
Identification and Functional Characterization of General Odorant Binding Proteins in Orthaga achatina

 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing and Tissue Collection
2.2. RNA Extraction and cDNA Synthesis
2.3. Cloning, Expression and Purification of Recombinant Proteins
2.4. Phylogenetic Tree
2.5. RT-PCR and qPCR
2.6. Fluorescence Competitive Binding Assay
2.7. Homology Modeling and Molecular Docking
2.8. Data Analysis
3. Results
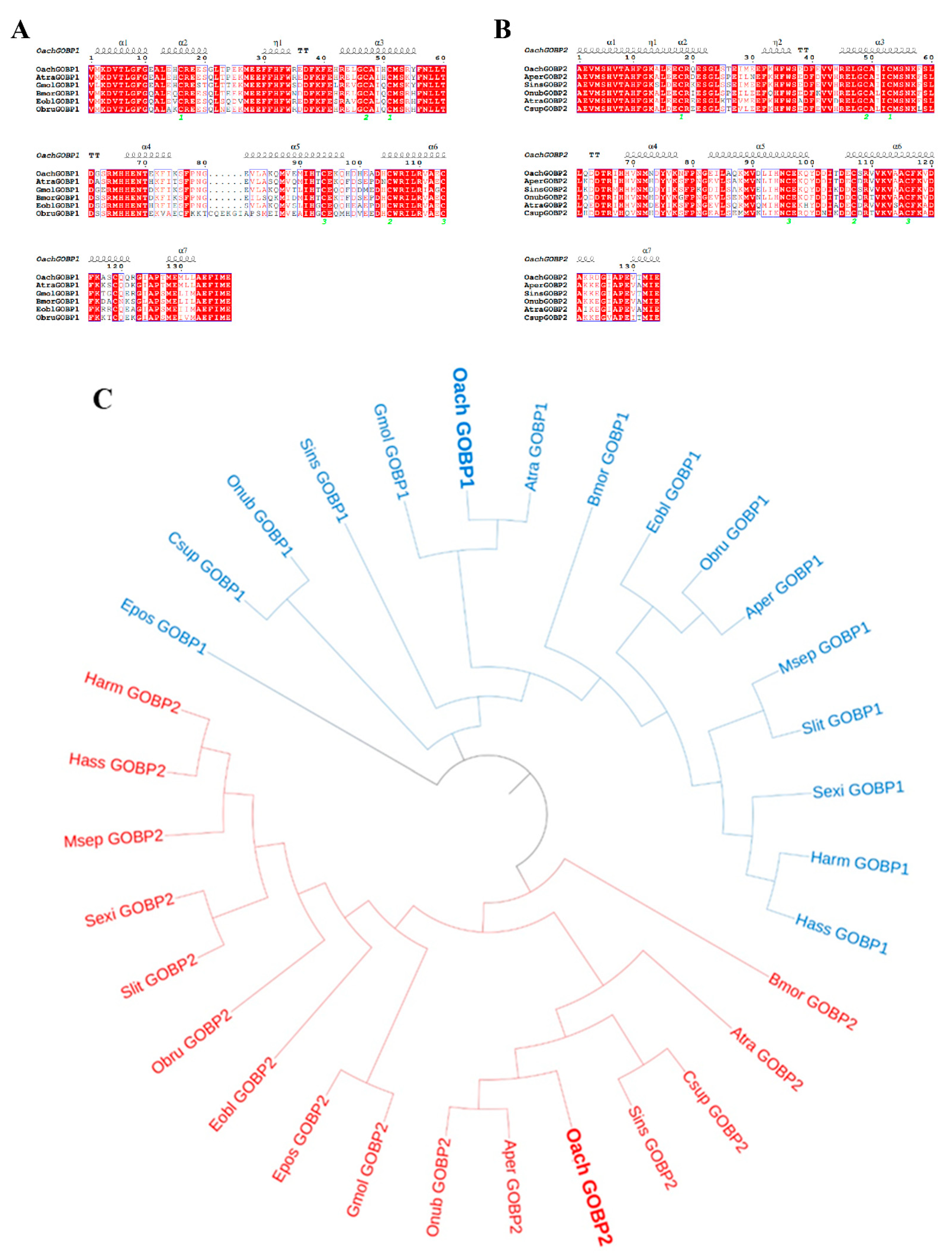
3.1. Phylogenetic and Sequence Analyses of OachGOBPs
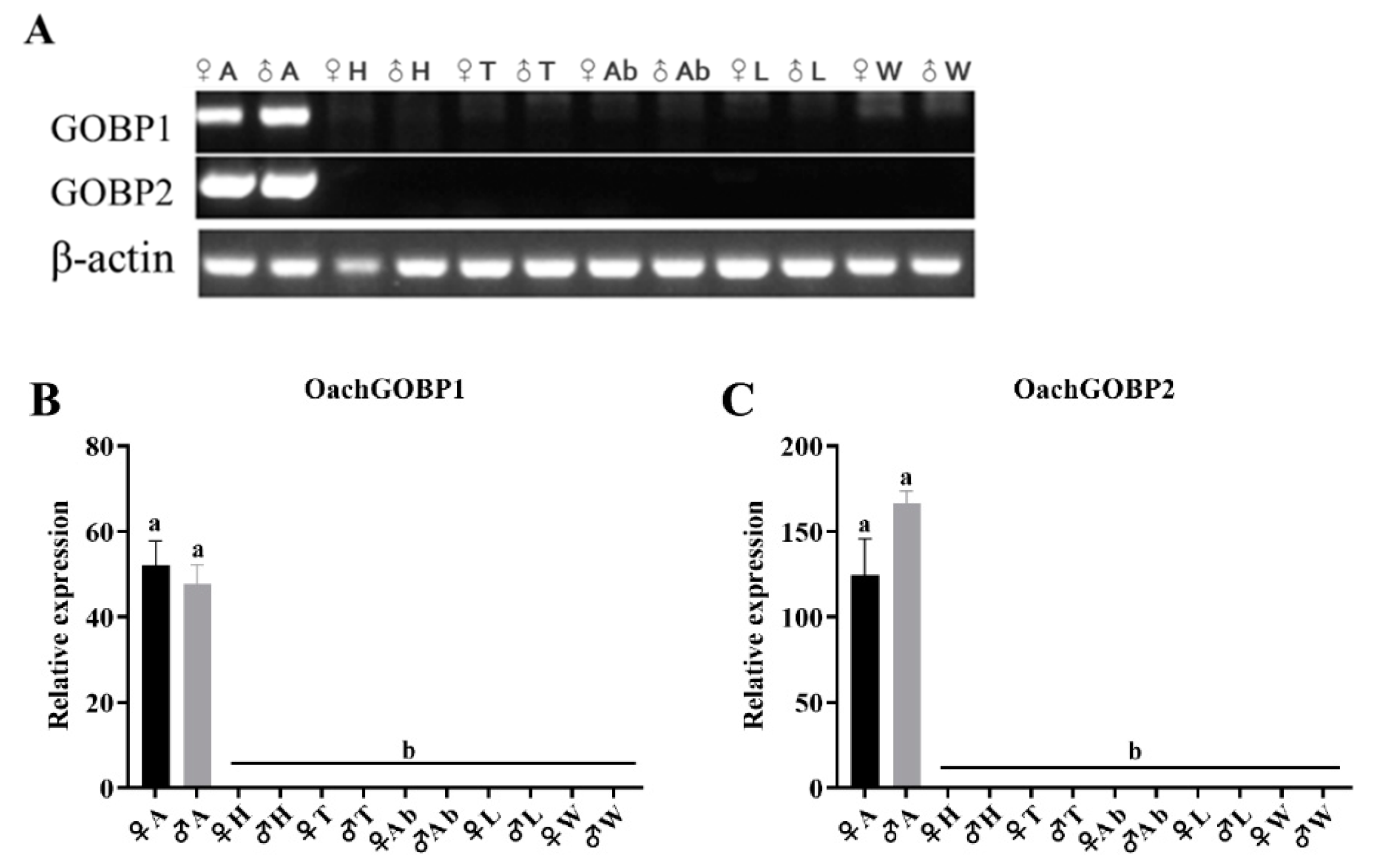
3.2. Tissue Expression Analysis
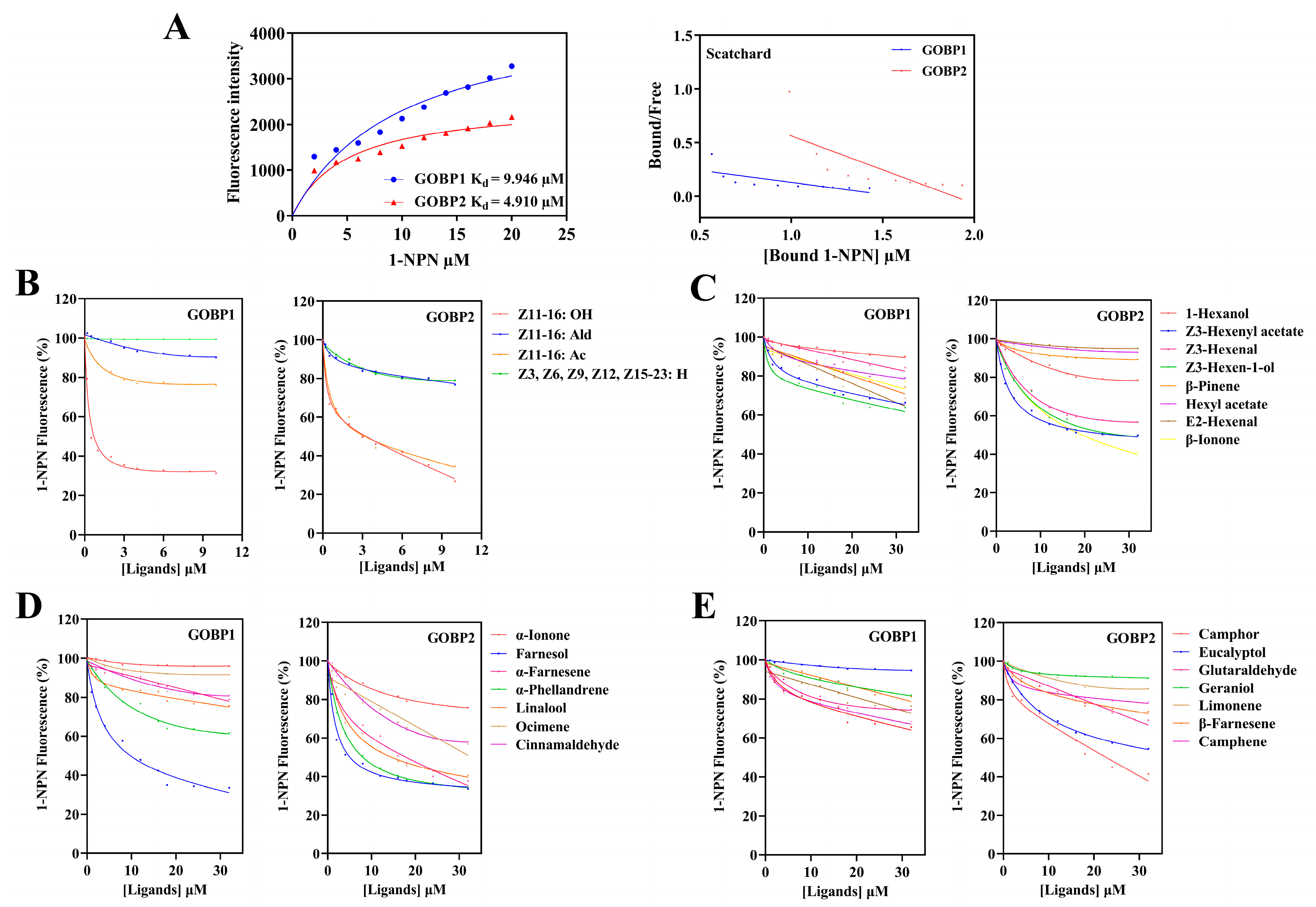
3.3. Fluorescence Binding Assays
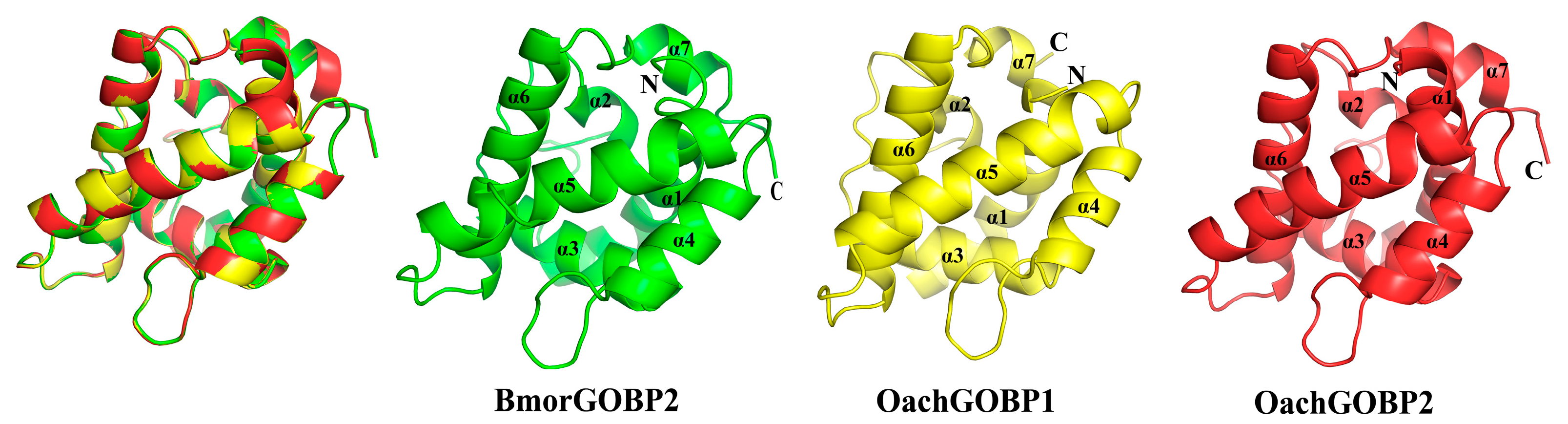
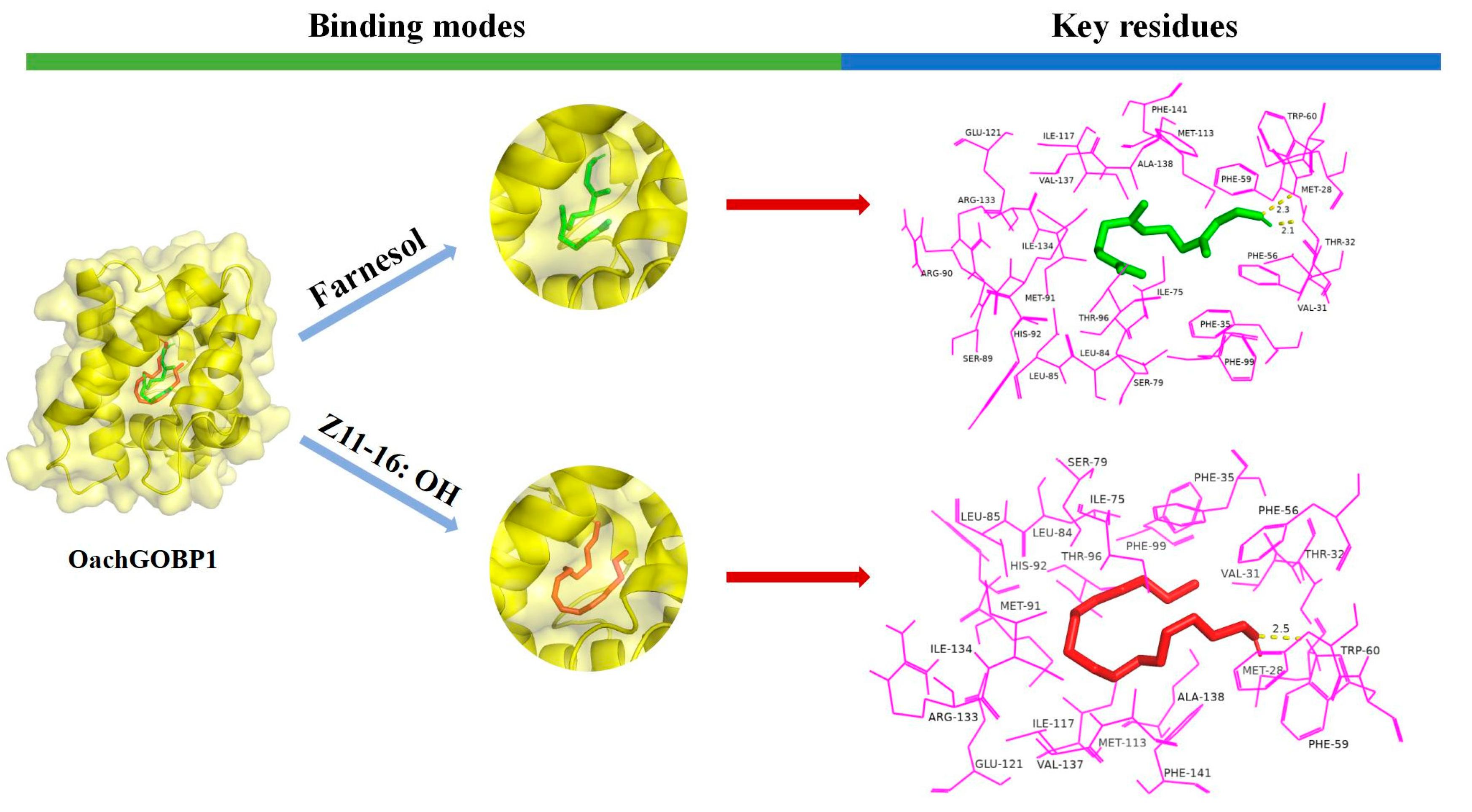
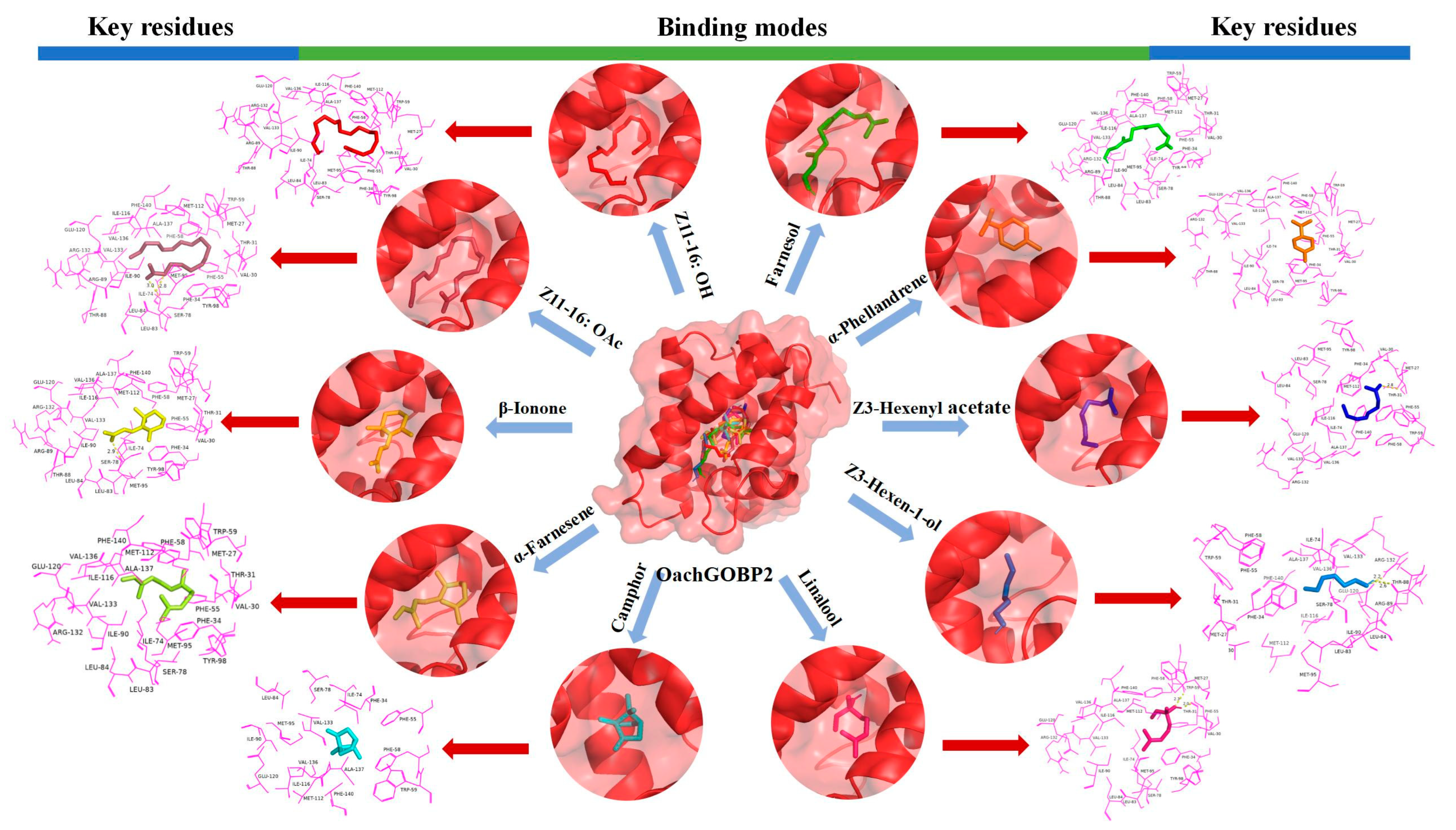
3.4. Protein Structure Homology Modeling and Molecular Docking
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gong, D.; Zhang, H.; Zhao, P.; Xia, Q.; Xiang, Z. The Odorant binding protein gene family from the genome of Silkworm, Bombyx mori. BMC Genom. 2009, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Brito, N.F.; Moreira, M.F.; Melo, A.C.A. A look inside odorant-binding proteins in insect chemoreception. J. Insect. Physiol. 2016, 95, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Blanchet, M.A.; Bains, G.; Pelosi, P.; Pevsner, J.; Snyder, S.H.; Monaco, H.L.; Amzel, L.M. The three-dimensional structure of bovine odorant binding protein and its mechanism of odor recognition. Nat. Struct. Biol. 1996, 3, 934–939. [Google Scholar] [CrossRef] [PubMed]
- Vogt, R.G.; Prestwich, G.D.; Lerner, M.R. Odorant-binding protein subfamililies associate with distinct classes of olfactory receptor neurons in insects. J. Neurobiol. 1991, 22, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Maida, R.; Proebstl, T.; Laue, M. Heterogeneity of odorant-binding proteins in the antennae of Bombyx mori. Chem. Senses 1997, 22, 503–515. [Google Scholar] [CrossRef]
- Liu, S.; Liu, N.; He, P.; Li, Z.; Dong, S.; Mu, L. Molecular characterization, expression patterns, and ligand-binding properties of two odorant-binding protein genes from Orthaga achatina (Butler) (Lepidoptera: Pyralidae). Arch. Insect Biochem. 2012, 80, 123–139. [Google Scholar] [CrossRef]
- Laue, M.; Steinbrecht, R.A.; Ziegelberger, G. Immunocytochemical localization of general odorant-binding protein in olfactory sensilla of the silkmoth Antheraea polyphemus. Naturwissenschaften 1994, 81, 178–180. [Google Scholar] [CrossRef]
- Huang, G.; Liu, J.; Zhou, J.; Wang, Q.; Dong, J.; Zhang, Y.; Li, X.; Li, J.; Gu, S. Expressional and functional comparisons of two general odorant binding proteins in Agrotis ipsilon. Insect Biochem. Molec. 2018, 98, 34–47. [Google Scholar] [CrossRef]
- Khuhro, S.A.; Liao, H.; Dong, X.; Yu, Q.; Yan, Q.; Dong, S. Two general odorant binding proteins display high bindings to both host plant volatiles and sex pheromones in a pyralid moth Chilo suppressalis (Lepidoptera: Pyralidae). J. Asia-Pac. Entomol. 2017, 20, 521–528. [Google Scholar] [CrossRef]
- Sun, H.; Yin, J.; Feng, H.; Li, K.; Xi, J.; Cao, Y. Expression, purification and binding characteristics of general odorant binding protein I (GOBP1) from the meadow moth, Loxostege sticticalia (Linnaeus) (Lepidoptera: Pyralididae). Acta Entomol. Sin. 2011, 54, 381–389. [Google Scholar]
- Acin, P.; Carrascal, M.; Abian, J.; Guerrero, A.; Quero, C. Expression of differential antennal proteins in males and females of an important crop pest, Sesamia nonagrioides. Insect Biochem. Molec. 2009, 39, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Zhou, W.; Yu, H.; Mao, C.; Zhang, C.; Cheng, J.; Zhu, Z. Cloning, expression and functional analysis of a general odorant-binding protein 2 gene of the rice striped stem borer, Chilo suppressalis (Walker) (Lepidoptera: Pyralidae). Insect Mol. Biol. 2009, 18, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Ma, L.; Zhao, J.; Xue, Z.; Yan, X.; Hao, C. Electrophysiological and behavioral responses of Plutella xylostella (Lepidoptera: Plutellidae) to volatiles from a non-host plant, geranium, Pelargonium hortorum (Geraniaceae). J. Agr. Food Chem. 2022, 70, 5982–5992. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.Q.; Qiu, G.S.; Li, Y.Y.; Zhang, H.J.; Yan, W.T.; Yue, Q.; Sun, L.N. Molecular characterization and functional analysis of pheromone binding proteins and general odorant binding proteins from Carposina sasakii Matsumura (Lepidoptera: Carposinidae). Pest Manag. Sci. 2019, 75, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Yang, K.; Liu, Y.; Xu, W.; Anderson, A.; Dong, S. Two general-odorant binding proteins in Spodoptera litura are differentially tuned to sex pheromones and plant odorants. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 180, 23–31. [Google Scholar] [CrossRef]
- Zhang, X.; Yan, Q.; Li, L.; Xu, J.; Mang, D.; Zhang, Y. Different binding properties of two general-odorant binding proteins in Athetis lepigone with sex pheromones, host plant volatiles and insecticides. Pestic. Biochem. Phys. 2020, 164, 173–182. [Google Scholar] [CrossRef]
- Khuhro, S.A.; Liao, H.; Zhu, G.H.; Li, S.M.; Ye, Z.F.; Dong, S. Tissue distribution and functional characterization of odorant binding proteins in Chilo suppressalis (Lepidoptera: Pyralidae). J. Asia-Pac. Entomol. 2017, 20, 1104–1111. [Google Scholar] [CrossRef]
- Si, Y.; Guo, J.; Liao, H.; Li, Y.; Ma, Y.; Zhu, Y.; Wei, Z.; Dong, S.; Yan, Q. Functional differentiation of three pheromone binding proteins in Orthaga achatina using mixed-type sex pheromones. Pestic. Biochem. Phys. 2022, 184, 105097. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Lopez, R. Clustal W and clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Sun, M.; Liu, Y.; Walker, W.B.; Liu, C.; Lin, K.; Gu, S.; Zhang, Y.; Zhou, J.; Wang, G. Identification and characterization of pheromone receptors and interplay between receptors and pheromone binding proteins in the Diamondback Moth, Plutella xyllostella. PLoS ONE 2013, 8, e62098. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Robertson, G.; He, X.; Dufour, S.; Hooper, A.M.; Pickett, J.A.; Keep, N.H.; Field, L.M. Characterisation of Bombyx mori odorant-binding proteins reveals that a general odorant-binding protein discriminates between Sex Pheromone components. J. Mol. Biol. 2009, 389, 529–545. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Yang, F.; Yang, K.; He, P.; Niu, X.; Xu, W.; Anderson, A.; Dong, S. Two subclasses of odorant-binding proteins in Spodoptera exigua display structural conservation and functional divergence. Insect Mol. Biol. 2015, 24, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Li, H.; Chen, Y.; Ye, Z.; You, X.; Zhou, J.; Dong, S. Identification and field evaluation of the sex pheromone of Orthaga achatina (Lepidoptera: Pyralidae). J. Chem. Ecol. 2018, 44, 886–893. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. Software News and Update AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated Docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Li, G.; Chen, X.; Li, B.; Zhang, G.; Li, Y.; Wu, J. Binding properties of general odorant binding proteins from the Oriental Fruit Moth, Grapholita molesta (Busck) (Lepidoptera: Tortricidae). PLoS ONE 2016, 11, e0155096. [Google Scholar] [CrossRef]
- Liu, Z.; Vidal, D.M.; Syed, Z.; Ishida, Y.; Leal, W.S. Pheromone binding to general odorant-binding proteins from the navel orangeworm. J. Chem. Ecol. 2010, 36, 787–794. [Google Scholar] [CrossRef]
- Liu, H.; Duan, H.; Wang, Q.; Xiao, Y.; Wang, Q.; Xiao, Q.; Sun, L.; Zhang, Y. Key amino residues determining binding activities of the odorant binding protein AlucOBP22 to two host plant terpenoids of Apolygus lucorum. J. Agr. Food Chem. 2019, 67, 5949–5956. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Y.; Wang, Q.; Sun, L.; Zhang, Y. The Adelphocoris lineolatus OBP4: Support for evolutionary and functional divergence of a mirid pheromone-binding protein from that found in lepidopteran moths. Insect Sci. 2022, 29, 151–161. [Google Scholar] [CrossRef]
- Han, W.; Yang, Y.; Si, Y.; Wei, Z.; Liu, S.; Liu, X.; Yan, Q.; Dong, S. Involvement of GOBP2 in the perception of a sex pheromone component in both larval and adult Spodoptera litura revealed using CRISPR/Cas9 mutagenesis. Insect Biochem. Molec. 2022, 141, 103719. [Google Scholar] [CrossRef]
- Cheng, X.; Cai, L.; Zheng, L.; Qin, J.; Huang, Y.; You, M. Cloning, expression profiling and binding characterization of the OBP2 gene in the diamondback moth, Plutella xylostella (Lepidoptera Plutellidae). Acta Entomol. Sin. 2016, 59, 365–376. [Google Scholar] [CrossRef]
- Wen, M.; Li, E.; Li, J.; Chen, Q.; Zhou, H.; Zhang, S.; Li, K.; Ren, B.; Wang, Y.; Yin, J. Molecular characterization and key binding sites of sex pheromone-binding proteins from the Meadow Moth, Loxostege sticticalis. J. Agr. Food Chem. 2019, 67, 12685–12695. [Google Scholar] [CrossRef]
- Hasan, R.; Rony, M.N.H.; Ahmed, R. In silico characterization and structural modeling of bacterial metalloprotease of family M4. J. Genet. Eng. Biotechnol. 2021, 19, 25. [Google Scholar] [CrossRef]
- Tian, Z.; Li, Y.; Xing, Y.; Li, R.; Liu, J. Structural insights into two representative conformations of the complex formed by Grapholita molesta (Busck) pheromone binding protein 2 and Z-8-Dodecenyl Acetate. J. Agr. Food Chem. 2019, 67, 4425–4434. [Google Scholar] [CrossRef]
- Fu, X.; Zhang, Y.; Qiu, Y.; Song, X.; Wu, F.; Feng, Y.; Zhang, J.; Li, H. Physicochemical basis and comparison of two type II sex pheromone components binding with pheromone-binding protein 2 from Tea Geometrid, Ectropis obliqua. J. Agr. Food Chem. 2018, 66, 13084–13095. [Google Scholar] [CrossRef] [PubMed]
- Northey, T.; Venthur, H.; De Biasio, F.; Chauviac, F.X.; Keep, N.H. Crystal structures and binding dynamics of odorant-binding Protein 3 from two aphid species Megoura viciae and Nasonovia ribisnigri. Sci. Rep. 2016, 6, 24739. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Xu, J.; Cui, Z.; Dong, X.; Ye, Z.; Niu, D.; Huang, Y.; Dong, S. Functional characterization of SlitPBP3 in Spodoptera litura by CRISPR/Cas9 mediated genome editing. Insect Biochem. Molec. 2016, 75, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Ban, L.; Song, L.; Liu, Y.; Pelosi, P.; Wang, G. General odorant-binding proteins and sex pheromone guide larvae of Plutella xylostella to better food. Insect Biochem. Molec. 2016, 72, 10–19. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Odor Compounds | GOBP1 | GOBP2 | ||||
|---|---|---|---|---|---|---|
| IC50 (μM) | Ki (μM) | Free Binding Energy (kcal·mol−1) | IC50 (μM) | Ki (μM) | Free Binding Energy (kcal·mol−1) | |
| Volatile compounds | ||||||
| 1-Hexanol | - | - | N | - | - | N |
| Z3-Hexenyl acetate | - | - | N | 16.77 | 14.30 | −5.4 |
| Z3-Hexenal | - | - | N | - | - | N |
| Z3-Hexen-1-ol | - | - | N | 21.36 | 18.18 | −4.7 |
| β-Pinene | - | - | N | - | - | N |
| Hexyl acetate | - | - | N | - | - | N |
| E2-Hexenal | - | - | N | - | - | N |
| β-Ionone | - | - | N | 17.76 | 15.51 | −8.0 |
| α-Ionone | - | - | N | - | - | N |
| Farnesol | 10.90 | 9.49 | −7.5 | 8.68 | 7.33 | −7.8 |
| α-Farnesene | - | - | N | 15.91 | 13.42 | −7.7 |
| α-Phellandrene | - | - | N | 10.21 | 8.71 | −7.2 |
| Linalool | - | - | N | 13.89 | 12.05 | −6.1 |
| Ocimene | - | - | N | - | - | N |
| Cinnamaldehyde | - | - | N | - | - | N |
| Camphor | - | - | N | 20.72 | 17.49 | −6.6 |
| Eucalyptol | - | - | N | - | - | N |
| Glutaraldehyde | - | - | N | - | - | N |
| Geraniol | - | - | N | - | - | N |
| Limonene | - | - | N | - | - | N |
| β-Farnesene | - | - | N | - | - | N |
| Camphene | - | - | N | - | - | N |
| Pheromones compounds | ||||||
| Z11-16: OH | 1.79 | 1.57 | −6.5 | 3.86 | 3.30 | −6.7 |
| Z11-16: OAc | - | - | N | 3.30 | 2.84 | −7.3 |
| Z11-16: Ald | - | - | N | - | - | N |
| Z3, Z6, Z9,Z12,Z15–23: H | - | - | N | - | - | N |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Li, Y.; Wei, Z.-Q.; Hou, J.-H.; Si, Y.-X.; Zhang, J.; Dong, S.-L.; Yan, Q. Identification and Functional Characterization of General Odorant Binding Proteins in Orthaga achatina. Insects 2023, 14, 216. https://doi.org/10.3390/insects14030216
Ma Y, Li Y, Wei Z-Q, Hou J-H, Si Y-X, Zhang J, Dong S-L, Yan Q. Identification and Functional Characterization of General Odorant Binding Proteins in Orthaga achatina. Insects. 2023; 14(3):216. https://doi.org/10.3390/insects14030216
Chicago/Turabian StyleMa, Yu, Yu Li, Zhi-Qiang Wei, Jing-Hao Hou, Yu-Xiao Si, Jin Zhang, Shuang-Lin Dong, and Qi Yan. 2023. "Identification and Functional Characterization of General Odorant Binding Proteins in Orthaga achatina" Insects 14, no. 3: 216. https://doi.org/10.3390/insects14030216
APA StyleMa, Y., Li, Y., Wei, Z.-Q., Hou, J.-H., Si, Y.-X., Zhang, J., Dong, S.-L., & Yan, Q. (2023). Identification and Functional Characterization of General Odorant Binding Proteins in Orthaga achatina. Insects, 14(3), 216. https://doi.org/10.3390/insects14030216