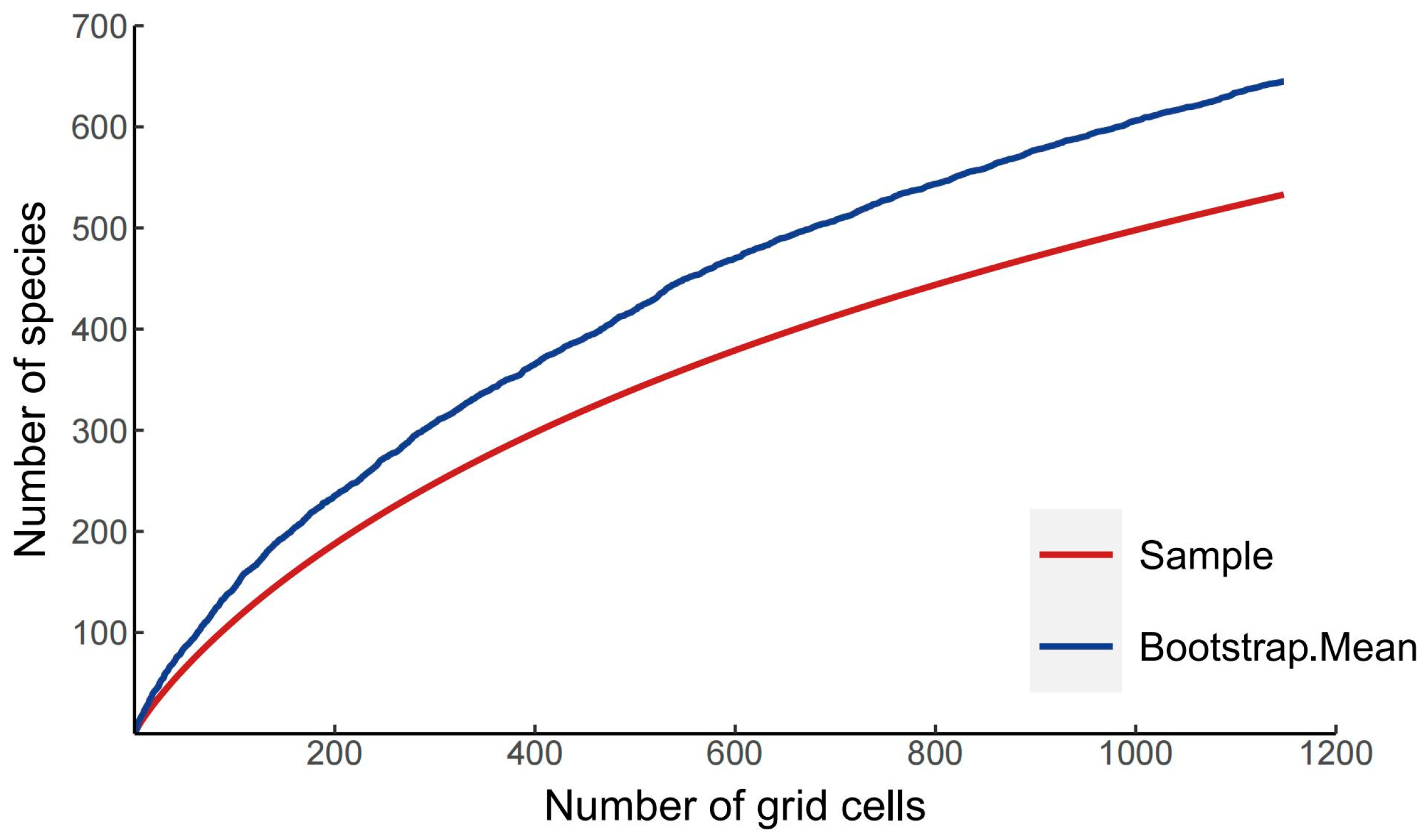
2.3. Analysis of Species Richness
2.4. Model Evaluation Index
2.5. Model Setting
2.6. Predicting the Appropriate Panorpidae Habitat under Climate Change
3. Results
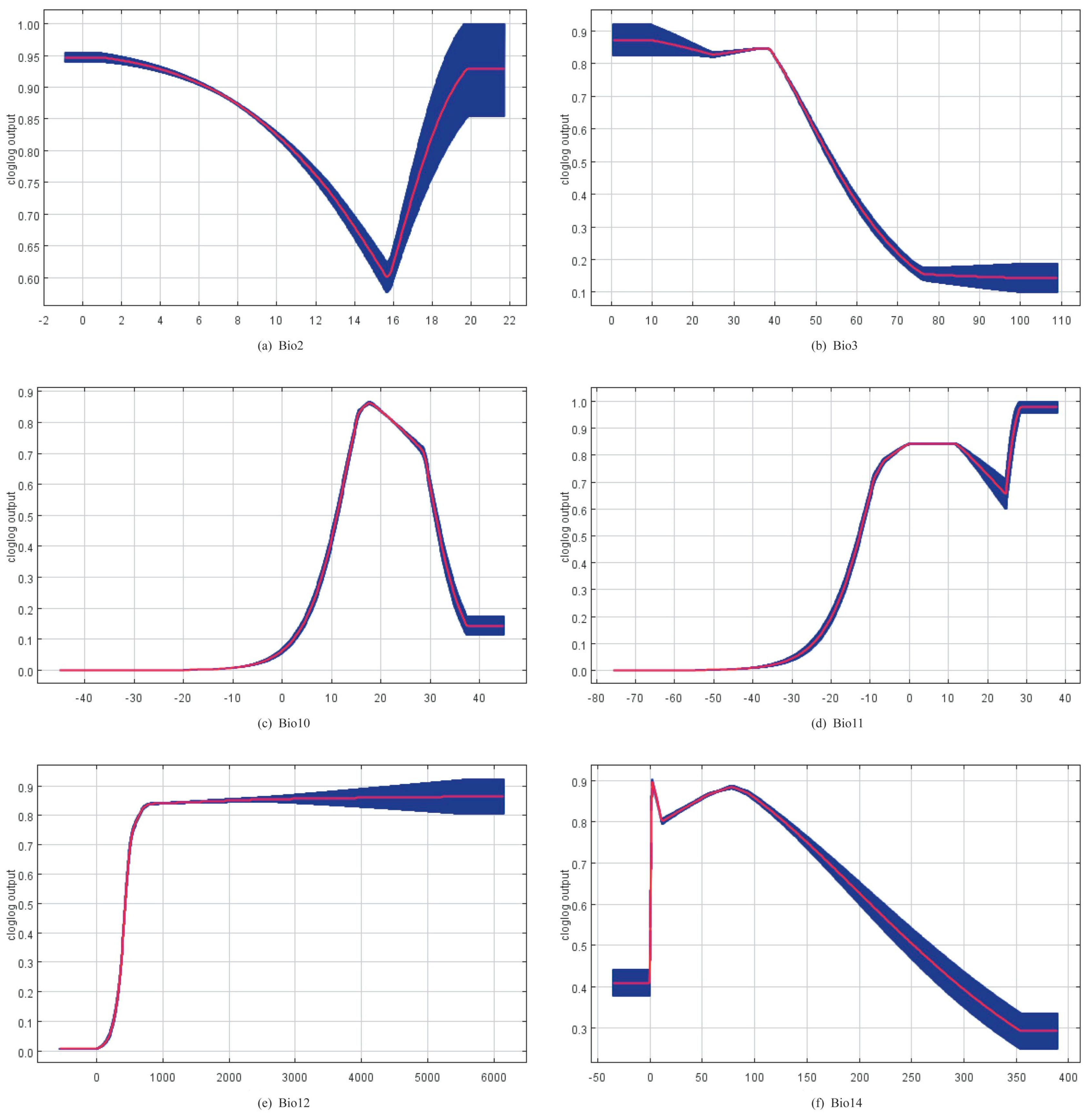
3.1. Relationships between Species Richness and Environmental Variables
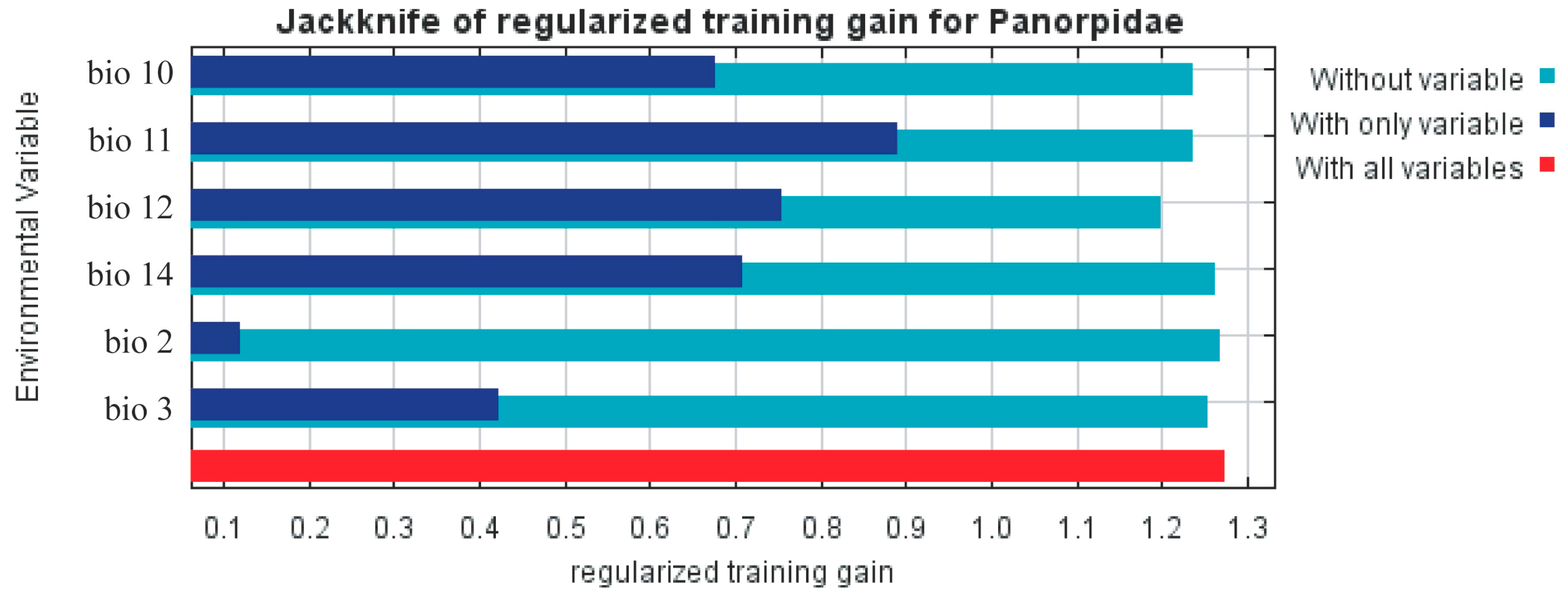
3.2. Evaluation of the MaxEnt Model and the Impact of Environmental Variables
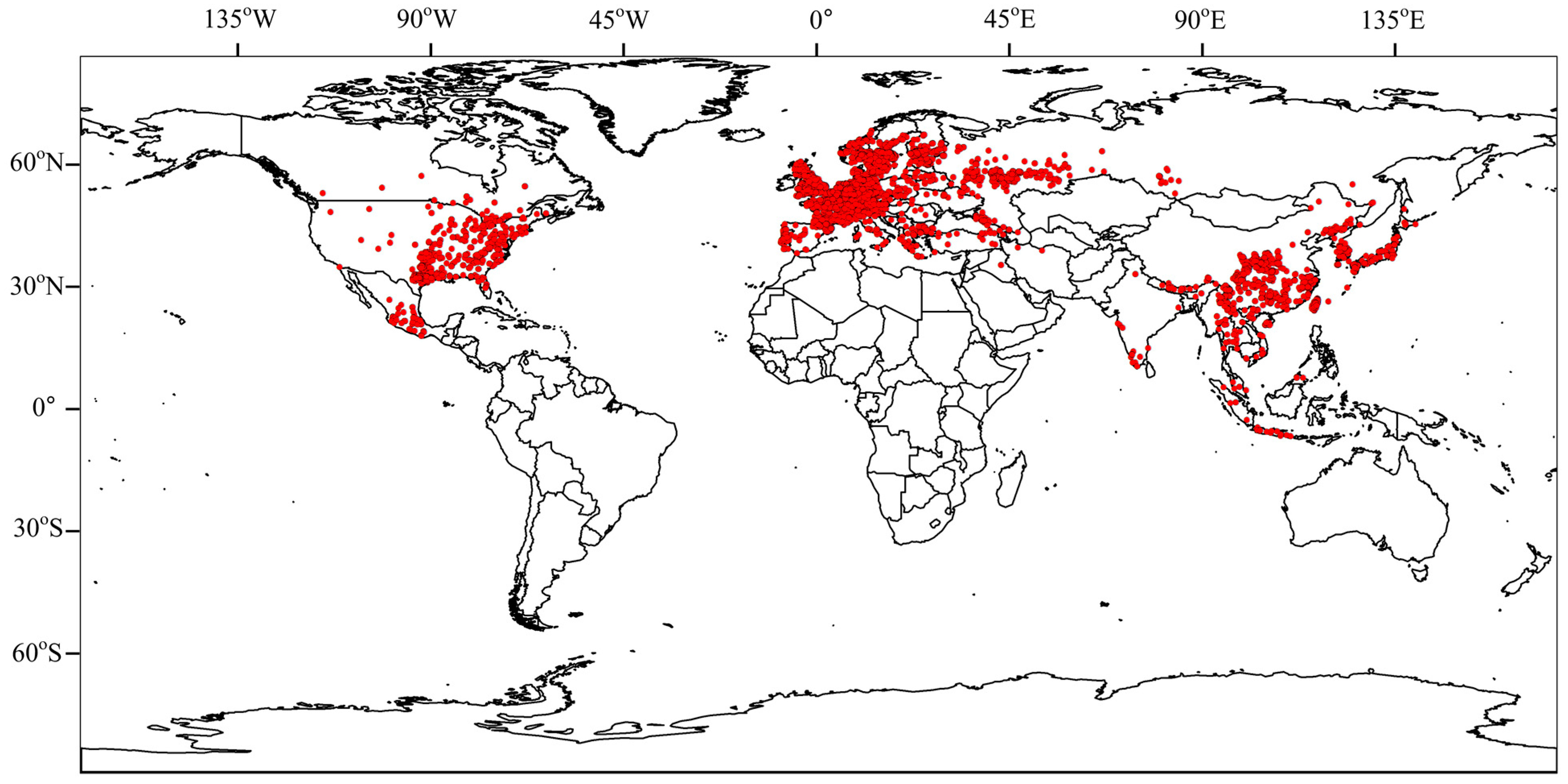
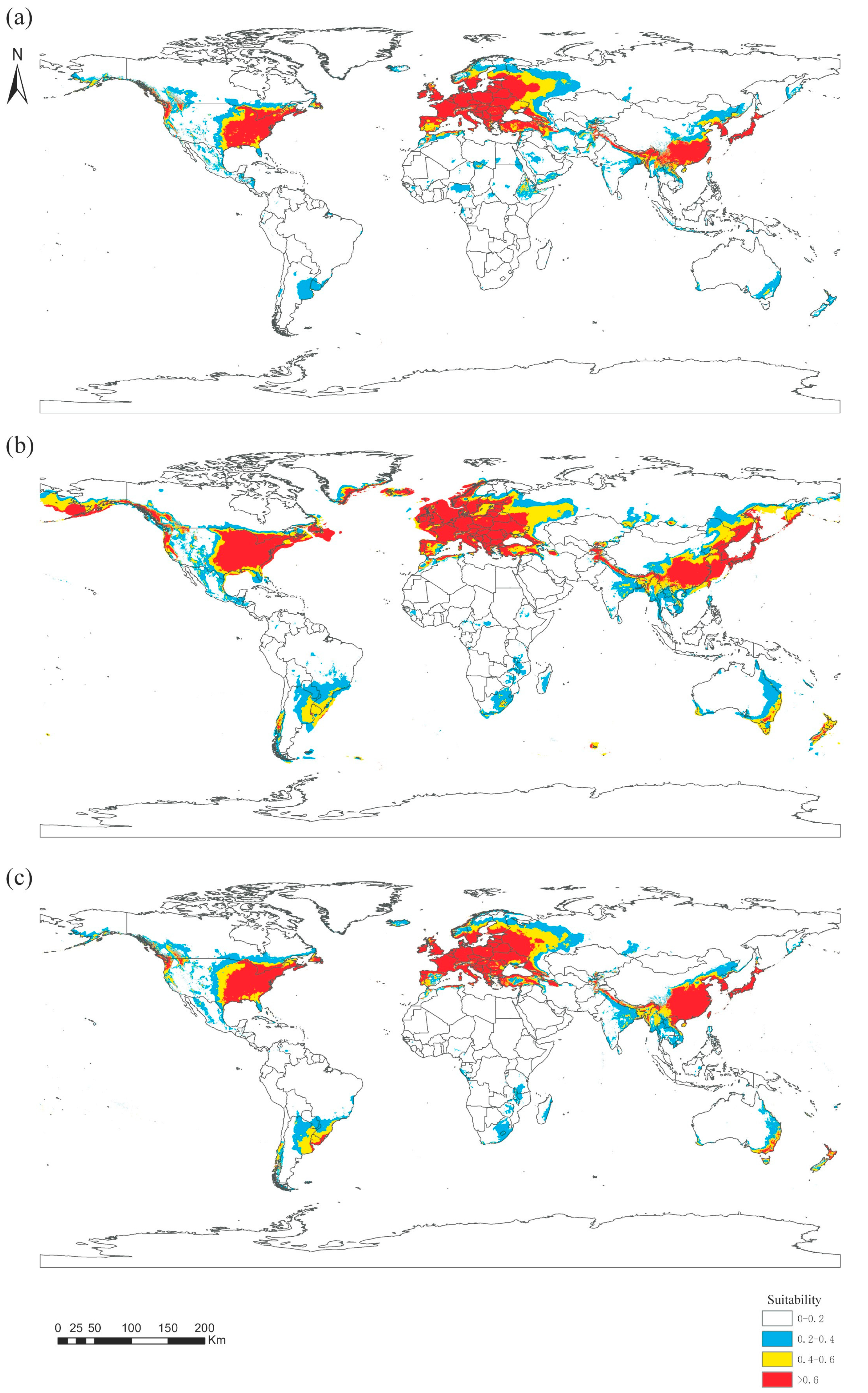
3.3. Global Distribution of Panorpidae
3.4. Predicted Habitat Suitability for Three Historical Eras
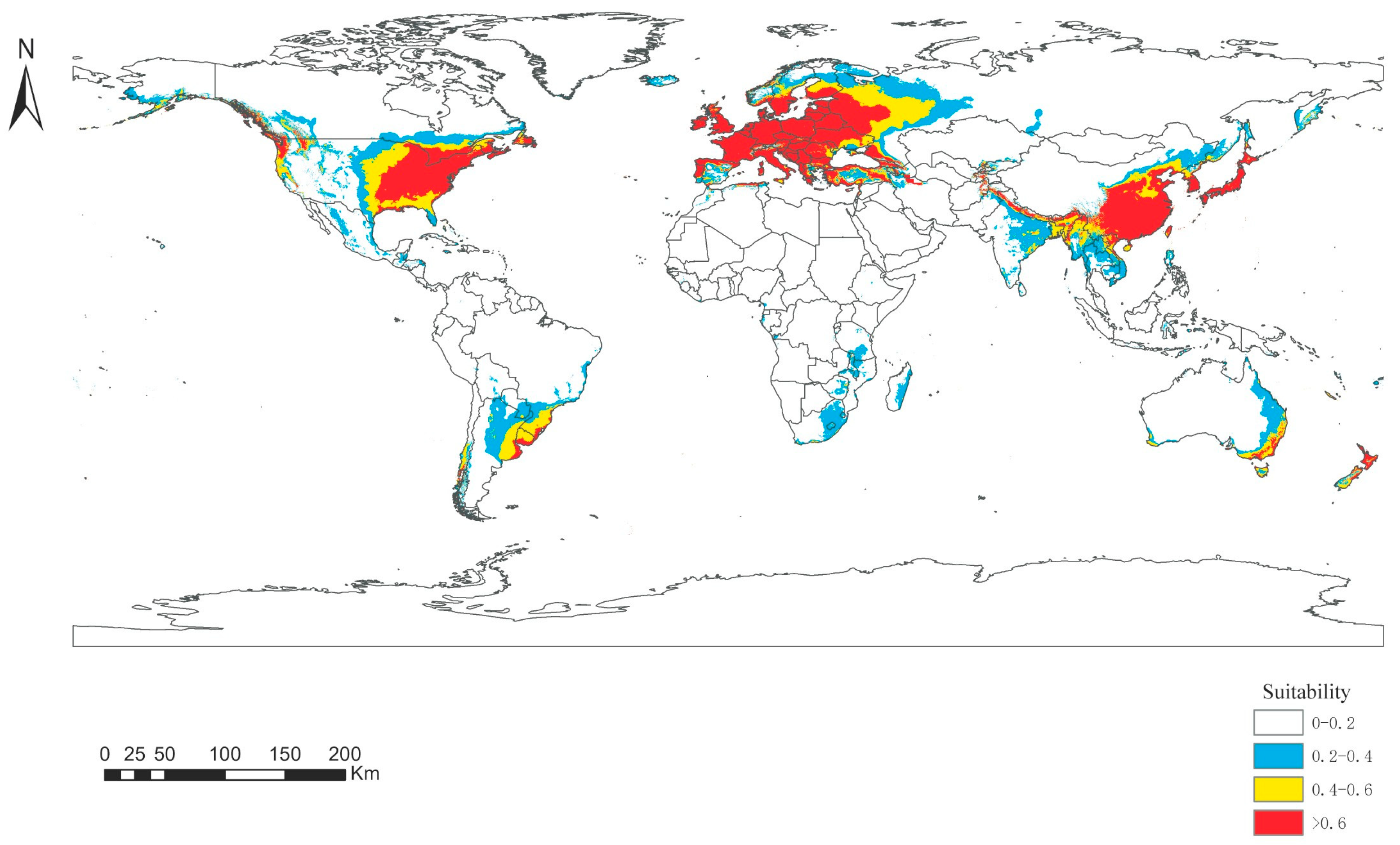
3.5. Distribution Trends of Panorpidae during Future Global Warming
4. Discussion
4.1. Patterns of Species Richness and Influencing Factors
4.2. Relationship between Environmental Factors and the Suitability of Panorpidae Habitat
4.3. Simulation of Habitat Suitability for Three Periods of History
4.4. Changes in Panorpidae Distribution in the Future
4.5. Implications for Biological Conservation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Kohler, T.; Maselli, D. Mountains and Climate Change. From Understanding to Action; Geographica Bernensia: Bern, Switzerland, 2009. [Google Scholar]
- Knowles, L.L.; Massatti, R. Distributional shifts—Not geographic isolation—As a probable driver of montane species divergence. Ecography 2017, 40, 1475–1485. [Google Scholar] [CrossRef]
- Hu, G.L.; Hua, Y.; Hebert, P.D.; Hua, B.Z. Evolutionary history of the scorpionfly Dicerapanorpa magna (Mecoptera, Panorpidae). Zool. Scr. 2019, 48, 93–105. [Google Scholar] [CrossRef]
- Liu, T.; Liu, H.; Tong, J.; Yang, Y. Habitat suitability of neotenic net-winged beetles (Coleoptera: Lycidae) in China using combined ecological models, with implications for biological conservation. Divers. Distrib. 2022, 28, 2806–2823. [Google Scholar] [CrossRef]
- Peterson, A.T.; Shaw, J. Lutzomyia vectors for cutaneous leishmaniasis in Southern Brazil: Ecological niche models, predicted geographic distributions, and climate change effects. Int. J. Parasitol. 2003, 33, 919–931. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, H.; He, D.; Wang, X.; Bai, M. The modeling and forecasting of carabid beetle distribution in northwestern China. Insects 2021, 12, 168. [Google Scholar] [CrossRef]
- Miao, Y.; Wang, J.S.; Hua, B.Z. Molecular phylogeny of the scorpionflies Panorpidae (Insecta: Mecoptera) and chromosomal evolution. Cladistics 2019, 35, 385–400. [Google Scholar] [CrossRef]
- Tong, X.; Hua, B.Z. Nuptial feeding and genital coupling of Neopanorpa scorpionflies (Insecta: Mecoptera: Panorpidae) with notal organs of various lengths. Contrib. Zool. 2019, 88, 498–512. [Google Scholar] [CrossRef]
- Gao, K.; Hua, Y.; Xing, L.X.; Hua, B.Z. Speciation of the cold-adapted scorpionfly Cerapanorpa brevicornis (Mecoptera: Panorpidae) via interglacial refugia. Insect Conserv. Divers. 2022, 15, 114–127. [Google Scholar] [CrossRef]
- Dvořák, L.; Japoshvili, G. The scorpionflies (Mecoptera) from Lagodekhi Reserve, with some distributional and seasonal activity data. Zootaxa 2018, 4407, 137–140. [Google Scholar] [CrossRef]
- Wang, P.Y.; Hua, B.Z. Elevational diversity pattern and allochronic divergence of scorpionflies in the Qinling Mountains. Ecol. Indic. 2022, 134, 108500. [Google Scholar] [CrossRef]
- Harrison, S.; Cornell, H. Toward a better understanding of the regional causes of local community richness. Ecol. Lett. 2008, 11, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Ryder, O.A. Species conservation and systematics: The dilemma of subspecies. Trends Ecol. Evol. 1986, 1, 9–10. [Google Scholar] [CrossRef]
- Liu, G.; Zhang, X.; Cui, Z.; Wu, Y.; Ju, Y. A review of glacial sequences of the Kunlun Pass, northern Tibetan Plateau. Quat. Int. 2006, 154, 63–72. [Google Scholar] [CrossRef]
- Lattin, G.d. Grundriss der Zoogeographie; Gustav Fischer Verlag: Jena, Germany, 1967. [Google Scholar]
- Comes, H.P.; Kadereit, J.W. Spatial and temporal patterns in the evolution of the flora of the European Alpine System. Taxon 2003, 52, 451–462. [Google Scholar] [CrossRef]
- Bannikova, A.; Lebedev, V. Genetic heterogenity of the Caucasian shrew Sorex satununi (Mammalia, Lipotyphla, Soricidae) inferred from the mtDNA markers as a potential consequence of ancient hybridization. Mol. Biol. 2010, 44, 658. [Google Scholar] [CrossRef]
- Bell, J.R.; Botham, M.S.; Henrys, P.A.; Leech, D.I.; Pearce-Higgins, J.W.; Shortall, C.R.; Brereton, T.M.; Pickup, J.; Thackeray, S.J. Spatial and habitat variation in aphid, butterfly, moth and bird phenologies over the last half century. Glob. Chang. Biol. 2019, 25, 1982–1994. [Google Scholar] [CrossRef]
- Fourcade, Y.; WallisDeVries, M.F.; Kuussaari, M.; van Swaay, C.A.; Heliölä, J.; Öckinger, E. Habitat amount and distribution modify community dynamics under climate change. Ecol. Lett. 2021, 24, 950–957. [Google Scholar] [CrossRef]
- Bertolino, S.; Sciandra, C.; Bosso, L.; Russo, D.; Lurz, P.W.; Di Febbraro, M. Spatially explicit models as tools for implementing effective management strategies for invasive alien mammals. Mamm. Rev. 2020, 50, 187–199. [Google Scholar] [CrossRef]
- Fung, T.; Verma, S.; Chisholm, R.A. Probability distributions of extinction times, species richness, and immigration and extinction rates in neutral ecological models. J. Theor. Biol. 2020, 485, 110051. [Google Scholar] [CrossRef]
- Hill, J.K.; Thomas, C.D.; Huntley, B. Climate and habitat availability determine 20th century changes in a butterfly’s range margin. Proc. R. Soc. Lond. B Biol. Sci. 1999, 266, 1197–1206. [Google Scholar] [CrossRef]
- Wagner, D.L. Insect declines in the Anthropocene. Annu. Rev. Entomol. 2020, 65, 457–480. [Google Scholar] [CrossRef] [PubMed]
- Soroye, P.; Newbold, T.; Kerr, J. Climate change contributes to widespread declines among bumble bees across continents. Science 2020, 367, 685–688. [Google Scholar] [CrossRef]
- Sillero, N. What does ecological modelling model? A proposed classification of ecological niche models based on their underlying methods. Ecol. Model. 2011, 222, 1343–1346. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Kumar, P. Assessment of impact of climate change on Rhododendrons in Sikkim Himalayas using Maxent modelling: Limitations and challenges. Biodivers Conserv. 2012, 21, 1251–1266. [Google Scholar] [CrossRef]
- Roth, N.; Zoder, S.; Zaman, A.A.; Thorn, S.; Schmidl, J. Long-term monitoring reveals decreasing water beetle diversity, loss of specialists and community shifts over the past 28 years. Insect Conserv. Divers. 2020, 13, 140–150. [Google Scholar] [CrossRef]
- Dvořák, L.; Georgiev, D. New and interesting records of scorpion flies of the genus Panorpa (Mecoptera: Panorpidae) from Bulgaria with a country checklist. Ecol. Montenegrina 2017, 15, 22–25. [Google Scholar] [CrossRef]
- Dvořák, L.; Haenni, J.P.; Máca, J.; Mariychuk, R.; Oboňa, J. Some insects (Dermaptera, Diptera, Mecoptera) from beer traps in Uzhhorod City (Ukraine). Acta Univ. Presov. 2017, 9, 11. [Google Scholar]
- Dvořák, L. New records of the scorpion fly Panorpa rufostigma Westwood, 1846 (Mecoptera: Panorpidae) from Greece. Parnass. Arch. 2018, 6, 7–9. [Google Scholar]
- Makarkin, V.N.; Shchurov, V.I. Neuropterida and Mecoptera from the North-Western Caucasus. Cauc. Entomol. Bull. 2019, 15, 299–316. [Google Scholar] [CrossRef]
- Shchurov, V.I.; Makarkin, V.N. Neuroptera, Raphidioptera and Mecoptera from the North Caucasus and Western Transcaucasia. Cauc. Entomol. Bull. 2022, 18, 103–129. [Google Scholar] [CrossRef]
- Dvořák, L.; Ruchin, A.B.; Egorov, L.V.; Aleksanov, V.V.; Alekseev, S.K.; Shulaev, N.V.; Zakharova, E.Y. Distribution of species from the genus Panorpa (Mecoptera, Panorpidae) in European Russia except the Caucasus. Nat. Conserv. Res. 2023, 8, 24–33. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Ma, B.; Sun, J. Predicting the distribution of Stipa purpurea across the Tibetan Plateau via the MaxEnt model. BMC Ecol. 2018, 18, 10. [Google Scholar] [CrossRef]
- Moss, R.H.; Babiker, M.; Brinkman, S.; Calvo, E.; Carter, T.; Edmonds, J.A.; Elgizouli, I.; Emori, S.; Lin, E.; Hibbard, K. Towards New Scenarios for Analysis of Emissions, Climate Change, Impacts, and Response Strategies; Pacific Northwest National Lab (PNNL): Richland, WA, USA, 2008. [Google Scholar]
- Weyant, J.; Azar, C.; Kainuma, M.; Jiang, K.; Nakicenovic, N.; Shukla, P.; Rovere, E.L.; Yohe, G. Report of 2.6 versus 2.9 Watts/m2 RCPP Evaluation Panel; Integrated Assessment Modeling Consortium: College Park, MD, USA, 2009. [Google Scholar]
- Yi, Y.J.; Cheng, X.; Yang, Z.F.; Zhang, S.H. Maxent modeling for predicting the potential distribution of endangered medicinal plant (H. riparia Lour) in Yunnan, China. Ecol. Eng. 2016, 92, 260–269. [Google Scholar] [CrossRef]
- Du, J.; Chen, Z. Method of path analysis with SPSS linear regression. Bull. Biol. 2010, 45, 4–6. [Google Scholar]
- Sun, Y.; Shi, M.; Peng, H.; Zhu, P.; Liu, S.; Wu, S.; He, C.; Chen, F. Forest lighting fire forecasting for Daxing’anling Mountains based on MAXENT model. J. Appl. Ecol. 2014, 25, 1100–1106. [Google Scholar]
- Hijmans, R.; Guarino, G.; Macathur, P. DIVA-GIS Version 7.5 Manual. 2012. Available online: https://www.diva-gis.org/docs/DIVA-GIS_manual_7.pdf (accessed on 5 May 2022).
- Colwell, R. EstimateS, version 9.1. Statistical Estimation of Species Richness and Shared Species from Samples. University of Connecticut: Storrs, CT, USA, 2013.
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef]
- Yang, X.Q.; Kushwaha, S.P.S.; Saran, S.; Xu, J.C.; Roy, P.S. Maxent modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda L. in Lesser Himalayan foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Ray, D.; Behera, M.D.; Jacob, J. Predicting the distribution of rubber trees (Hevea brasiliensis) through ecological niche modelling with climate, soil, topography and socioeconomic factors. Ecol. Res. 2016, 31, 75–91. [Google Scholar] [CrossRef]
- Morales, N.S.; Fernández, I.C.; Baca-González, V. MaxEnt’s parameter configuration and small samples: Are we paying attention to recommendations? A systematic review. PeerJ 2017, 5, e3093. [Google Scholar] [CrossRef] [PubMed]
- Galante, P.J.; Alade, B.; Muscarella, R.; Jansa, S.A.; Goodman, S.M.; Anderson, R.P. The challenge of modeling niches and distributions for data-poor species: A comprehensive approach to model complexity. Ecography 2018, 41, 726–736. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Li, G.; Du, S.; Guo, K. Evaluation of limiting climatic factors and simulation of a climatically suitable habitat for Chinese sea buckthorn. PLoS ONE 2015, 10, e0131659. [Google Scholar]
- Sosa, V.; Loera, I. Influence of current climate, historical climate stability and topography on species richness and endemism in Mesoamerican geophyte plants. PeerJ 2017, 5, e3932. [Google Scholar] [CrossRef]
- Graham, C.H.; Elith, J.; Hijmans, R.J.; Guisan, A.; Townsend Peterson, A.; Loiselle, B.A.; Group, N.P.S.D.W. The influence of spatial errors in species occurrence data used in distribution models. J. Appl. Ecol. 2008, 45, 239–247. [Google Scholar] [CrossRef]
- Hu, G.L.; Gao, K.; Wang, J.S.; Hebert, P.D.; Hua, B.Z. Molecular phylogeny and species delimitation of the genus Dicerapanorpa (Mecoptera: Panorpidae). Zool. J. Linn. Soc. 2019, 187, 1173–1195. [Google Scholar] [CrossRef]
- Byers, G.W. Geographic affinities of the North American Mecoptera. Mem. Ent. Soc. Can. 1988, 120, 25–30. [Google Scholar] [CrossRef]
- Wang, J.S.; Hua, B.Z. Morphological phylogeny of Panorpidae (Mecoptera: Panorpoidea). Syst. Entomol. 2021, 46, 526–557. [Google Scholar] [CrossRef]
- Axelrod, D.I. Rise of the grassland biome, central North America. Bot. Rev. 1985, 51, 163–201. [Google Scholar] [CrossRef]
- Willmann, R. The phylogenetic system of the Mecoptera. Syst. Entomol. 1987, 12, 519–524. [Google Scholar] [CrossRef]
- Fløjgaard, C.; Normand, S.; Skov, F.; Svenning, J.C. Deconstructing the mammal species richness pattern in Europe–towards an understanding of the relative importance of climate, biogeographic history, habitat heterogeneity and humans. Glob. Ecol. Biogeogr. 2011, 20, 218–230. [Google Scholar] [CrossRef]
- Penny, N.D.; Byers, G.W. A check-list of the Mecoptera of the world. Acta Amazon. 1979, 9, 365–388. [Google Scholar] [CrossRef]
- Li, N.; Jiang, L.; Wang, J.S.; Hua, B.Z. Integrative taxonomy of the seasonally polyphenic scorpionfly Panorpa liui Hua, 1997 (Mecoptera: Panorpidae). Org. Divers. Evol. 2021, 21, 533–545. [Google Scholar] [CrossRef]
- Chau, H.C.S.; Byers, G.W. The Mecoptera of Indonesia: Genus Neopanorpa [New taxa]. Univ. Kans. Sci. Bull. 1978. [Google Scholar]
- Cai, L.; Li, J.; Hua, B. Life history of the scorpionfly Panorpa qinlingensis in Qinling Mountains of Shaanxi. Chin. Bull. Entomol. 2006, 43, 681–683. [Google Scholar]
- Byers, G.W.; Thornhill, R. Biology of the Mecoptera. Annu. Rev. Entomol. 1983, 28, 203–228. [Google Scholar] [CrossRef]
- Malohlava, V.; Bocak, L. Evidence of extreme habitat stability in a Southeast Asian biodiversity hotspot based on the evolutionary analysis of neotenic net-winged beetles. Mol. Ecol. 2010, 19, 4800–4811. [Google Scholar] [CrossRef] [PubMed]
- Masek, M.; Palata, V.; Bray, T.C.; Bocak, L. Molecular phylogeny reveals high diversity, geographic structure and limited ranges in neotenic net-winged beetles Platerodrilus (Coleoptera: Lycidae). PLoS ONE 2015, 10, e0123855. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Tierno de Figueroa, J.M.; Lek, S.; Park, Y.S. Continental drift and climate change drive instability in insect assemblages. Sci. Rep. 2015, 5, 11343. [Google Scholar] [CrossRef] [PubMed]
- Byers, G.W. The life history of Panorpa nuptialis (Mecoptera: Panorpidae). Ann. Entomol. Soc. Am. 1963, 56, 142–149. [Google Scholar] [CrossRef]
- Lamb, R.J.; Gerber, G.H. Effects of temperature on the development, growth, and survival of larvae and pupae of a north-temperate chrysomelid beetle. Oecologia 1985, 67, 8–18. [Google Scholar] [CrossRef]
- Koster, R.D.; Dirmeyer, P.A.; Guo, Z.; Bonan, G.; Chan, E.; Cox, P.; Gordon, C.; Kanae, S.; Kowalczyk, E.; Lawrence, D. Regions of strong coupling between soil moisture and precipitation. Science 2004, 305, 1138–1140. [Google Scholar] [CrossRef]
- Ernst, C.M.; Buddle, C.M. Drivers and patterns of ground-dwelling beetle biodiversity across Northern Canada. PLoS ONE 2015, 10, e0122163. [Google Scholar] [CrossRef]
- Knapp, A.K.; Fay, P.A.; Blair, J.M.; Collins, S.L.; Smith, M.D.; Carlisle, J.D.; Harper, C.W.; Danner, B.T.; Lett, M.S.; McCarron, J.K. Rainfall variability, carbon cycling, and plant species diversity in a mesic grassland. Science 2002, 298, 2202–2205. [Google Scholar] [CrossRef]
- Zavaleta, E.S.; Shaw, M.R.; Chiariello, N.R.; Mooney, H.A.; Field, C.B. Additive effects of simulated climate changes, elevated CO2, and nitrogen deposition on grassland diversity. Proc. Natl. Acad. Sci. USA 2003, 100, 7650–7654. [Google Scholar] [CrossRef]
- Palmer, C. Diversity of feeding strategies in adult Mecoptera. Terr. Arthropod Rev. 2010, 3, 111–128. [Google Scholar] [CrossRef]
- Flügel, H.J.; Malec, F. Aktuelle Schnabelfliegen-Nachweise (Mecoptera) aus Nord-hessen und ihre Blütenbesuche. Lebbimuk 2013, 10, 3–18. [Google Scholar]
- Bicha, W. Biodiversity of Mecoptera. In Insect Biodiversity: Science and Society, II; Foottit, R.G., Adler, P.H., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2018. [Google Scholar]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.; De Siqueira, M.F.; Grainger, A.; Hannah, L. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Trisos, C.H.; Merow, C.; Pigot, A.L. The projected timing of abrupt ecological disruption from climate change. Nature 2020, 580, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Wolda, H. Altitude, habitat and tropical insect diversity. Biol. J. Linn. Soc. 1987, 30, 313–323. [Google Scholar] [CrossRef]
- McGEOCH, M.A. The selection, testing and application of terrestrial insects as bioindicators. Biol. Rev. 1998, 73, 181–201. [Google Scholar] [CrossRef]
- Menéndez, R. How are insects responding to global warming? Tijdschr. Entomol. 2007, 150, 355. [Google Scholar]
- Li, Y.; Pang, H.; Bocak, L. A review of the neotenic genus Atelius Waterhouse, 1878 from China (Coleoptera: Lycidae). Ann. Zool. 2018, 68, 351–356. [Google Scholar] [CrossRef]
- Deutsch, C.A.; Tewksbury, J.J.; Huey, R.B.; Sheldon, K.S.; Ghalambor, C.K.; Haak, D.C.; Martin, P.R. Impacts of climate warming on terrestrial ectotherms across latitude. Proc. Natl. Acad. Sci. USA 2008, 105, 6668–6672. [Google Scholar] [CrossRef]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef]
- Pounds, J.; Fogden, M.; Masters, K. Responses of natural communities to climate change in a highland tropical forest. Case Study. See Lovejoy Hannah 2005, 2005, 70–74. [Google Scholar]
- Wilson, R.J.; Gutiérrez, D.; Gutiérrez, J.; Martínez, D.; Agudo, R.; Monserrat, V.J. Changes to the elevational limits and extent of species ranges associated with climate change. Ecol. Lett. 2005, 8, 1138–1146. [Google Scholar] [CrossRef] [PubMed]
- Belfer-Cohen, A. After the ice: A global human history—20,000-5,000 BC. J. R. Anthropol. Inst. 2004, 10, 714–715. [Google Scholar]
- Lambeck, K.; Chappell, J. Sea level change through the last glacial cycle. Science 2001, 292, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Chen, C.; Zhao, Q.; Wei, J.; Zhang, H. Identifying Potentially Climatic Suitability Areas for Arma custos (Hemiptera: Pentatomidae) in China under Climate Change. Insects 2020, 11, 674. [Google Scholar] [CrossRef]
- Wang, F.; Wang, D.; Guo, G.; Zhang, M.; Lang, J.; Wei, J. Potential distributions of the invasive barnacle scale Ceroplastes cirripediformis (Hemiptera: Coccidae) under climate change and implications for its management. J. Econ. Entomol. 2021, 114, 82–89. [Google Scholar] [CrossRef]
- Xu, D.; Li, X.; Jin, Y.; Zhuo, Z.; Yang, H.; Hu, J.; Wang, R. Influence of climatic factors on the potential distribution of pest Heortia vitessoides Moore in China. Glob. Ecol. Conserv. 2020, 23, e01107. [Google Scholar] [CrossRef]
- Ji, W.; Han, K.; Lu, Y.; Wei, J. Predicting the potential distribution of the vine mealybug, Planococcus ficus under climate change by MaxEnt. Crop Protect. 2020, 137, 105268. [Google Scholar] [CrossRef]
- Meulebrouck, K.; Ameloot, E.; Verheyen, K.; Hermy, M. Local and regional factors affecting the distribution of the endangered holoparasite Cuscuta epithymum in heathlands. Biol. Conserv. 2007, 140, 8–18. [Google Scholar] [CrossRef]
- Society, R. Biodiversity-Climate Interactions: Adaptation, Mitigation and Human Livelihoods; The Clyvedon Press: Cardiff, UK, 2008. [Google Scholar]
- Coope, G. Interpretations of Quaternary insect fossils. Annu. Rev. Entomol. 1970, 15, 97–121. [Google Scholar] [CrossRef]
- Coope, G. The response of late quaternary insect communities to sudden climatic changes. In Organization of Communities, Past and Present; Gee, J., Giller, P., Eds.; Blackwell Scientific Publications: Oxford, UK, 1987; pp. 421–438. [Google Scholar]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Glob. Chang. Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- Willott, S.; Hassall, M. Life-history responses of British grasshoppers (Orthoptera: Acrididae) to temperature change. Funct. Ecol. 1998, 12, 232–241. [Google Scholar] [CrossRef]
- Hodkinson, I.; Bird, J.; Miles, J.; Bale, J.; Lennon, J. Climatic signals in the life histories of insects: The distribution and abundance of heather psyllids (Strophingia spp.) in the UK. Funct. Ecol. 1999, 13, 83–95. [Google Scholar] [CrossRef]
- Guo, K.; Hao, S.G.; Sun, J.; Kang, L.E. Differential responses to warming and increased precipitation among three contrasting grasshopper species. Glob. Chang. Biol. 2009, 15, 2539–2548. [Google Scholar] [CrossRef]
- Parmesan, C.; Ryrholm, N.; Stefanescu, C.; Hill, J.K.; Thomas, C.D.; Descimon, H.; Huntley, B.; Kaila, L.; Kullberg, J.; Tammaru, T. Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature 1999, 399, 579–583. [Google Scholar] [CrossRef]
- Kazakis, G.; Ghosn, D.; Vogiatzakis, I.; Papanastasis, V. Vascular plant diversity and climate change in the alpine zone of the Lefka Ori, Crete. Biodivers. Conserv. 2007, 16, 1603–1615. [Google Scholar] [CrossRef]
- Pickering, C.; Hill, W.; Green, K. Vascular plant diversity and climate change in the alpine zone of the Snowy Mountains, Australia. Biodivers. Conserv. 2008, 17, 1627–1644. [Google Scholar] [CrossRef]
- Stevens, G.C. The elevational gradient in altitudinal range: An extension of Rapoport’s latitudinal rule to altitude. Am. Nat. 1992, 140, 893–911. [Google Scholar] [CrossRef]
- Körner, C. The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef]
- Wiens, J.J.; Parra-Olea, G.; García-París, M.; Wake, D.B. Phylogenetic history underlies elevational biodiversity patterns in tropical salamanders. Proc. R. Soc. B 2007, 274, 919–928. [Google Scholar] [CrossRef]
- Werenkraut, V.; Ruggiero, A. Quality of basic data and method to identify shape affect richness–altitude relationships in meta-analysis. Ecology 2011, 92, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Matsuhisa, S. Population abundance gradient of Inurois punctigera along altitude. Entomol. Sci. 2020, 23, 23–27. [Google Scholar] [CrossRef]
- Davis, M.B.; Shaw, R.G. Range shifts and adaptive responses to Quaternary climate change. Science 2001, 292, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef]
- Yamamoto, S.; Sota, T. Incipient allochronic speciation by climatic disruption of the reproductive period. Proc. R. Soc. B 2009, 276, 2711–2719. [Google Scholar] [CrossRef] [PubMed]
- Lawson, A.M.; Weir, J.T. Latitudinal gradients in climatic-niche evolution accelerate trait evolution at high latitudes. Ecol. Lett. 2014, 17, 1427–1436. [Google Scholar] [CrossRef] [PubMed]
- Boumans, L.; Hogner, S.; Brittain, J.; Johnsen, A. Ecological speciation by temporal isolation in a population of the stonefly Leuctra hippopus (Plecoptera, Leuctridae). Ecol. Evol. 2017, 7, 1635–1649. [Google Scholar] [CrossRef]
- Polato, N.R.; Gill, B.A.; Shah, A.A.; Gray, M.M.; Casner, K.L.; Barthelet, A.; Messer, P.W.; Simmons, M.P.; Guayasamin, J.M.; Encalada, A.C. Narrow thermal tolerance and low dispersal drive higher speciation in tropical mountains. Proc. Natl. Acad. Sci. USA 2018, 115, 12471–12476. [Google Scholar] [CrossRef]
- Yuan, M.L.; Zhang, Q.L.; Zhang, L.; Jia, C.L.; Li, X.P.; Yang, X.Z.; Feng, R.Q. Mitochondrial phylogeny, divergence history and high-altitude adaptation of grassland caterpillars (Lepidoptera: Lymantriinae: Gynaephora) inhabiting the Tibetan Plateau. Mol. Phylogen. Evol. 2018, 122, 116–124. [Google Scholar] [CrossRef]
- Santos, H.; Paiva, M.; Tavares, C.; Kerdelhue, C.; Branco, M. Temperature niche shift observed in a Lepidoptera population under allochronic divergence. J. Evol. Biol. 2011, 24, 1897–1905. [Google Scholar] [CrossRef]
- Suzuki, T.; Suzuki, N.; Tojo, K. Parallel evolution of an alpine type ecomorph in a scorpionfly: Independent adaptation to high-altitude environments in multiple mountain locations. Mol. Ecol. 2019, 28, 3225–3240. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Liu, T.; Li, L.; Zhao, Y.; Pei, L.; Zhao, J. Predicting the potential distribution of Polygala tenuifolia Willd. under climate change in China. PLoS ONE 2016, 11, e0163718. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Yao, L.; Meng, J.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef]
- Li, J.; Fan, G.; He, Y. Predicting the current and future distribution of three Coptis herbs in China under climate change conditions, using the MaxEnt model and chemical analysis. Sci. Total Environ. 2020, 698, 134141. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, P.; Erwin, T.L.; Borges, P.A.; New, T.R. The seven impediments in invertebrate conservation and how to overcome them. Biol. Conserv. 2011, 144, 2647–2655. [Google Scholar] [CrossRef]
- Samways, M.J. Future-proofing insect diversity. Curr. Opin. Insect Sci. 2015, 12, 71–78. [Google Scholar] [CrossRef]
- Rocha-Ortega, M.; Rodríguez, P.; Bried, J.; Abbott, J.; Córdoba-Aguilar, A. Why do bugs perish? Range size and local vulnerability traits as surrogates of Odonata extinction risk. Proc. R. Soc. B 2020, 287, 20192645. [Google Scholar] [CrossRef]
- Gaston, K.J.; Fuller, R.A. The sizes of species’ geographic ranges. J. Appl. Ecol. 2009, 46, 1–9. [Google Scholar] [CrossRef]
- Boys, W.A.; Adam, M. Predicting the distributions of regional endemic dragonflies using a combined model approach. Insect Conserv. Divers. 2021, 14, 52–66. [Google Scholar] [CrossRef]
- Settele, J.; Kühn, E. Insect conservation. Science 2009, 325, 41–42. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Description | Percent Contribution |
|---|---|---|
| Bio1 | Annual mean temperature | 6.1 |
| Bio2 | Mean diurnal range (mean of monthly (max temp-min temp)) | 1.4 |
| Bio3 | Isothermality (Bio2/Bio7)(×100) | 2.3 |
| Bio4 | Temperature seasonality (standard deviation ×100) | 3.5 |
| Bio5 | Maximum temperature of warmest month | 0.6 |
| Bio6 | Minimum temperature of coldest month | 8.1 |
| Bio7 | Temperature annual range (Bio5-Bio6) | 0.2 |
| Bio8 | Mean temperature of wettest quarter | 0.4 |
| Bio9 | Mean temperature of driest quarter | 0.2 |
| Bio10 | Mean temperature of warmest quarter | 1.3 |
| Bio11 | Mean temperature of coldest quarter | 15.1 |
| Bio12 | Annual precipitation | 46.3 |
| Bio13 | Precipitation of wettest month | 0.1 |
| Bio14 | Precipitation of driest month | 13.3 |
| Bio15 | Precipitation seasonality (coefficient of variation) | 0.2 |
| Bio16 | Precipitation of wettest quarter | 0.1 |
| Bio17 | Precipitation of driest quarter | 0.2 |
| Bio18 | Precipitation of warmest quarter | 0.2 |
| Bio19 | Precipitation of coldest quarter | 0.3 |
| Species Richness | ||||
|---|---|---|---|---|
| CoefOLS | R2OLS | CoefSAR | Pseudo R2SAR | |
| Bio1 | 0.043 | 0.098 ns | 0.003 | 52.538 ns |
| Bio2 | −0.086 | 0.660 ** | −0.064 | 52.385 ** |
| Bio3 | −0.010 | 0.009 ns | 0.018 | 53.273 ns |
| Bio4 | −0.056 | 0.232 ns | −0.040 | 52.893 ns |
| Bio5 | −0.027 | 0.007 ns | −0.057 | 52.977 * |
| Bio6 | 0.054 | 0.201 ns | 0.028 | 53.071 ns |
| Bio7 | −0.072 | 0.427 * | −0.056 | 53.088 * |
| Bio8 | 0.049 | 0.158 ns | −0.041 | 52.676 ns |
| Bio9 | −0.031 | 0.096 ns | −0.006 | 52.744 ns |
| Bio10 | 0.020 | 0.040 ns | −0.043 | 52.637 ns |
| Bio11 | 0.048 | 0.139 ns | 0.021 | 53.074 ns |
| Bio12 | 0.155 | 2.318 *** | 0.064 | 51.806 ** |
| Bio13 | 0.148 | 2.091 *** | 0.030 | 51.205 ns |
| Bio14 | 0.013 | 0.017 ns | 0.066 | 51.552 ** |
| Bio15 | 0.036 | 0.042 ns | −0.064 | 51.482 ** |
| Bio16 | 0.160 | 2.480 *** | 0.040 | 51.141 ns |
| Bio17 | 0.008 | 0.006 ns | 0.064 | 51.625 ** |
| Bio18 | 0.238 | 5.581 *** | 0.087 | 50.743 *** |
| Bio19 | −0.072 | 0.4291 * | 0.023 | 52.439 ns |
| Elevation | 0.183 | 3.246 *** | 0.077 | 51.563 ** |
| Data Source | Category | Variables | Abbreviation | Units |
|---|---|---|---|---|
| Panorpidae Worldclim | Bioclimatic | Mean diurnal range | Bio2 | °C |
| Isothermality (Bio2/Bio7)(×100) | Bio3 | % | ||
| Mean temperature of warmest quarter | Bio10 | °C | ||
| Mean temperature of coldest quarter | Bio11 | °C | ||
| Annual precipitation | Bio12 | mm | ||
| Precipitation of driest month | Bio14 | mm |
| Habitat Suitability | Historical Periods | Global Warming Scenarios | ||
|---|---|---|---|---|
| Last Interglacial | Last Glacial Maximum | 1970–2000 | 2070s RCP4.5 | |
| Area (million km2) | Area (million km2) | Area (million km2) | Area (million km2) | |
| 0–0.2 | 114.7584 | 94.8089 | 111.2099 | 112.3447 |
| 0.2–0.4 | 13.1229 | 16.8937 | 13.2336 | 12.8701 |
| 0.4–0.6 | 6.5181 | 14.5506 | 8.3257 | 7.9061 |
| >0.6 | 14.5005 | 22.6467 | 16.1308 | 15.7791 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, J.; Liu, W.; Hu, F.; Miao, P.; Xing, L.; Hua, Y. The Distribution Pattern and Species Richness of Scorpionflies (Mecoptera: Panorpidae). Insects 2023, 14, 332. https://doi.org/10.3390/insects14040332
Su J, Liu W, Hu F, Miao P, Xing L, Hua Y. The Distribution Pattern and Species Richness of Scorpionflies (Mecoptera: Panorpidae). Insects. 2023; 14(4):332. https://doi.org/10.3390/insects14040332
Chicago/Turabian StyleSu, Jian, Wanjing Liu, Fangcheng Hu, Panpan Miao, Lianxi Xing, and Yuan Hua. 2023. "The Distribution Pattern and Species Richness of Scorpionflies (Mecoptera: Panorpidae)" Insects 14, no. 4: 332. https://doi.org/10.3390/insects14040332
APA StyleSu, J., Liu, W., Hu, F., Miao, P., Xing, L., & Hua, Y. (2023). The Distribution Pattern and Species Richness of Scorpionflies (Mecoptera: Panorpidae). Insects, 14(4), 332. https://doi.org/10.3390/insects14040332