

Triatoma yelapensis sp. nov. (Hemiptera: Reduviidae) from Mexico, with a Key of Triatoma Species Recorded in Mexico
 , , and
, , and
Abstract
Simple Summary
Abstract
1. Introduction
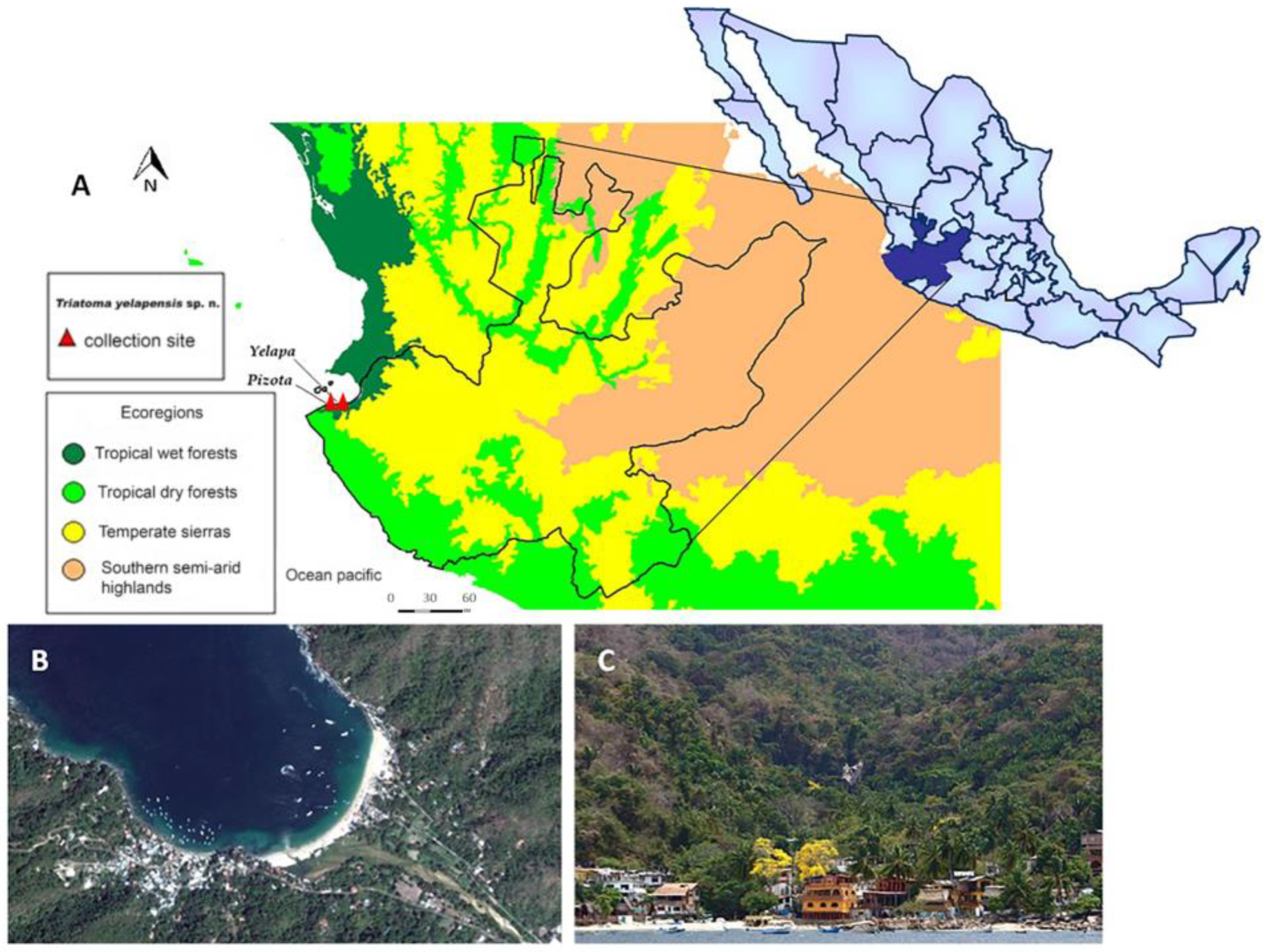
2. Materials and Methods
3. Results
3.1. Taxonomy
- Familia Reduviidae Latreille, 1807
- Subfamilia Triatominae Jeannel, 1919
- Genero Triatoma Laporte, 1832
- Triatoma yelapensis sp. nov.
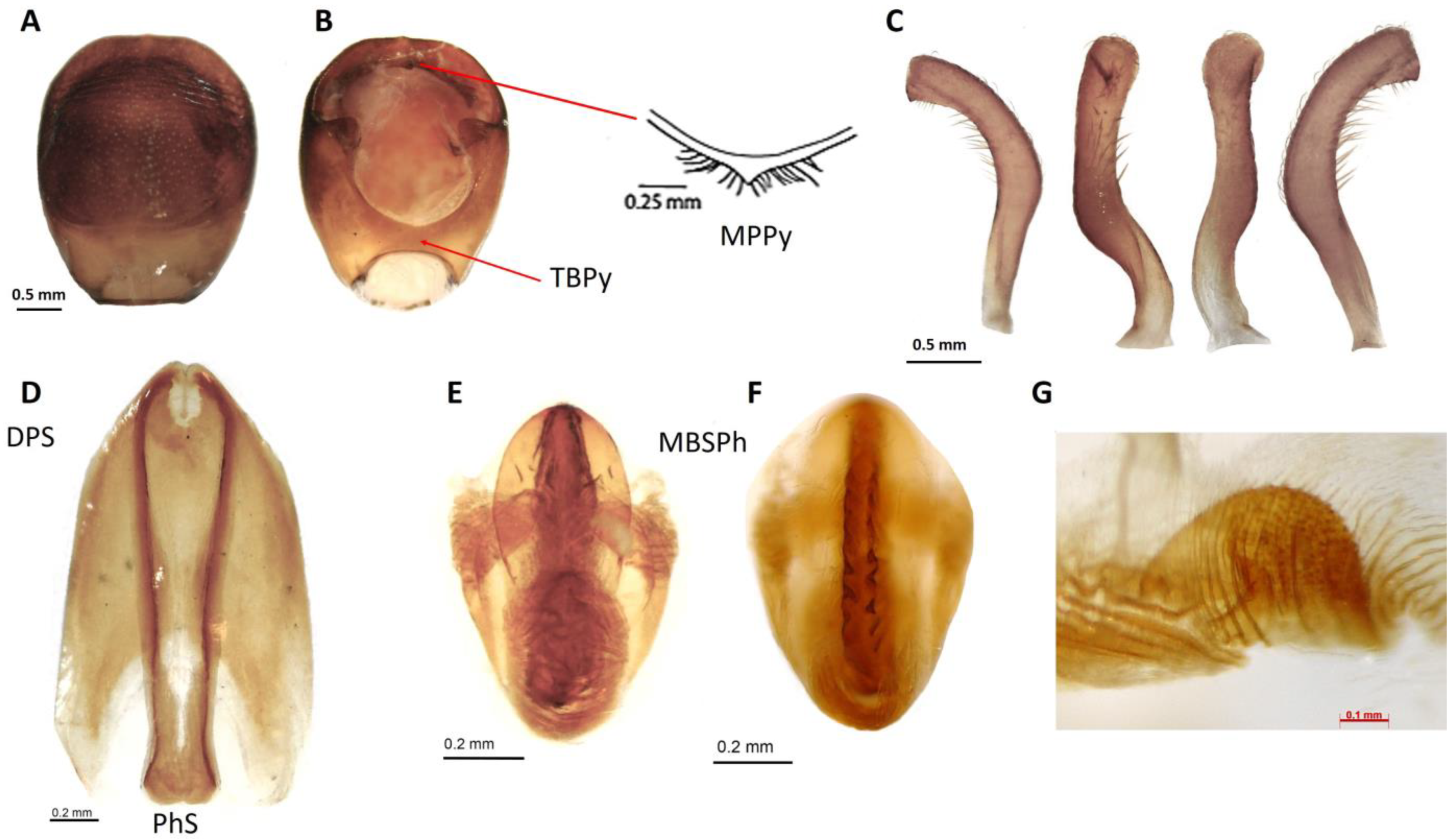
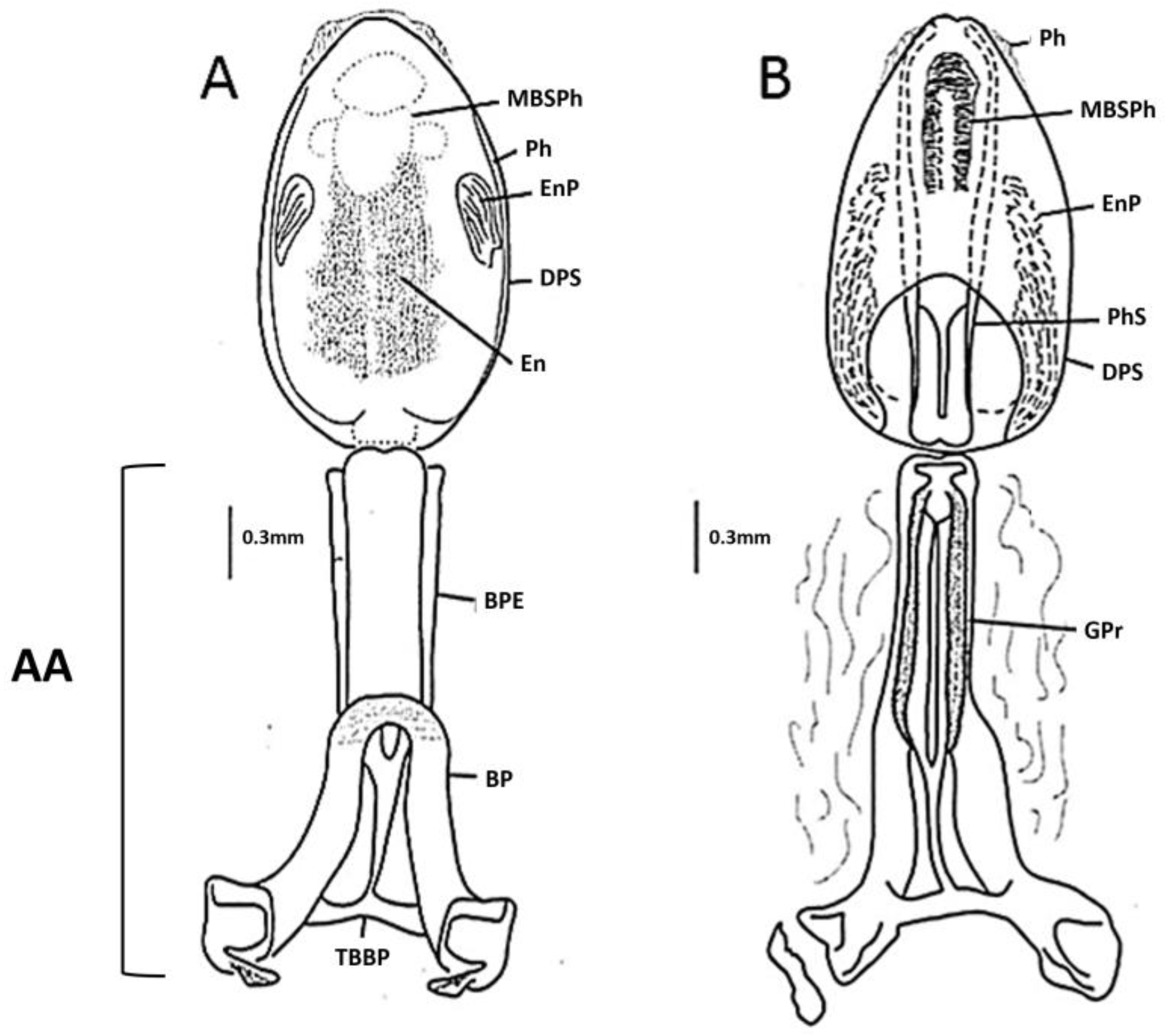
3.2. Description
3.2.1. Coloration
3.2.2. Morphological Features
3.3. Etymology
3.4. Host–Parasite Data
3.5. Species Group Assignment
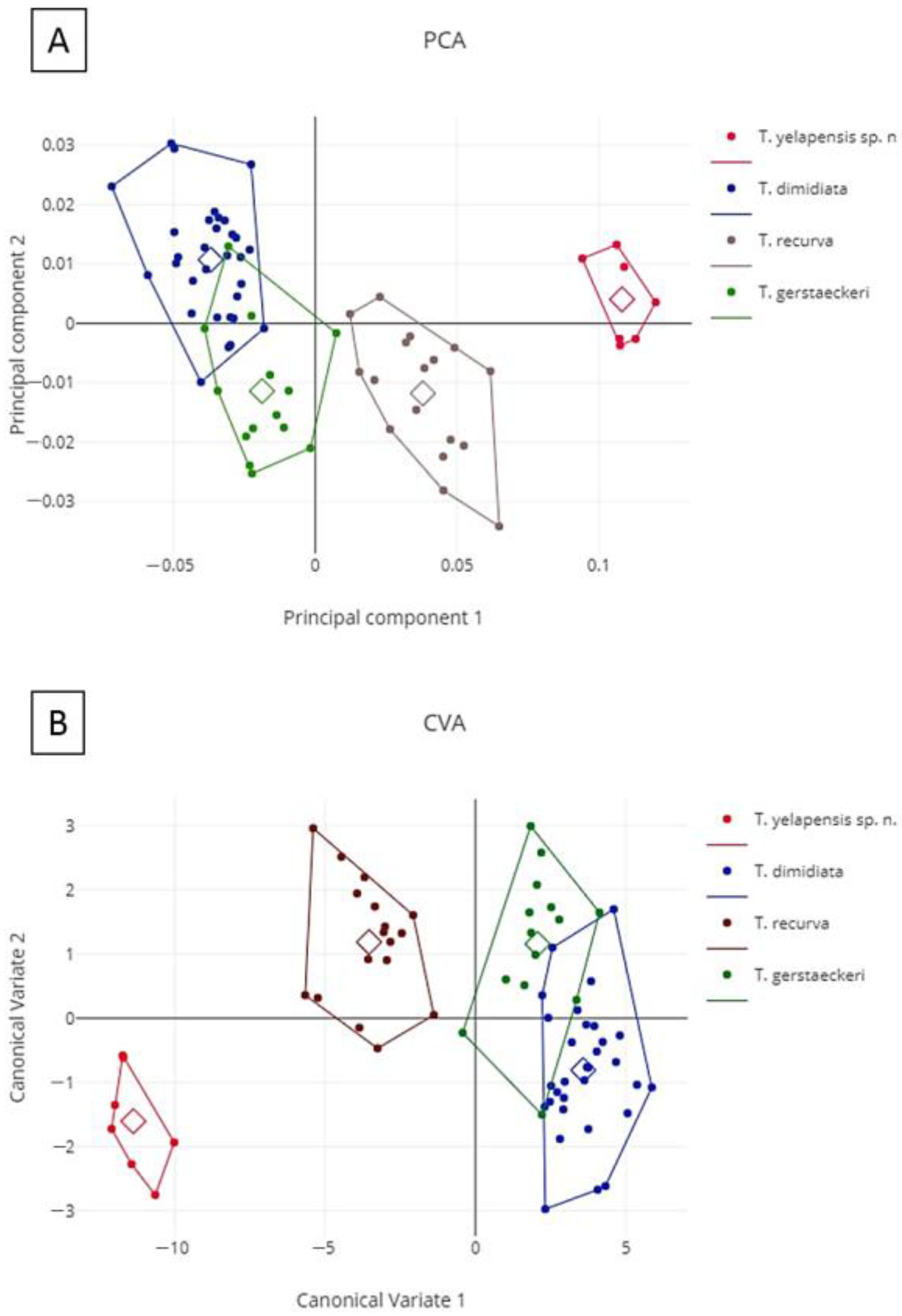
3.6. Morphometry
3.7. Key to the Species of Triatoma recorded from Mexico (based on Lent and Wygodzisnky [7], Alejandre-Aguilar et al. [32], and Rengifo-Correa et al. [20])
| T. mexicana |
| 2 |
| 3 |
| 15 |
| 4 |
| 9 |
| 5 |
| T. picturata |
| 6 |
| T. pallidipennis |
| 7 |
| T. longipennis |
| 8 |
| T. mazzottii |
| T. phyllosoma |
| T. bassolsae |
| T. recurva |
| 10 |
| T. hegneri |
| 11 |
| T. gerstaeckeri (part) |
| 12 |
| T. yelapensis sp. nov |
| 13 |
| T. huehuetenanguensis |
| 14 |
| T. mopan |
| T. dimidiata |
| T. sanguisuga |
| 16 |
| 17 |
| 19 |
| T. rubida |
| 18 |
| T. nitida |
| T. neotomae |
| T. brailovskyi |
| 20 |
| 21 |
| 25 |
| 22 |
| 23 |
| T. gomeznunezi |
| T. bolivari |
| T. protracta |
| 24 |
| T. peninsularis |
| T. sinaloensis |
| 26 |
| 28 |
| T. infestans |
| 27 |
| T. gerstaeckeri |
| T. indictiva |
| T. incrassata |
| T. barberi |
4. Discussion and Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- PAHO. La Enfermedad de Chagas. Organización Panamericana de la Salud. 2017. Available online: http://www.paho.org/hq/index.php?option=com_topics&view=article&id=10&Itemid=40743&lang=es (accessed on 11 September 2022).
- Ramsey, J.M.; Tello, L.A.; Pohls, J.L. Actualidades sobre la epidemiología de la enfermedad de Chagas en México. In Iniciativa para la Vigilancia y el Control de la Enfermedad de Chagas en la República Mexicana; Ramsey, J.M., Tello-López, A., Pohls, J.L., Eds.; INSP: Cuernavaca, Mexico, 2003; pp. 85–103. [Google Scholar]
- Rojo-Medina, J.; Ruiz-Matus, C.; Salazar-Schettino, P.M.; González-Roldan, J.F. Enfermedad de Chagas en México. Gac. Méd. Méx. 2018, 154, 605–612. [Google Scholar] [CrossRef]
- Dorn, P.L.; Justi, S.A.; Dale, C.; Stevens, L.; Galvão, C.; Lima-Cordón, R.; Monroy, C. Description of Triatoma mopan sp. n. from a cave in Belize (Hemiptera, Reduviidae, Triatominae). Zookeys 2018, 775, 69–95. [Google Scholar] [CrossRef]
- Galvão, C.; Carcavallo, R.U.; Rocha, D.S.; Jurberg, J. A checklist of the current valid species of the subfamily Triatominae Jeannel, 1919 (Hemiptera: Reduviidae) and their geographical distribution, with nomenclatural and taxonomic notes. Zootaxa 2003, 202, 1–36. [Google Scholar] [CrossRef]
- Justi, S.A.; Galvão, C. The evolutionary origin of diversity in Chagas disease vectors. Trends Parasitol. 2017, 33, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Lent, H.; Wygodzynsky, P. Revision of the Triatominae (Hemiptera–Reduviidae) and their significance as vectors of Chagas’ disease. Bull. Am. Mus. Nat. Hist. 1979, 163, 123–520. [Google Scholar]
- Lima-Cordón, R.A.; Monroy, M.C.; Stevens, L.; Rodas, A.; Rodas, G.A.; Dorn, P.L.; Justi, S.A. Description of Triatoma huehuetenanguensis sp. n., a potential Chagas disease vector (Hemiptera, Reduviidae, Triatominae). ZooKeys 2019, 820, 51–70. [Google Scholar] [CrossRef]
- Nascimiento, J.D.; da Rosa, J.A.; Salgado-Roa, F.C.; Hernández, C.; Pardo-Diaz, C.; Alevi, K.C.C.; Ravazi, A.; de Oliveira, J.; de Azeredo Oliveira, M.T.V.; Salazar, C.; et al. Taxonomical over splitting in the Rhodnius prolixus (Insecta: Hemiptera: Reduviidae) clade: Are R. taquarussuensis (da Rosa et al. 2017) and R. neglectus (Lent, 1954) the same species? PLoS ONE 2019, 14, e0211285. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.; Ayala, J.M.; Justi, S.A.; Rosa, J.A.; Galvão, C. Description of a new species of Nesotriatoma Usinger, 1944 from Cuba and revalidation of synonymy between Nesotriatoma bruneri (Usinger, 1944) and N. flavida (Neiva, 1911) (Hemiptera, Reduviidae, Triatominae). J. Vector Ecol. 2018, 43, 148–157. [Google Scholar] [CrossRef]
- Poinar, G., Jr. A primitive triatomine bug, Paleotriatoma metaxytaxa gen. et sp. nov. (Hemiptera: Reduviidae: Triatominae), in mid-Cretaceous amber from northern Myanmar. Cretac. Res. 2019, 93, 90–97. [Google Scholar] [CrossRef]
- Rosa, J.A.; Justino, H.H.G.; Nascimiento, J.D.; Mendoça, V.J.; Rocha, C.S.; Carvalho, D.B.; Falcone, R.; Azeredo-Oliveira, M.T.V.; Alevi, K.C.C.; Oliveira, J. A new species of Rhodnius from Brazil (Hemiptera, Reduviidae, Triatominae). Zookeys 2017, 675, 1–25. [Google Scholar] [CrossRef]
- Zhao, Y.; Galvão, C.; Cai, W. Rhodnius micki, a new species of Triatominae (Hemiptera, Reduviidae) from Bolivia. Zookeys 2021, 1012, 71–93. [Google Scholar] [CrossRef] [PubMed]
- Alevi, K.C.C.; da Silva Rocha, D.; Galvão, C. Trends in Taxonomy of Chagas Disease Vectors (Hemiptera, Reduviidae, Triatominae): From Linnaean to Integrative Taxonomy. Pathogens 2021, 10, 1627. [Google Scholar] [CrossRef] [PubMed]
- Correia, J.P.S.O.; Gil-Santana, H.R.; Dale, C.; Galvão, G. Triatoma guazu Lent and Wygodzinsky Is a Junior Synonym of Triatoma williami Galvão, Souza and Lima. Insects 2022, 13, 591. [Google Scholar] [CrossRef]
- Rivas, N.; González-Guzmán, S.; Alejandre-Aguilar, R. First record of Triatoma barberi Usinger, 1939 (Hemiptera: Reduviidae) in northern State of Mexico, Mexico. J. Vector Ecol. 2018, 43, 337–339. [Google Scholar] [CrossRef]
- Espinosa-González, C.A.; Dávalos-Becerril, E.; Correa-Morales, F.; González-Acosta, C.; López-Bello, R.; Alvarado-Estrada, J.M.; Esquinca-Calvo, L.M.; García-Gutiérrez, C.V.; Gómez-Maldonado, C.A.; Cuevas-González, E.; et al. New records of Triatoma huehuetenanguensis in an urban area of Southwest Mexico. J. Vector Borne Dis. 2022, 59, 86–90. [Google Scholar] [CrossRef]
- Ramsey, J.M.; Peterson, A.T.; Carmona-Castro, O.; Moo-Llanes, D.A.; Nakazawa, Y.; Butrick, M.; Tun-Ku, E.; De la Cruz-Félix, K.; Ibarra-Cerdeña, C.N. Atlas of Mexican Triatominae (Reduviidae:Hemiptera) and vector transmission of Chagas disease. Mem. Inst. Oswaldo Cruz 2015, 110, 339–352. [Google Scholar] [CrossRef]
- Rengifo-Correa, L.; Abad-Franch, F.; Martínez-Hernández, F.; Salazar-Schettino, P.M.; Téllez-Rendón, J.L.; Villalobos, G.; Morrone, J.J. A Biogeographic-ecological approach to disentangle reticulate evolution in the Triatoma phyllosoma species group (Heteroptera: Triatominae), vectors of Chagas disease. J. Zool. Syst. Evol. 2020, 59, 1–17. [Google Scholar] [CrossRef]
- Rengifo-Correa, L.; Téllez-Rendón, J.L.; Esteban, L.; Huerta, H.; Morrone, J.J. The Triatoma phyllosoma species group (Hemiptera: Reduviidae: Triatominae), vectors of Chagas disease: Diagnoses and a key to the species. Zootaxa 2021, 3, 335–365. [Google Scholar] [CrossRef]
- Brenière, S.F.; Bosseno, M.F.; Magallón-Gastelúm, E.; Castillo Ruvalcaba, E.G.; Gutierrez, M.S.; Montaño Luna, E.C.; Basulto, J.T.; Mathieu-Daudé, F.; Walter, A.; Lozano-Kasten, F. Peridomestic colonization of Triatoma longipennis (Hemiptera, Reduviidae) and Triatoma barberi (Hemiptera, Reduviidae) in a rural community with active transmission of Trypanosoma cruzi in Jalisco state, Mexico. Acta Trop. 2007, 101, 249–257. [Google Scholar] [CrossRef]
- Magallón-Gastélum, E.; Lozano-Kasten, F.; Gutierréz, M.S.; Flores-Pérez, A.; Sánchez, B.; Espinoza, B.; Bosseno, M.F.; Brenière, S.F. Epidemiological risk for Trypanosoma cruzi transmission by species of Phyllosoma complex in the occidental part of Mexico. Acta Trop. 2006, 97, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Magallón-Gastelum, E.; Peñaloza Magdaleno, N.C.; Katthain-Duchateau, G.; Trujillo-Contreras, F.; Lozano-Kasten, F.J.; Hernández-Gutiérrez, R.J. Distribution of Chagas disease vectors (Hemiptera: Reduviidae: Triatominae), in the state of Jalisco, Mexico. Rev. Bioméd. 1998, 9, 151–157. [Google Scholar]
- Martínez-Ibarra, J.A.; Martínez-Grant, J.A.; Verdugo-Cervantes, M.R.; Bustos-ldaña, R.; Nogueda-Torres, B. Vigilancia de la presencia de triatominos mediante gallineros en el sur de Jalisco, México. Biomédica 2010, 30, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ibarra, J.A.; Martínez-Hernández, F.; Villalobos, V.; Vences-Blanco, M.O.; Salazar-Schettino, P.M. Update on the distribution of Triatoma bolivari and Triatoma brailovskyi (Hemiptera: Reduviidae: Triatominae) in western Mexico. J. Vector Ecol. 2010, 35, 432–434. [Google Scholar] [CrossRef]
- Zarate, L.G.; Zarate, R.J. A checklist of the Triatominae (Hemiptera: Reduviidae) of Mexico. Int. J. Entomol. 1985, 27, 102–127. [Google Scholar]
- INEGI. Instituto Nacional de Estadística y Geografía. Prontuario de Información Geográfica Municipal de los Estados Unidos Mexicanos, Cabo Corrientes, Jalisco, CG14020. 2009. Available online: http://mapserver.inegi.org.mx/mgn2k/ (accessed on 15 September 2022).
- Weirauch, C. Cladistic analysis of Reduviidae (Heteroptera: Cimicomorpha) based on morphological characters. Syst. Entomol. 2008, 33, 229–274. [Google Scholar] [CrossRef]
- May-Concha, I.; Guerenstein, P.G.; Ramsey, J.M.; Rojas, J.C.; Catalá, S. Antennal phenotype of the Triatoma dimidiata complex, vectors of Chagas disease. Infect. Genet. Evol. 2016, 40, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Dujardin, J.P. Morphometrics applied to medical entomology. Infect. Genet. Evol. 2008, 8, 875–890. [Google Scholar] [CrossRef] [PubMed]
- Dujardin, S.; Dujardin, J.P. Geometric morphometrics in the cloud. Infect. Genet. Evol. 2019, 70, 189–196. [Google Scholar] [CrossRef]
- Aguillar, R.A.; Torres, B.N.; Jímenez, M.C.; Jurberg, J.; Galvão, C.; Carcavallo, R. Triatoma bassolsae sp. n. do México, com uma chave para as espécies do complexo phyllosoma (Hemiptera, Reduviidae). Mem. Inst. Oswaldo Cruz 1999, 94, 353–359. [Google Scholar] [CrossRef]
- Nattero, J.; Piccinali, R.V.; Macedo, C.; Hernández, M.L.; Abrahan, L.; Lobbia, P.A.; Rodriguez, C.S.; Carbajal de la Fuente, A.L. Morphometric variability among the species of the Sordida subcomplex (Hemiptera: Reduviidae: Triatominae): Evidence for differentiation across the distribution range of Triatoma sordida. Parasit. Vectors 2017, 10, 412. [Google Scholar] [CrossRef]
- Lent, H.; Jurberg, J. A genitália externa dos machos de sete espécies de Triatoma Laporte, 1832 da região Neártica (Hemiptera, Reduviidae). Mem. Inst. Oswaldo Cruz 1987, 82, 227–246. [Google Scholar] [CrossRef]
- Lent, H.; Jurberg, J. Sobre a variação intra-especifica em Triatoma dimidiata (Latreille) e Triatoma infestans (Klug) (Hemiptera, Reduviidae). Mem. Inst. Oswaldo Cruz 1985, 80, 285–299. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Character | T. yelapensis | T. recurva | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Female (4 Specimens) | Male (3 Specimens) | Holotype CAIMTriTp-0006 | Female (8 Specimens) | Male (9 Specimens) | |||||||||||||
| Mean | SD | Min | Max | Mean | SD | Min | Max | Mean | SD | Min | Max | Mean | SD | Min | Max | ||
| TL | 29.88 | (1.31) | 28 | 31 | 28.17 | (1.04) | 27 | 29 | 29 | 30.33 | (1.28) | 29 | 32 | 28.86 | (0.69) | 28 | 30 |
| HL | 5.63 | (0.13) | 5.45 | 5.76 | 5.55 | (0.14) | 5.40 | 5.67 | 5.67 | 5.01 | (0.24) | 4.65 | 5.39 | 4.81 | (0.13) | 4.61 | 4.99 |
| WH | 2.16 | (0.07) | 2.05 | 2.21 | 2.11 | (0.14) | 2.02 | 2.27 | 2.26 | 2.3 | (0.15) | 2.15 | 2.57 | 2.22 | (0.17) | 2.00 | 2.51 |
| Sy | 1.02 | (0.05) | 0.95 | 1.09 | 0.95 | (0.04) | 0.90 | 0.99 | 0.99 | 0.99 | (0.04) | 0.91 | 1.05 | 0.91 | (0.06) | 0.83 | 0.99 |
| WE | 0.58 | (0.02) | 0.56 | 0.60 | 0.60 | (0.03) | 0.56 | 0.62 | 0.62 | 0.66 | (0.07) | 0.57 | 0.82 | 0.67 | (0.07) | 0.56 | 0.78 |
| ARL | 3.51 | (0.10) | 3.36 | 3.58 | 3.44 | (0.06) | 3.37 | 3.48 | 3.48 | 2.96 | (0.20) | 2.63 | 3.26 | 2.77 | (0.11) | 2.62 | 3.01 |
| PRL | 1.09 | (0.03) | 1.06 | 1.13 | 1.05 | (0.09) | 0.98 | 1.16 | 1.16 | 0.95 | (0.09) | 0.86 | 1.12 | 0.95 | (0.06) | 0.86 | 1.03 |
| WPP | 5.84 | (0.13) | 5.71 | 5.99 | 5.97 | (0.14) | 5.82 | 6.10 | 6.09 | 6.25 | (0.44) | 5.60 | 6.84 | 6.03 | (0.50) | 5.40 | 6.74 |
| PL | 4.20 | (0.24) | 3.84 | 4.35 | 4.27 | (0.09) | 4.17 | 4.35 | 4.34 | 4.57 | (0.35) | 4.09 | 5.08 | 4.35 | (0.37) | 3.73 | 4.92 |
| lb1 | 2.22 | (0.16) | 2.07 | 2.39 | 2.07 | (0.33) | 1.70 | 2.31 | 1.96 | 2.00 | (0.14) | 1.81 | 2.21 | 1.95 | (0.15) | 1.66 | 2.12 |
| lb2 | 3.64 | (0.05) | 3.58 | 3.68 | 3.51 | (0.10) | 3.40 | 3.59 | 3.39 | 3.09 | (0.15) | 2.87 | 3.26 | 3.04 | (0.17) | 2.82 | 3.36 |
| lb3 | 0.95 | (0.04) | 0.91 | 1.01 | 0.87 | (0.07) | 0.79 | 0.92 | 0.89 | 0.91 | (0.05) | 0.83 | 0.97 | 0.88 | (0.05) | 0.78 | 0.94 |
| PedL | 1.50 | (0.06) | 1.44 | 1.55 | 1.44 | (0.03) | 1.41 | 1.46 | 1.46 | 1.18 | 1.26 | (0.08) | 1.2 | 1.35 | |||
| bfla | 3.1 | 3.34 | (0.16) | 3.23 | 3.46 | 3.46 | 3.6 | ||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Téllez-Rendón, J.; Esteban, L.; Rengifo-Correa, L.; Díaz-Albiter, H.; Huerta, H.; Dale, C. Triatoma yelapensis sp. nov. (Hemiptera: Reduviidae) from Mexico, with a Key of Triatoma Species Recorded in Mexico. Insects 2023, 14, 331. https://doi.org/10.3390/insects14040331
Téllez-Rendón J, Esteban L, Rengifo-Correa L, Díaz-Albiter H, Huerta H, Dale C. Triatoma yelapensis sp. nov. (Hemiptera: Reduviidae) from Mexico, with a Key of Triatoma Species Recorded in Mexico. Insects. 2023; 14(4):331. https://doi.org/10.3390/insects14040331
Chicago/Turabian StyleTéllez-Rendón, Juan, Lyda Esteban, Laura Rengifo-Correa, Héctor Díaz-Albiter, Herón Huerta, and Carolina Dale. 2023. "Triatoma yelapensis sp. nov. (Hemiptera: Reduviidae) from Mexico, with a Key of Triatoma Species Recorded in Mexico" Insects 14, no. 4: 331. https://doi.org/10.3390/insects14040331
APA StyleTéllez-Rendón, J., Esteban, L., Rengifo-Correa, L., Díaz-Albiter, H., Huerta, H., & Dale, C. (2023). Triatoma yelapensis sp. nov. (Hemiptera: Reduviidae) from Mexico, with a Key of Triatoma Species Recorded in Mexico. Insects, 14(4), 331. https://doi.org/10.3390/insects14040331