
The Multifaceted Effects of Short-Term Acute Hypoxia Stress: Insights into the Tolerance Mechanism of Propsilocerus akamusi (Diptera: Chironomidae)


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals, Hypoxia Challenge, and Tissue Sample Preparation
2.2. HE Staining
2.3. Tissue Enzyme Activity and Glycogen Assays
2.4. Transcriptome Analysis
2.5. Metabolome Analysis
2.6. The Integrated Analysis of Transcriptomic and Metabolomic Data
2.7. Data Analysis
3. Results
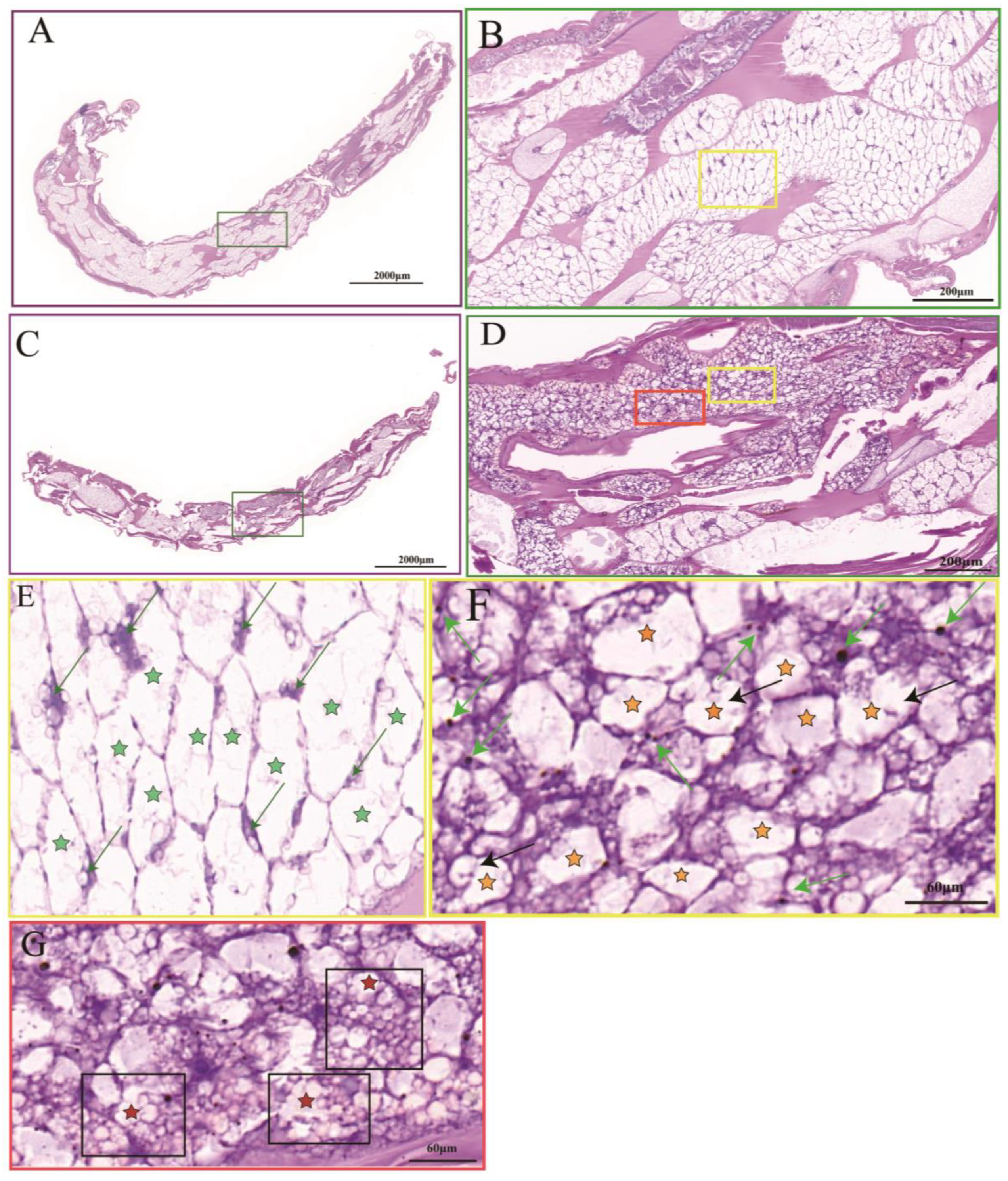
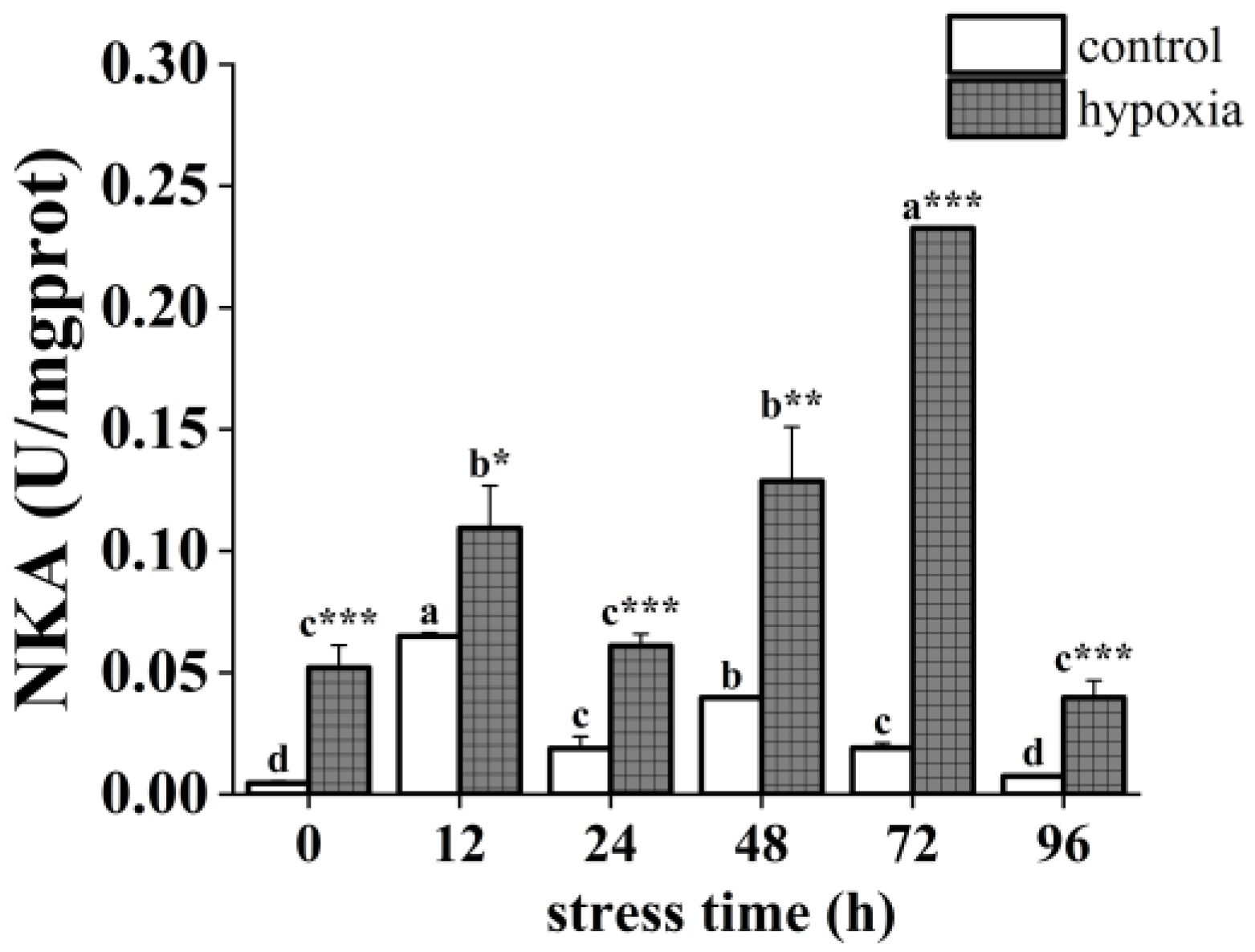
3.1. Effect of Hypoxic Stress on Histomorphological Feature and NKA Activity in P. akamusi
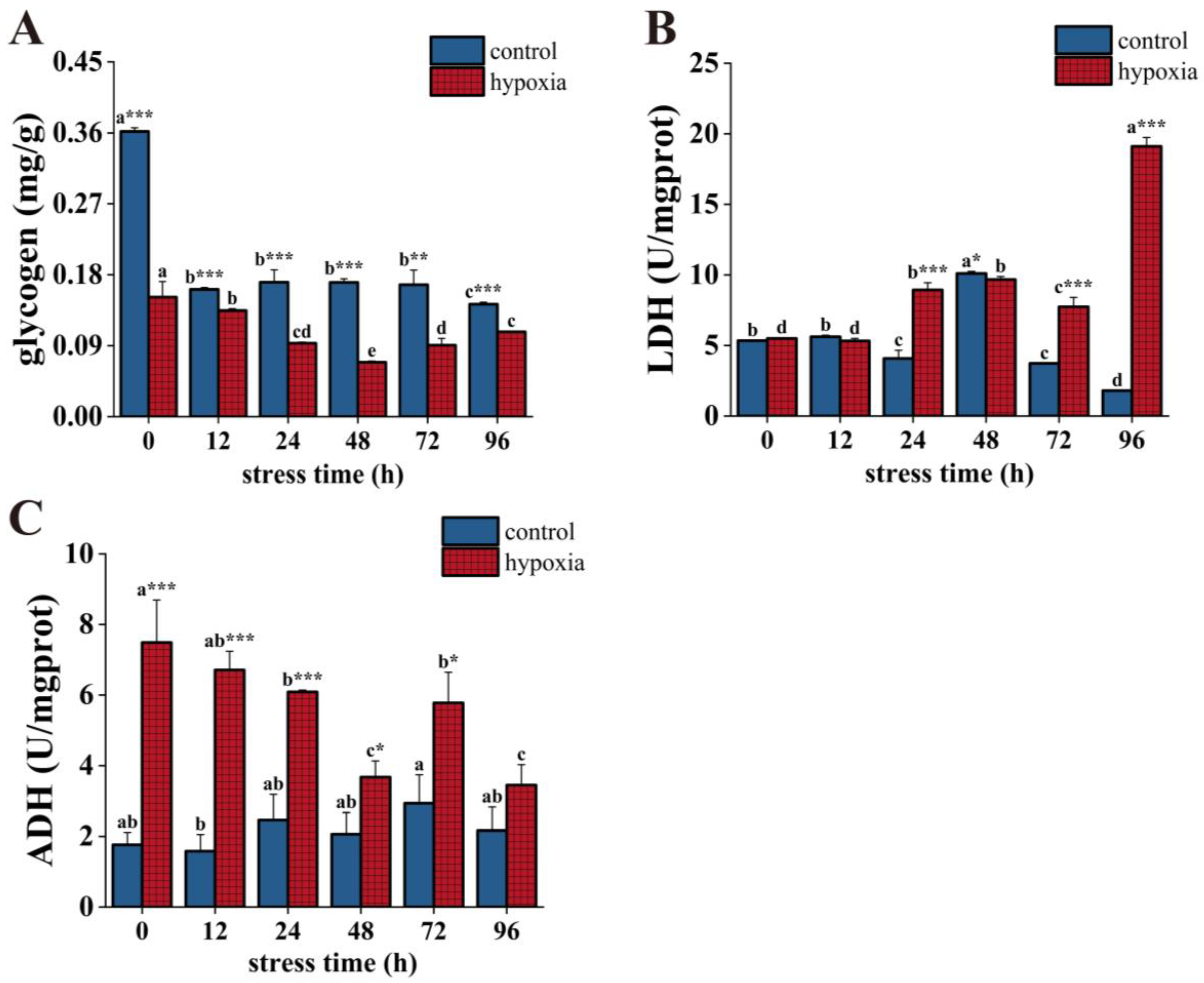
3.2. Effect of Hypoxic Stress on Energy Metabolism in P. akamusi
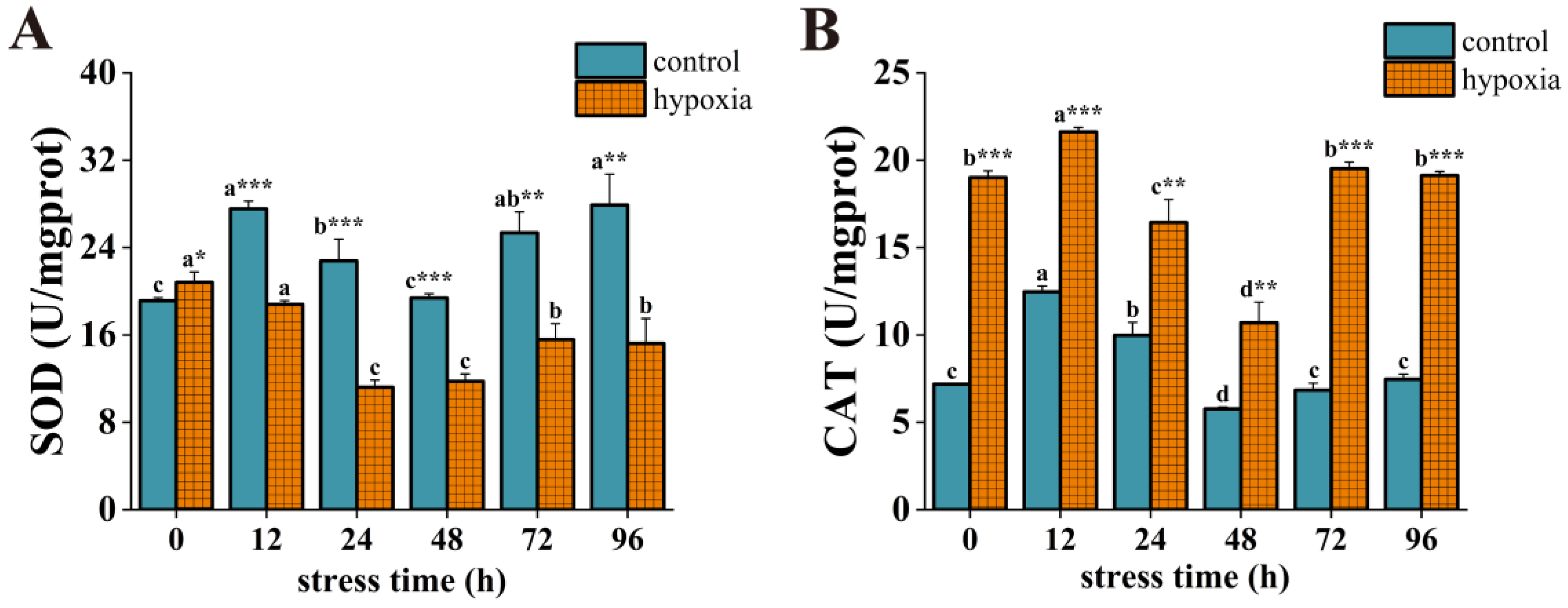
3.3. Effect of Hypoxic Stress on Antioxidant Enzyme Activity in P. akamusi
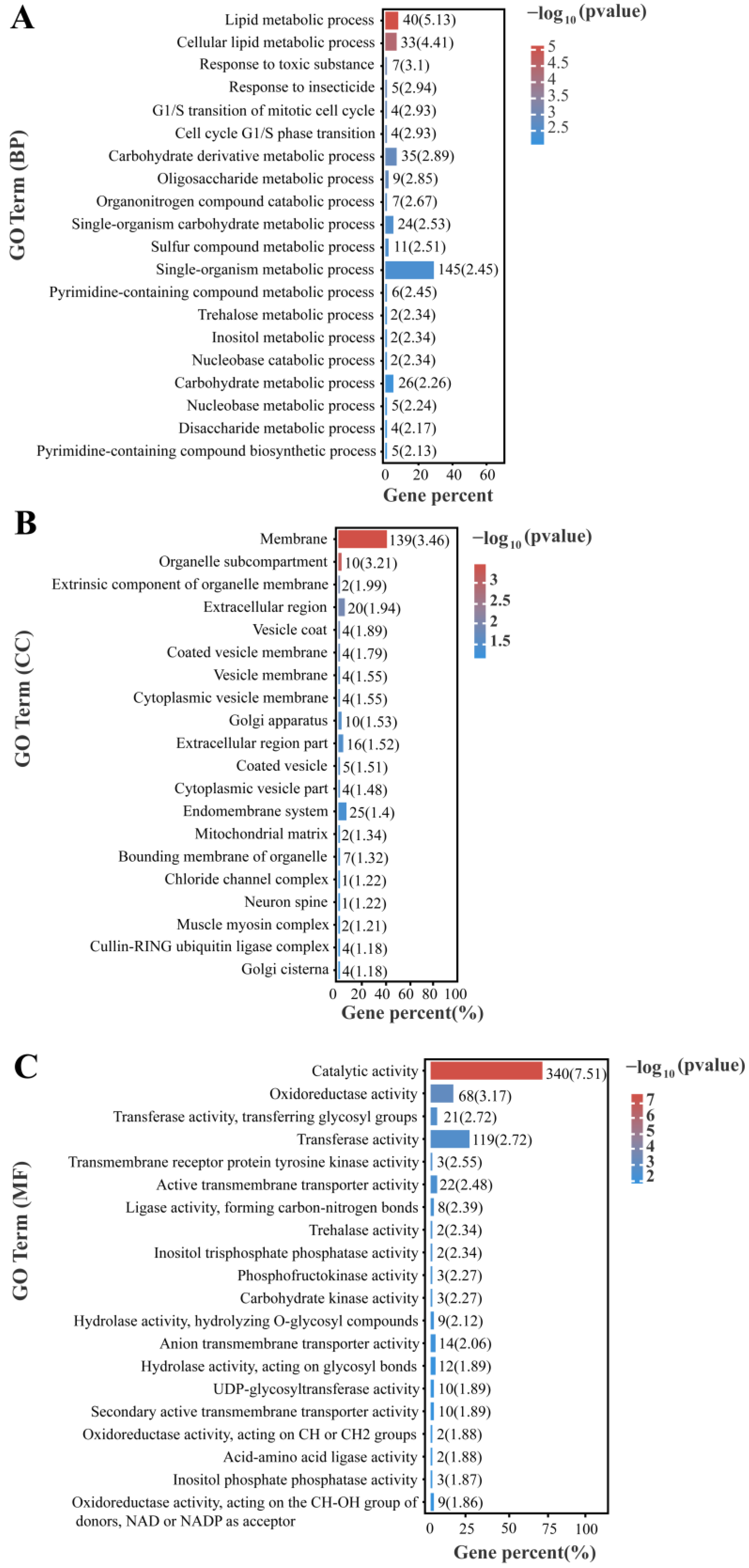
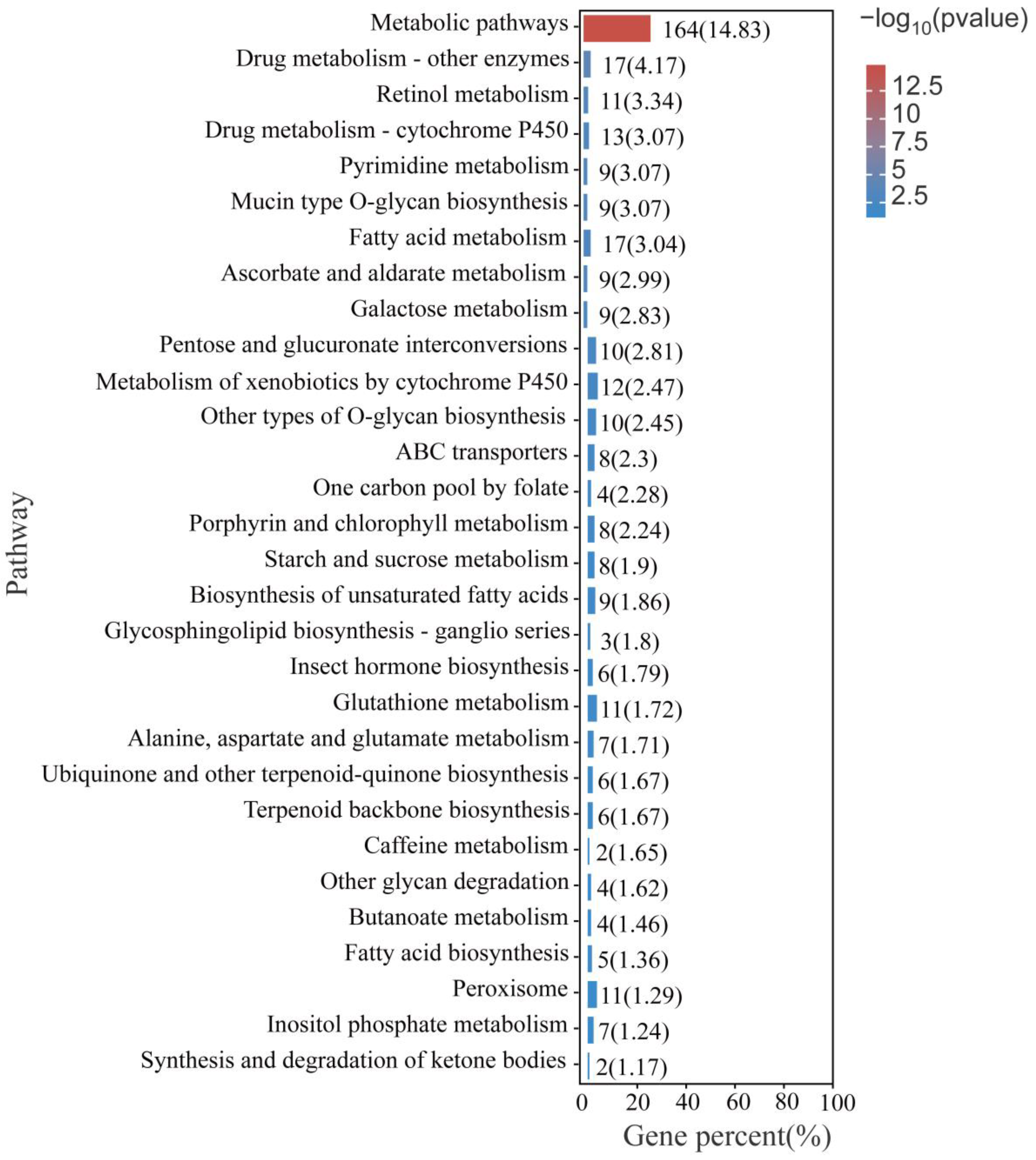
3.4. GO Enrichment and KEGG Enrichment Analysis of All Differentially Expressed Genes
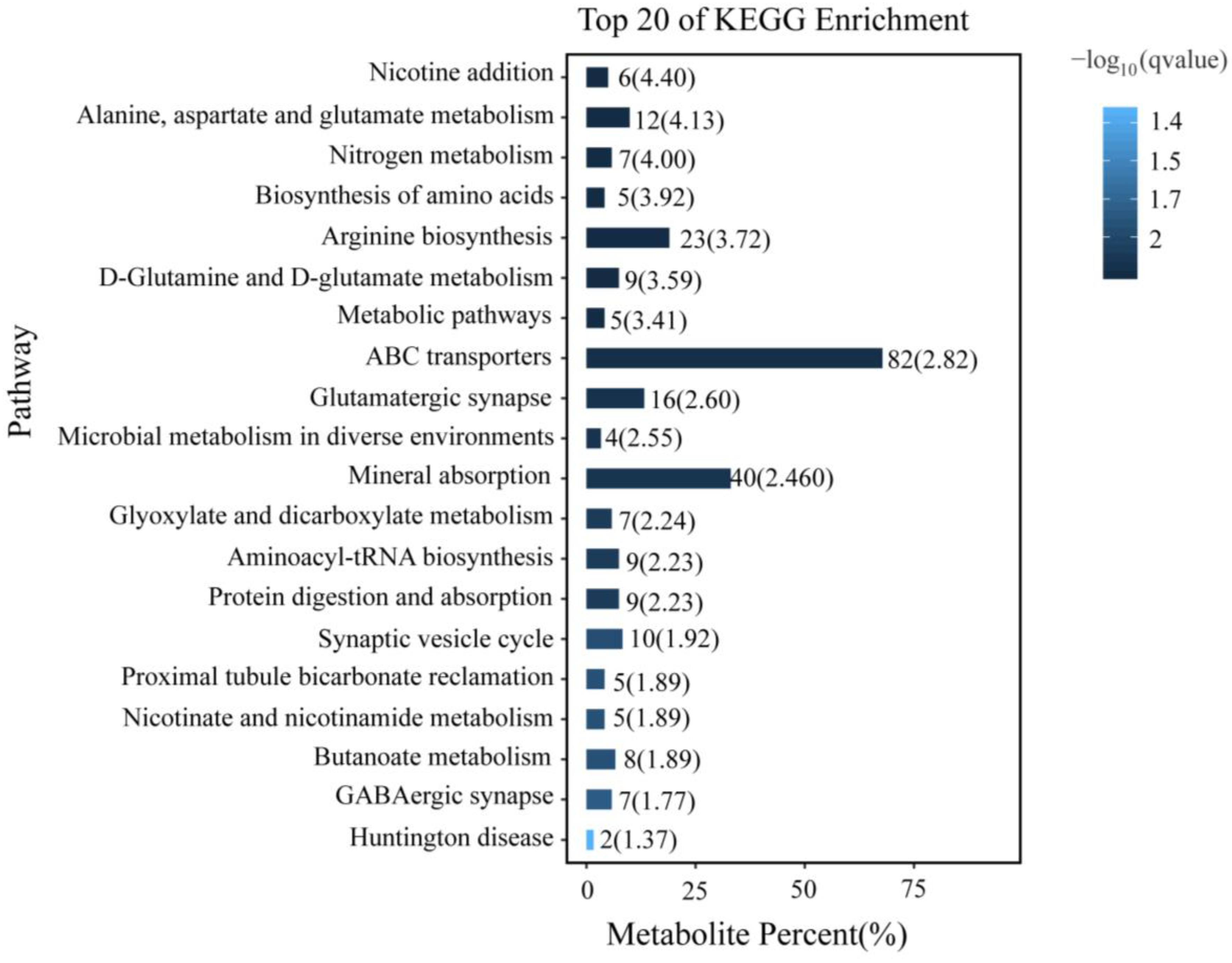
3.5. KEGG Enrichment Analysis of Differentially Accumulated Metabolites
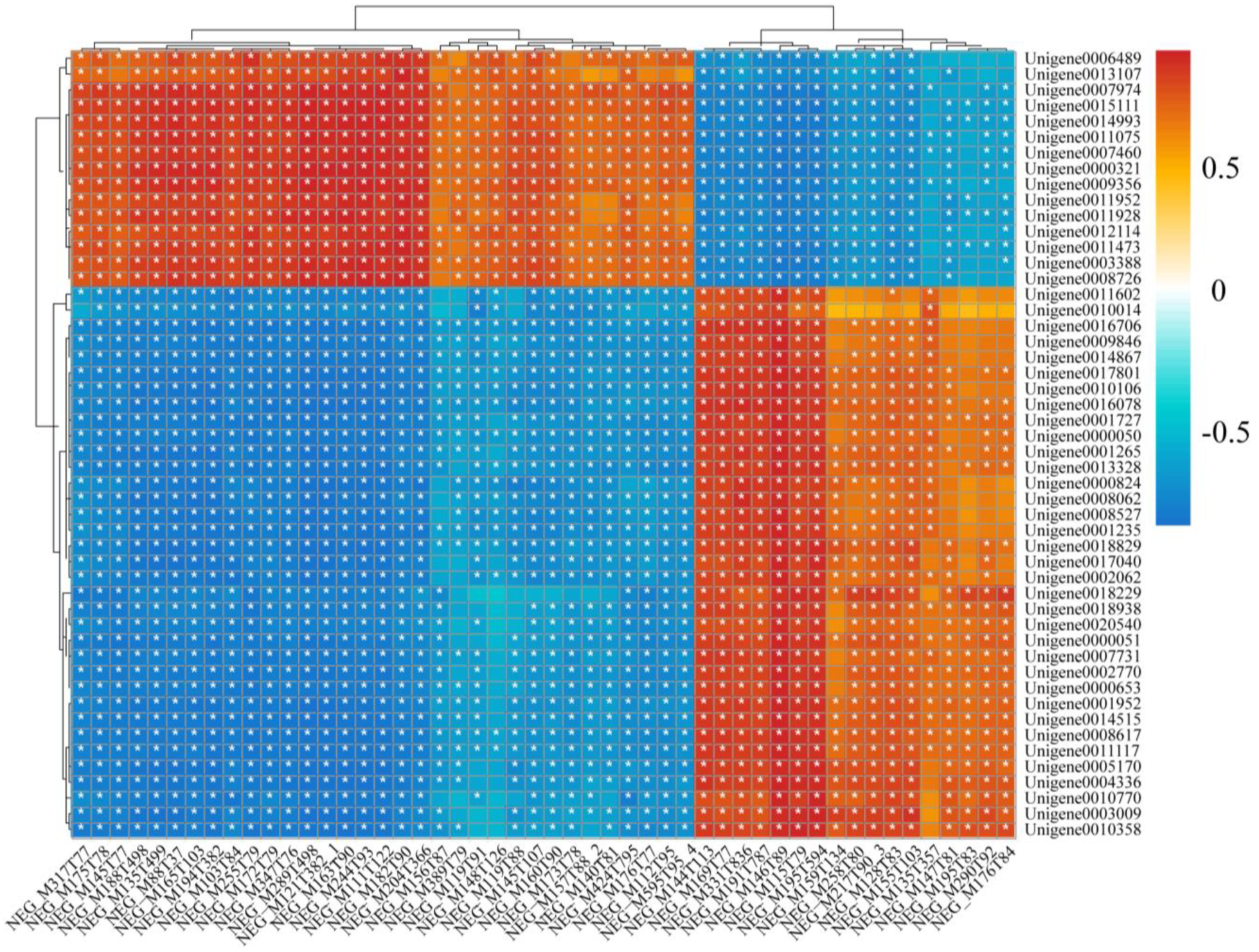
3.6. The Integrative Analysis of Transcriptome and Metabolome for P. akamusi in Response to Acute Hypoxia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jenny, J.P.; Francus, P.; Normandeau, A.; Lapointe, F.; Perga, M.E.; Ojala, A.; Schimmelmann, A.; Zolitschka, B. Global spread of hypoxia in freshwater ecosystems during the last three centuries is caused by rising local human pressure. Glob. Change Biol. 2016, 22, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Jenny, J.P.; Arnaud, F.; Alric, B.; Dorioz, J.M.; Sabatier, P.; Meybeck, M.; Perga, M.E. Inherited hypoxia: A new challenge for reoligotrophicated lakes under global warming. Glob. Biogeochem. Cycles 2014, 28, 1413–1423. [Google Scholar] [CrossRef]
- Wang, J.F.; Chen, J.G.; Ding, S.M.; Guo, J.Y.; Christopher, D.; Dai, Z.H.; Yang, H.Q. Effects of seasonal hypoxia on the release of phosphorus from sediments in deep-water ecosystem: A case study in Hongfeng Reservoir, Southwest China. Environ. Pollut. 2016, 219, 858–865. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.D.; Fu, H.; Liao, H.L.; Chen, H.H.; Liu, Z.W. Geochemical records of Lake Erhai (South-Western China) reveal the anthropogenically-induced intensification of hypolimnetic anoxia in monomictic lakes. Environ. Pollut. 2022, 299, 1873–6424. [Google Scholar] [CrossRef] [PubMed]
- Pardo, I.; García, L. Water abstraction in small lowland streams: Unforeseen hypoxia and anoxia effects. Sci. Total Environ. 2016, 568, 226–235. [Google Scholar] [CrossRef]
- Galic, N.; Hawkins, T.; Forbes, V.E. Adverse impacts of hypoxia on aquatic invertebrates: A meta-analysis. Sci. Total Environ. 2019, 652, 736–743. [Google Scholar] [CrossRef]
- Saari, G.N.; Wang, Z.; Brooks, B.W. Revisiting inland hypoxia: Diverse exceedances of dissolved oxygen thresholds for freshwater aquatic life. Environ. Sci. Pollut. Res. 2018, 25, 3139–3150. [Google Scholar] [CrossRef] [PubMed]
- Verberk, W.C.E.P.; Sommer, U.; Davidson, R.L.; Viant, M.R. Anaerobic Metabolism at Thermal Extremes: A Metabolomic Test of the Oxygen Limitation Hypothesis in an Aquatic Insect. Integr. Comp. Biol. 2013, 53, 609–619. [Google Scholar] [CrossRef]
- Cui, S.; Wang, L.; Qiu, J.; Liu, Z.; Geng, X. Comparative metabolomics analysis of Callosobruchus chinensis larvae under hypoxia, hypoxia/hypercapnia and normoxia. Pest. Manag. Sci. 2017, 73, 1267–1276. [Google Scholar] [CrossRef]
- Campbell, J.B.; Werkhoven, S.; Harrison, J.F. Metabolomics of anoxia tolerance in Drosophila melanogaster: Evidence against substrate limitation and for roles of protective metabolites and paralytic hypometabolism. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 317, R442–R450. [Google Scholar] [CrossRef]
- Correia, A.D.; Costa, M.H.; Luis, O.J.; Livingstone, D.R. Age-related changes in antioxidant enzyme activities, fatty acid composition and lipid peroxidation in whole body Gammarus locusta (Crustacea: Amphipoda). J. Exp. Mar. Biol. Ecol. 2003, 289, 83–101. [Google Scholar] [CrossRef]
- Via, J.D.; Thillart, G.V.d.; Cattani, O.; Cortesi, P. Behavioural responses and biochemical correlates in Solea solea to gradual hypoxic exposure. Can. J. Zool. 1998, 76, 2108–2113. [Google Scholar] [CrossRef]
- Lavarías, S.; Ocon, C.; van Oosterom, V.L.; Laino, A.; Medesani, D.A.; Fassiano, A.; Garda, H.; Donadelli, J.; de Molina, M.R.; Capítulo, A.R. Multibiomarker responses in aquatic insect Belostoma elegans (Hemiptera) to organic pollution in freshwater system. Environ. Sci. Pollut. Res. 2017, 24, 1322–1337. [Google Scholar] [CrossRef] [PubMed]
- Redecker, B.; Zebe, E. Anaerobic metabolism in aquatic insect larvae: Studies on Chironomus thummi and Culex pipiens. J. Comp. Physiol. B 1988, 158, 307–315. [Google Scholar] [CrossRef]
- Zebe, E. In vivo-studies on the function of hemoglobin in the larvae of Chironomus thummi (Insecta, Diptera). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 1991, 99, 525–529. [Google Scholar] [CrossRef]
- Englisch, H.; Opalka, B.; Zebe, E. The anaerobic metabolism of the larvae of the midge Chaoborus crystallinus. Insect Biochem. 1982, 12, 149–155. [Google Scholar] [CrossRef]
- Sun, S.; Wu, Y.; Fu, H.; Yang, M.; Ge, X.; Zhu, J.; Xuan, F.; Wu, X. Evaluating expression of autophagy-related genes in oriental river prawn Macrobrachium nipponense as potential biomarkers for hypoxia exposure. Ecotoxicol. Environ. Saf. 2019, 171, 484–492. [Google Scholar] [CrossRef]
- Andreyeva, A.Y.; Kladchenko, E.S.; Vyalova, O.Y.; Kukhareva, T.A. Functional Characterization of the Pacific Oyster, Crassostrea gigas (Bivalvia: Ostreidae), Hemocytes Under Normoxia and Short-Term Hypoxia. Turk. J. Fish. Aquat. Sci. 2021, 21, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Geihs, M.A.; Vargas, M.A.; Maia Nery, L.E. Damage caused during hypoxia and reoxygenation in the locomotor muscle of the crab Neohelice granulata (Decapoda: Varunidae). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2014, 172, 1–9. [Google Scholar] [CrossRef][Green Version]
- Felix-Portillo, M.; Martínez-Quintana, J.A.; Arenas-Padilla, M.; Mata-Haro, V.; Gómez-Jiménez, S.; Yepiz-Plascencia, G. Hypoxia drives apoptosis independently of p53 and metallothionein transcript levels in hemocytes of the whiteleg shrimp Litopenaeus vannamei. Chemosphere 2016, 161, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhong, P.; Yi, J.-Q.; Xu, A.-X.; Lin, W.-Y.; Guo, Z.-C.; Wang, C.-G.; Sun, C.-B.; Chan, S. Potential role for microRNA in facilitating physiological adaptation to hypoxia in the Pacific whiteleg shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 84, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, S.; Liu, T.; Chen, M.; Li, W.; Zhang, X. The transcriptomic responses of the ark shell, Anadara broughtonii, to sulfide and hypoxia exposure. Mol. Biol. Rep. 2019, 46, 4245–4257. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Liu, H.; Zhao, S.; Yu, S.; Xie, S.; Hua, S.; Yan, B.; Xing, C.; Gao, H. Hypoxia stress affects the physiological responses, apoptosis and innate immunity of Kuruma shrimp, Marsupenaeus japonicus. Fish Shellfish Immunol. 2022, 122, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Feng, J.; Song, H.; Zhou, C.; Yu, Z.-L.; Yang, M.-J.; Shi, P.; Guo, Y.-J.; Li, Y.-R.; Zhang, T. Mechanisms of heat and hypoxia defense in hard clam: Insights from transcriptome analysis. Aquaculture 2022, 549, 737792. [Google Scholar] [CrossRef]
- Ivanina, A.V.; Nesmelova, I.; Leamy, L.; Sokolov, E.P.; Sokolova, I.M. Intermittent hypoxia leads to functional reorganization of mitochondria and affects cellular bioenergetics in marine molluscs. J. Exp. Biol. 2016, 219, 1659–1674. [Google Scholar] [CrossRef] [PubMed]
- Lucu, Č. Hypoxia attenuate ionic transport in the isolated gill epithelium of Carcinus maenas. J. Comp. Physiol. B 2020, 190, 391–401. [Google Scholar] [CrossRef]
- Frank, C. Ecology, production and anaerobic metabolism of Chironomus plumosus L. larvae in a shallow lake. Arch. Hydrobiol. 1983, 96, 354–362. [Google Scholar]
- Hamburger, K.; Lindegaard, C.; Dall, P. Energy metabolism of Chironomus anthracinus (Chironomidae, Diptera) from the profundal zone of Lake Esrom, Denmark, as a function of body size, temperature and oxygen concentration. Hydrobiologia 1994, 294, 43–50. [Google Scholar] [CrossRef]
- Sun, S.M.; Guo, Z.B.; Fu, H.T.; Zhu, J.; Ge, X.P. Integrated metabolomic and transcriptomic analysis of brain energy metabolism in the male Oriental river prawn (Macrobrachium nipponense) in response to hypoxia and reoxygenation. Environ. Pollut. 2018, 243, 1154–1165. [Google Scholar] [CrossRef]
- Choi, J. Biomarkers in Environmental Monitoring and Its Application in Chironomus Spp. In Ecological Issues in a Changing World; Springer: Dordrecht, The Netherlands, 2004; pp. 203–215. [Google Scholar]
- Lencioni, V.; Bernabo, P.; Vanin, S. Respiration rate and oxy-regulatory capacity in cold stenothermal chironomids. J. Insect Physiol. 2008, 54, 1337–1342. [Google Scholar] [CrossRef]
- Rosenberg, D.M. Freshwater biomonitoring and Chironomidae. Neth. J. Aquat. Ecol. 1992, 26, 101–122. [Google Scholar] [CrossRef]
- Nath, B.B. Extracellular hemoglobin and environmental stress tolerance in Chironomus larvae. J. Limnol. 2018, 77, 112–122. [Google Scholar] [CrossRef]
- Hirabayashi, K.; Yoshizawa, K.; Yoshida, N.; Kazama, F. Progress of eutrophication and change of chironomid fauna in Lake Yamanakako, Japan. Limnology 2004, 5, 47–53. [Google Scholar] [CrossRef]
- Gong, Z.; Li, Y.; Xie, P. Seasonal changes, life cycle, and production of a psychrophilic chironomid (Propsilocerus akamusi) in a Chinese Lake. J. Freshwater Ecol. 2008, 23, 467–469. [Google Scholar] [CrossRef]
- Zou, W.; Cai, Y.; Tolonen, K.T.; Zhu, G.; Qin, B.; Peng, K.; Gong, Z. The adaptations to tube-dwelling life of Propsilocerus akamusi (Diptera: Chironomidae) larvae and its eutrophication-tolerant mechanisms. Limnologica 2019, 77, 125684. [Google Scholar] [CrossRef]
- Liu, W.; Chang, T.; Zhao, K.; Sun, X.; Qiao, H.; Yan, C.; Wang, Y. Genome-wide annotation of cuticular protein genes in non-biting midge Propsilocerus akamusi and transcriptome analysis of their response to heavy metal pollution. Int. J. Biol. Macromol. 2022, 223, 555–566. [Google Scholar] [CrossRef]
- Liu, W.; Sun, X.; Sun, W.; Zhou, A.; Li, R.; Wang, B.; Li, X.; Yan, C. Genome-wide analyses of ATP-Binding Cassette (ABC) transporter gene family and its expression profile related to deltamethrin tolerance in non-biting midge Propsilocerus akamusi. Aquat. Toxicol. 2021, 239, 105940. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Liu, Y.; Xu, H.; Yan, C. Genome-Wide Identification of P450 Genes in Chironomid Propsilocerus akamusi Reveals Candidate Genes Involved in Gut Microbiota-Mediated Detoxification of Chlorpyrifos. Insects 2022, 13, 765. [Google Scholar] [CrossRef]
- Yan, C.; Sun, X.; Cao, W.; Li, R.; Zhao, C.; Sun, Z.; Liu, W.; Pan, L. Identification and expression pattern of chemosensory genes in the transcriptome of Propsilocerus akamusi. PeerJ 2020, 8, 2167–8359. [Google Scholar] [CrossRef]
- Zheng, X.; Xie, Z.; Wang, S.; Lin, P. Determination of the protein expression profiles of Propsilocerus akamusi (Tokunaga) Malpighian tubules response to cadmium stress by iTRAQ coupled LC-MS/MS. J. Proteom. 2017, 164, 85–93. [Google Scholar] [CrossRef]
- Manzoni, C.; Kia, D.A.; Vandrovcova, J.; Hardy, J.; Wood, N.W.; Lewis, P.A.; Ferrari, R. Genome, transcriptome and proteome: The rise of omics data and their integration in biomedical sciences. Brief. Bioinf. 2018, 19, 286–302. [Google Scholar] [CrossRef]
- Hasin, Y.; Seldin, M.; Lusis, A. Multi-omics approaches to disease. Genome Biol. 2017, 18, 83. [Google Scholar] [CrossRef]
- Zou, W.; Cai, Y.; Tolonen, K.T.; Zhu, G.; Qin, B.; Peng, K.; Gong, Z. Vertical sediment migrations of dominant midge species in subtropical lakes with implications for bioassessment. Ecol. Indic. 2018, 95, 711–719. [Google Scholar] [CrossRef]
- Ma, X.; Ge, Q.; Taha, R.H.; Chen, K.; Yuan, Y. Beauveria bassiana Ribotoxin (BbRib) Induces Silkworm Cell Apoptosis via Activating Ros Stress Response. Processes 2021, 9, 1470. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Want, E.J.; O’Maille, G.; Smith, C.A.; Brandon, T.R.; Uritboonthai, W.; Qin, C.; Trauger, S.A.; Siuzdak, G. Solvent-dependent metabolite distribution, clustering, and protein extraction for serum profiling with mass spectrometry. Anal. Chem. 2006, 78, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef]
- Wenger, R.H. Mammalian oxygen sensing, signalling and gene regulation. J. Exp. Biol. 2000, 203, 1253–1263. [Google Scholar] [CrossRef]
- Wenger, R.H.; Stiehl, D.P.; Camenisch, G. Integration of oxygen signaling at the consensus HRE. Sci. STKE 2005, 2005, re12. [Google Scholar] [CrossRef] [PubMed]
- Doonan, F.; Cotter, T.G. Morphological assessment of apoptosis. Methods 2008, 44, 200–204. [Google Scholar] [CrossRef]
- Nuñez-Hernandez, D.M.; Felix-Portillo, M.; Peregrino-Uriarte, A.B.; Yepiz-Plascencia, G. Cell cycle regulation and apoptosis mediated by p53 in response to hypoxia in hepatopancreas of the white shrimp Litopenaeus vannamei. Chemosphere 2018, 190, 253–259. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X. The critical roles of mitochondrial alternative chains in juvenile ark shells (Anadara broughtonii) exposed to acute hypoxia with or without sulfide. Aquat. Toxicol. 2021, 241, 105996. [Google Scholar] [CrossRef] [PubMed]
- Chkadua, G.; Nozadze, E.; Tsakadze, L.; Shioshvili, L.; Arutinova, N.; Leladze, M.; Dzneladze, S.; Javakhishvili, M. Effect of H2O2 on Na, K-ATPase. J. Bioenerg. Biomembr. 2022, 54, 241–249. [Google Scholar] [CrossRef]
- Pratscher, B.; Friedrich, C.; Goger, W.; Allen, M.; Fink, D.; Thallinger, C.; Wolschek, M.; Frei, K.; Schöfer, C.; Pehamberger, H. Characterization of NKIP: A novel, Na+/K+-ATPase interacting protein mediates neural differentiation and apoptosis. Exp. Cell Res. 2008, 314, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Li, X.; Xing, Y.; Feng, C.; Jiang, H. Effects of hypoxia on immune responses and carbohydrate metabolism in the Chinese mitten crab, Eriocheir sinensis. Aquacult. Res. 2020, 51, 2735–2744. [Google Scholar] [CrossRef]
- Ryabova, A.; Cornette, R.; Cherkasov, A.; Watanabe, M.; Okuda, T.; Shagimardanova, E.; Kikawada, T.; Gusev, O. Combined metabolome and transcriptome analysis reveals key components of complete desiccation tolerance in an anhydrobiotic insect. Proc. Natl. Acad. Sci. USA 2020, 117, 19209–19220. [Google Scholar] [CrossRef] [PubMed]
- Padilla-Guerrero, I.E.; Barelli, L.; Gonzalez-Hernandez, G.A.; Torres-Guzman, J.C.; Bidochka, M.J. Flexible metabolism in Metarhizium anisopliae and Beauveria bassiana: Role of the glyoxylate cycle during insect pathogenesis. Microbiology 2011, 157, 199–208. [Google Scholar] [CrossRef][Green Version]
- Popov, V.N.; Moskalev, E.A.; Shevchenko, M.U.; Eprintsev, A.T. Comparative analysis of glyoxylate cycle key enzyme isocitrate lyase from organisms of different systematic groups. J. Evol. Biochem. Physiol. 2005, 41, 631–639. [Google Scholar] [CrossRef]
- Choi, J.; Roche, H.; Caquet, T. Hypoxia, hyperoxia and exposure to potassium dichromate or fenitrothion alter the energy metabolism in Chironomus riparius Mg.(Diptera: Chironomidae) larvae. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 130, 11–17. [Google Scholar] [CrossRef]
- Jing, H.; Liu, Z.; Wu, B.; Tu, K.; Liu, Z.; Sun, X.; Zhou, L. Physiological and Molecular Responses to Hypoxia Stress in Manila Clam Ruditapes Philippinarum. Aquat. Toxicol. 2023, 257, 106428. [Google Scholar] [CrossRef]
- Cook, D.M.; Doran-Peterson, J. Mining diversity of the natural biorefinery housed within Tipula abdominalis larvae for use in an industrial biorefinery for production of lignocellulosic ethanol. Insect Sci. 2010, 17, 303–312. [Google Scholar] [CrossRef]
- Giordano, F.J. Oxygen, oxidative stress, hypoxia, and heart failure. J. Clin. Investig. 2005, 115, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, J.; Imamura, M.; Fujita, M. Oxidative stress response of caddisfly Stenopsyche marmorata larvae to abrupt hypoxia-normoxia shift. Aquat. Toxicol. 2019, 211, 66–72. [Google Scholar] [CrossRef]
- Welker, A.F.; Moreira, D.C.; Campos, É.G.; Hermes-Lima, M. Role of redox metabolism for adaptation of aquatic animals to drastic changes in oxygen availability. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2013, 165, 384–404. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, U.O.; da Rosa Araújo, A.S.; Belló-Klein, A.; da Silva, R.S.M.; Kucharski, L.C. Effects of environmental anoxia and different periods of reoxygenation on oxidative balance in gills of the estuarine crab Chasmagnathus granulata. Comp. Biochem. Physiol. Part. B Biochem. Mol. Biol. 2005, 140, 51–57. [Google Scholar] [CrossRef]
- Hou, L.; Zhu, L.; Kong, X.; Lu, Y.; Wang, W.; Gu, W.; Wang, L.; Meng, Q. Proteomic analysis of Eriocheir sinensis hemocytes in response to hypoxia stress. Aquac. Rep. 2021, 21, 100876. [Google Scholar] [CrossRef]
- García-Triana, A.; Zenteno-Savín, T.; Peregrino-Uriarte, A.B.; Yepiz-Plascencia, G. Hypoxia, reoxygenation and cytosolic manganese superoxide dismutase (cMnSOD) silencing in Litopenaeus vannamei: Effects on cMnSOD transcripts, superoxide dismutase activity and superoxide anion production capacity. Dev. Comp. Immunol. 2010, 34, 1230–1235. [Google Scholar] [CrossRef]
- Trasviña-Arenas, C.H.; Garcia-Triana, A.; Peregrino-Uriarte, A.B.; Yepiz-Plascencia, G. White shrimp Litopenaeus vannamei catalase: Gene structure, expression and activity under hypoxia and reoxygenation. Comp. Biochem. Physiol. Part. B Biochem. Mol. Biol. 2013, 164, 44–52. [Google Scholar] [CrossRef]
- Asai, K.; Buurman, W.A.; Reutelingsperger, C.P.M.; Schutte, B.; Kaminishi, M. Low concentrations of ethanol induce apoptosis in human intestinal cells. Scand. J. Gastroenterol. 2003, 38, 1154–1161. [Google Scholar] [CrossRef]
- Chen, P.; Hu, Y.-F.; Wang, L.; Xiao, W.-F.; Bao, X.-Y.; Pan, C.; Yi, H.-S.; Chen, X.-Y.; Pan, M.-H.; Lu, C. Mitochondrial apoptotic pathway is activated by H2O2-mediated oxidative stress in BmN-SWU1 cells from Bombyx mori ovary. PLoS ONE 2015, 10, e0134694. [Google Scholar] [CrossRef]
- Hasnain, S.E.; Taneja, T.K.; Sah, N.K.; Mohan, M.; Pathak, N.; Sahdev, S.; Athar, M.; Totey, S.M.; Begum, R. In vitro cultured Spodoptera frugiperda insect cells: Model for oxidative stress-induced apoptosis. J. BioSci. 1999, 24, 13–19. [Google Scholar] [CrossRef]
- Slomiany, B.L.; Piotrowski, J.; Slomiany, A. Chronic Alcohol Ingestion Enhances Tumor Necrosis Factor-α Expression and Salivary Gland Apoptosis. Alcohol. Clin. Exp. Res. 1997, 21, 1530–1533. [Google Scholar] [CrossRef]
- Souza, H.S.; Elia, C.C.; Braulio, V.B.; Côrtes, M.Q.; Furtado, V.C.; Garrofé, H.C.; Martinusso, C.A. Effects of ethanol on gut-associated lymphoid tissues in a model of bacterial translocation: A possible role of apoptosis. Alcohol 2003, 30, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Young, B.A.; Norris, R.H.; Sheldon, F. Is the hyporheic zone a refuge for macroinvertebrates in drying perennial streams? Mar. Freshw. Res. 2011, 62, 1373–1382. [Google Scholar] [CrossRef]
- Meskar, A.; Plee-Gautier, E.; Amet, Y.; Berthou, F.; Lucas, D. Alcohol-xenobiotic interactions: Role of cytochrome P450 2E1. Pathol. Biol. 2001, 49, 696–702. [Google Scholar] [CrossRef]
- Watts, R.J.; Kopf, R.K.; McCasker, N.; Howitt, J.A.; Conallin, J.; Wooden, I.; Baumgartner, L. Adaptive Management of Environmental Flows: Using Irrigation Infrastructure to Deliver Environmental Benefits During a Large Hypoxic Blackwater Event in the Southern Murray–Darling Basin, Australia. Environ. Manag. 2018, 61, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Baoligao, B.; Xu, F.; Chen, X.; Wang, X.; Chen, W. Acute impacts of reservoir sediment flushing on fishes in the Yellow River. J. Hydro-Environ. Res. 2016, 13, 26–35. [Google Scholar] [CrossRef]
- Hladyz, S.; Watkins, S.C.; Whitworth, K.L.; Baldwin, D.S. Flows and hypoxic blackwater events in managed ephemeral river channels. J. Hydrol. 2011, 401, 117–125. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhang, Q.-J.; Xu, W.-B.; Zou, W.; Xiang, X.-L.; Gong, Z.-J.; Cai, Y.-J. The Multifaceted Effects of Short-Term Acute Hypoxia Stress: Insights into the Tolerance Mechanism of Propsilocerus akamusi (Diptera: Chironomidae). Insects 2023, 14, 800. https://doi.org/10.3390/insects14100800
Zhang Y, Zhang Q-J, Xu W-B, Zou W, Xiang X-L, Gong Z-J, Cai Y-J. The Multifaceted Effects of Short-Term Acute Hypoxia Stress: Insights into the Tolerance Mechanism of Propsilocerus akamusi (Diptera: Chironomidae). Insects. 2023; 14(10):800. https://doi.org/10.3390/insects14100800
Chicago/Turabian StyleZhang, Yao, Qing-Ji Zhang, Wen-Bin Xu, Wei Zou, Xian-Ling Xiang, Zhi-Jun Gong, and Yong-Jiu Cai. 2023. "The Multifaceted Effects of Short-Term Acute Hypoxia Stress: Insights into the Tolerance Mechanism of Propsilocerus akamusi (Diptera: Chironomidae)" Insects 14, no. 10: 800. https://doi.org/10.3390/insects14100800
APA StyleZhang, Y., Zhang, Q.-J., Xu, W.-B., Zou, W., Xiang, X.-L., Gong, Z.-J., & Cai, Y.-J. (2023). The Multifaceted Effects of Short-Term Acute Hypoxia Stress: Insights into the Tolerance Mechanism of Propsilocerus akamusi (Diptera: Chironomidae). Insects, 14(10), 800. https://doi.org/10.3390/insects14100800