
Effects of Virgin Coconut Oil-Enriched Diet on Immune and Antioxidant Enzymatic Activity, Fat and Vitellogenin Contents in Newly Emerged and Forager Bees (Apis mellifera L.) Reared in Cages

 ,
,  , , , ,
, , , ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Honey Bee Sampling and Rearing
2.2. Feed Intake and Survival Rate
2.3. Enzymatic Activity
2.4. Fat Content
2.5. Vitellogenin Content
2.6. Statistical Analysis
3. Results
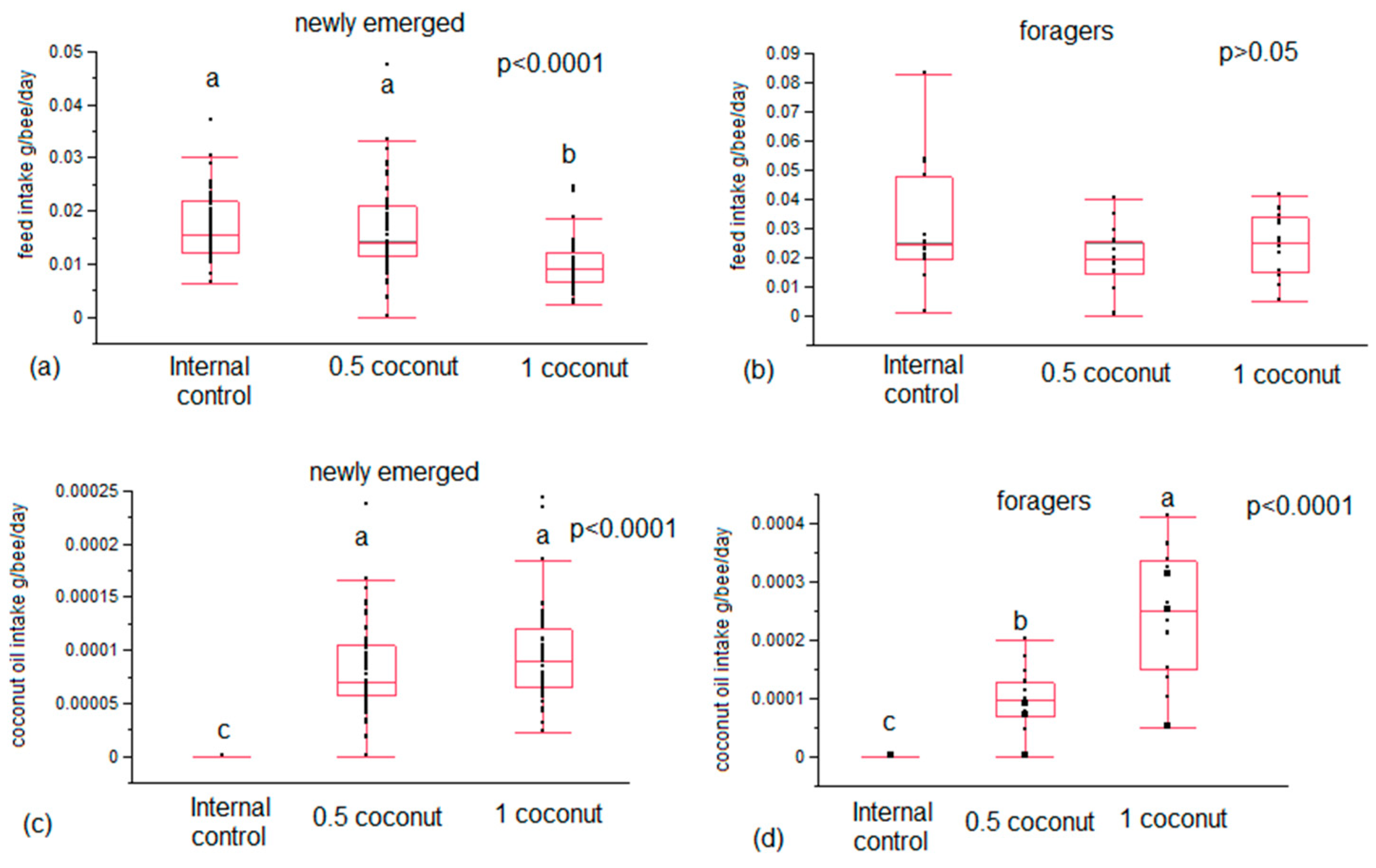
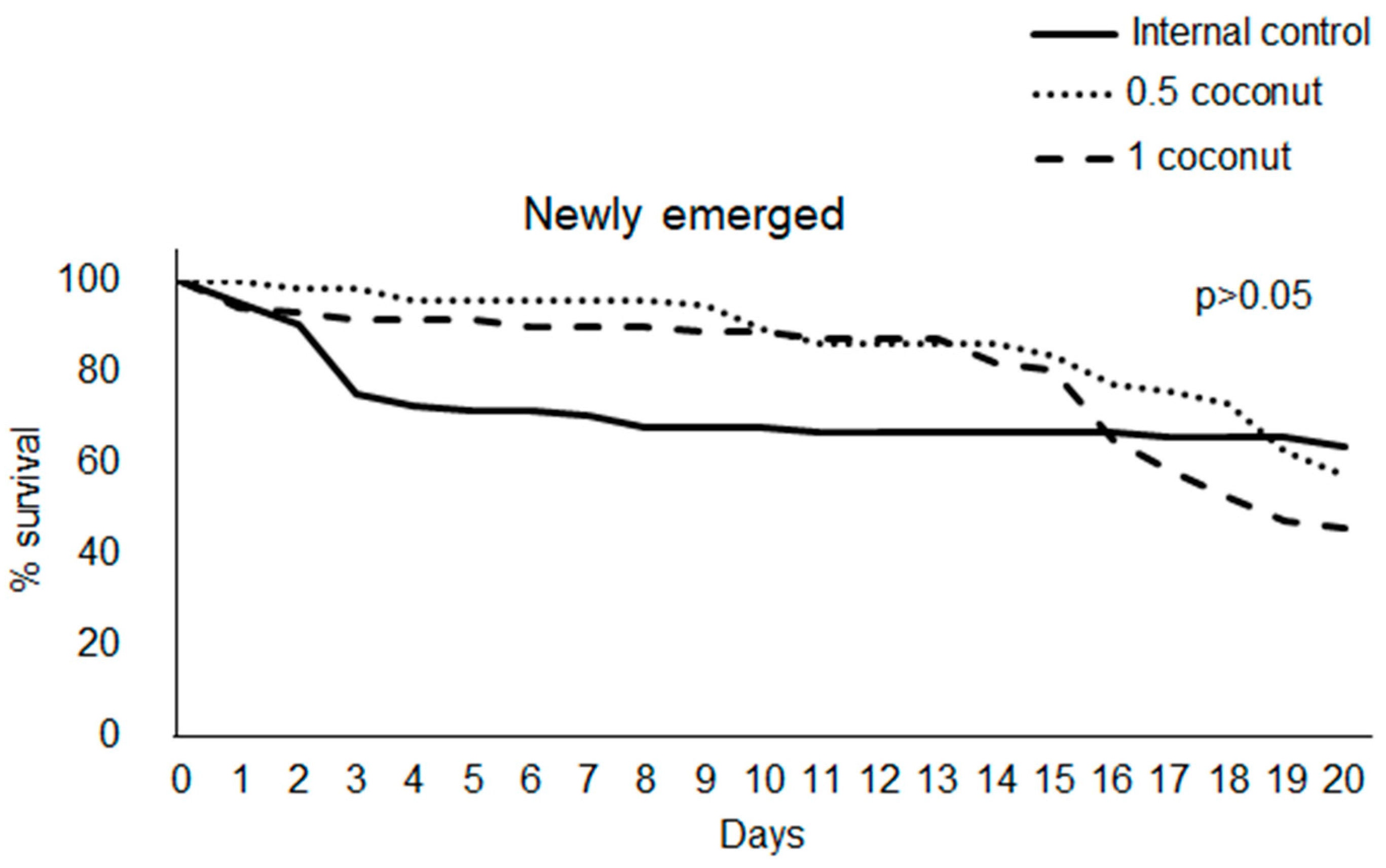
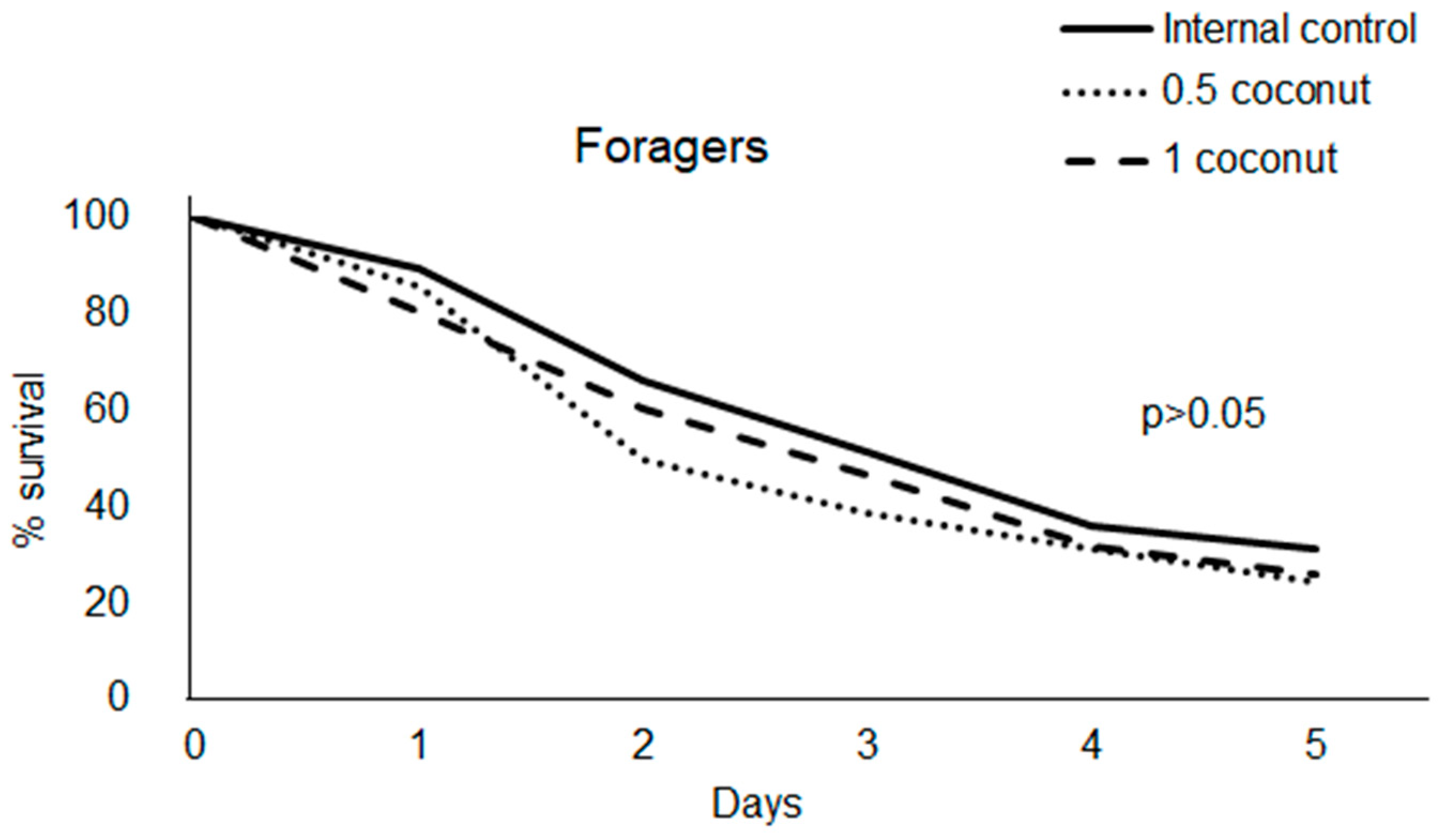
3.1. Feed Intake and Survival Rate
3.2. Enzymatic Activity
3.3. Fat and Vitellogenin Contents
3.4. Correlations
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Meixner, M.D. A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them. J. Invertebr. Pathol. 2010, 103, S80–S95. [Google Scholar] [CrossRef]
- Dolezal, A.G.; Toth, A.L. Feedbacks between nutrition and disease in honey bee health. Curr. Opin. Insect Sci. 2018, 26, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.D.; Aronstein, K.; Chen, Y.P.; Hetru, C.; Imler, J.L.; Jiang, H.; Kanost, M.; Thompson, G.J.; Zou, Z.; Hultmark, D. Immune pathways and defence mechanisms in honey bees Apis mellifera. Insect Mol. Biol. 2006, 15, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Wilson-Rich, N.; Dres, S.T.; Starks, P.T. The ontogeny of immunity: Development of innate immune strength in the honey bee (Apis mellifera). J. Insect Physiol. 2008, 54, 1392–1399. [Google Scholar] [CrossRef] [PubMed]
- Bucekova, M.; Valachova, I.; Kohutova, L.; Prochazka, E.; Klaudiny, J.; Majtan, J. Honeybee glucose oxidase-its expression in honeybee workers and comparative analyses of its content and H2O2-mediated antibacterial activity in natural honeys. Naturwissenschaften 2014, 101, 661–670. [Google Scholar] [CrossRef]
- Schmid-Hempel, P. Evolutionary ecology of insect immune defenses. Annu. Rev. Entomol. 2005, 50, 529–551. [Google Scholar] [CrossRef]
- González-Santoyo, I.; Córdoba-Aguilar, A. Phenoloxidase: A key component of the insect immune system. Entomol. Exp. Appl. 2012, 142, 1–16. [Google Scholar] [CrossRef]
- Amdam, G.V.; Simões, Z.L.; Hagen, A.; Norberg, K.; Schrøder, K.; Mikkelsen, Ø.; Kirkwood, T.B.L.; Omholt, S.W. Hormonal control of the yolk precursor vitellogenin regulates immune function and longevity in honeybees. Exp. Gerontol. 2004, 39, 767–773. [Google Scholar] [CrossRef]
- Salmela, H.; Sundström, L. Vitellogenin in inflammation and immunity in social insects. Inflamm. Cell Signal. 2018, 5, e1506. [Google Scholar] [CrossRef][Green Version]
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207. [Google Scholar] [CrossRef]
- Yu, S.J.; Robinson, F.A.; Nation, J.L. Detoxication capacity in the honey bee, Apis mellifera L. Pestic. Biochem. Physiol. 1984, 22, 360–368. [Google Scholar] [CrossRef]
- Weirich, G.F.; Collins, A.M.; Williams, V.P. Antioxidant enzymes in the honey bee, Apis mellifera. Apidologie 2002, 33, 3–14. [Google Scholar] [CrossRef]
- Sagona, S.; Betti, L.; Casini, L.; Palego, L.; Giannaccini, G.; Felicioli, A. Antioxidant enzymes activity during age polyethism in Apis mellifera L., 1758. J. Apic. Res. 2020, 60, 879–889. [Google Scholar] [CrossRef]
- Alaux, C.; Ducloz, F.; Crauser, D.; Le Conte, Y. Diet effects on honeybee immunocompetence. Biol. Lett. 2010, 6, 562–565. [Google Scholar] [CrossRef]
- DeGrandi-Hoffman, G.; Chen, Y.; Huang, E.; Huang, M.H. The effect of diet on protein concentration, hypopharyngeal gland development and virus load in worker honey bees (Apis mellifera L.). J. Insect Physiol. 2010, 56, 1184–1191. [Google Scholar] [CrossRef]
- Mazzei, M.; Fronte, B.; Sagona, S.; Carrozza, M.L.; Forzan, M.; Pizzurro, F.; Bibbiani, C.; Miragliotta, V.; Abramo, F.; Millanta, F.; et al. Effect of 1, 3-1, 6 β-glucan on natural and experimental deformed wing virus infection in newly emerged honeybees (Apis mellifera ligustica). PLoS ONE 2016, 11, e0166297. [Google Scholar] [CrossRef]
- Felicioli, A.; Forzan, M.; Sagona, S.; D’Agostino, P.; Baido, D.; Fronte, B.; Mazzei, M. Effect of oral administration of 1, 3-1, 6 β-glucans in DWV naturally infected newly emerged bees (Apis mellifera L.). Vet. Sci. 2020, 7, 52. [Google Scholar] [CrossRef]
- Brodschneider, R.; Haidmayer, C.; Riessberger-Gallé, U.; Crailsheim, K. Protein uptake in honey bee colonies supplemented with two protein diets simultaneously. Apidologie 2009, 40, 662. [Google Scholar]
- Pirk, C.W.; Boodhoo, C.; Human, H.; Nicolson, S.W. The importance of protein type and protein to carbohydrate ratio for survival and ovarian activation of caged honey bees (Apis mellifera scutellata). Apidologie 2010, 41, 62–72. [Google Scholar] [CrossRef]
- Szymaś, B.; Jędruszuk, A. The influence of different diets on haemocytes of adult worker honey bees, Apis mellifera. Apidologie 2003, 34, 97–102. [Google Scholar] [CrossRef]
- Zheng, B.; Wu, Z.; Xu, B. The effects of dietary protein levels on the population growth, performance, and physiology of honey bee workers during early spring. J. Insect Sci. 2014, 14, 191. [Google Scholar] [CrossRef] [PubMed]
- Lamontagne-Drolet, M.; Samson-Robert, O.; Giovenazzo, P.; Fournier, V. The impacts of two protein supplements on commercial honey bee (Apis mellifera L.) colonies. J. Apic. Res. 2019, 58, 800–813. [Google Scholar] [CrossRef]
- Camilli, M.P.; de Barros, D.C.; Justulin, L.A.; Tse, M.L.; Orsi, R.D.O. Protein feed stimulates the development of mandibular glands of honey bees (Apis mellifera). J. Apic. Res. 2021, 60, 165–171. [Google Scholar] [CrossRef]
- Sagona, S.; Coppola, F.; Nanetti, A.; Tafi, E.; Palego, L.; Betti, L.; Giannaccini, G.; Felicioli, A. Effects of Two Commercial Protein Diets on the Health of Two Imago Ages of Apis mellifera L. Reared in Laboratory. Animals 2022, 12, 968. [Google Scholar] [CrossRef] [PubMed]
- Sagona, S.; Minieri, S.; Coppola, F.; Gatta, D.; Casini, L.; Palego, L.; Betti, L.; Giannaccini, G.; Felicioli, A. Effects of chestnut hydrolysable tannin enrichment in the artificial diet of forager bees, Apis mellifera. J. Apic. Res. 2021, 1–7. [Google Scholar] [CrossRef]
- Farjan, M.; Dmitryjuk, M.; Lipiński, Z.; Biernat-Łopieńska, E.; Żółtowska, K. Supplementation of the honey bee diet with vitamin C: The effect on the antioxidative system of Apis mellifera carnica brood at different stages. J. Apic. Res. 2012, 51, 263–270. [Google Scholar] [CrossRef]
- Stabler, D.; Al-Esawy, M.; Chennells, J.A.; Perri, G.; Robinson, A.; Wright, G.A. Regulation of dietary intake of protein and lipid by nurse-age adult worker honeybees. J. Exp. Biol. 2021, 224, jeb230615. [Google Scholar] [CrossRef]
- Manning, R. Fatty acids in pollen: A review of their importance for honey bees. Bee World 2001, 82, 60–75. [Google Scholar] [CrossRef]
- Wang, X.; Zhong, Z.; Chen, X.; Hong, Z.; Lin, W.; Mu, X.; Hu, X.; Zheng, H. High-fat diets with differential fatty acids induce obesity and perturb gut microbiota in honey bee. Int. J. Mol. Sci. 2021, 22, 834. [Google Scholar] [CrossRef]
- Yu, J.; Zhang, W.; Chi, X.; Chen, W.; Li, Z.; Wang, Y.; Liu, Z.; Wang, H.; Xu, B. The dietary arachidonic acid improved growth and immunity of honey bee (Apis mellifera ligustica). Bull. Entomol. Res. 2022, 112, 261–270. [Google Scholar] [CrossRef]
- Manning, R.; Speijers, J.; Harvey, M.; Black, J. Added vegetable and fish oils to low-fat pollen diets: Effect on honey bee (Apis mellifera L.) consumption. Aust. J. Entomol. 2010, 49, 182–189. [Google Scholar] [CrossRef]
- Deen, A.; Visvanathan, R.; Wickramarachchi, D.; Marikkar, N.; Nammi, S.; Jayawardana, B.C.; Liyanage, R. Chemical composition and health benefits of coconut oil: An overview. J. Sci. Food Agric. 2021, 101, 2182–2193. [Google Scholar] [CrossRef] [PubMed]
- Haydak, M.H. Honey bee nutrition. Annu. Rev. Entomol. 1970, 15, 143–156. [Google Scholar] [CrossRef]
- Toth, A.L.; Kantarovich, S.; Meisel, A.F.; Robinson, G.E. Nutritional status influences socially regulated foraging ontogeny in honey bees. J. Exp. Biol. 2005, 208, 4641–4649. [Google Scholar] [CrossRef] [PubMed]
- Villarino, B.J.; Dy, L.M.; Lizada, M.C.C. Descriptive sensory evaluation of virgin coconut oil and refined, bleached and deodorized coconut oil. LWT Food Sci. Technol. 2007, 40, 193–199. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Ziegler, R.; Ibrahim, M.M. Formation of lipid reserves in fat body and eggs of the yellow fever mosquito, Aedes aegypti. J. Insect Physiol. 2002, 47, 623–627. [Google Scholar] [CrossRef]
- Dolezal, A.G.; Carrillo-Tripp, J.; Miller, W.A.; Bonning, B.C.; Toth, A.L. Intensively cultivated landscape and Varroa mite infestation are associated with reduced honeybee nutritional state. PLoS ONE 2016, 11, e0153531. [Google Scholar] [CrossRef]
- SAS Institute. JMP Statistics and Graphics Guide; SAS Institute Inc.: Cary, NC, USA, 2008. [Google Scholar]
- Schmid, M.R.; Brockmann, A.; Pirk, C.W.; Stanley, D.W.; Tautz, J. Adult honeybees (Apis mellifera L.) abandon hemocytic, but not phenoloxidase-based immunity. J. Insect Physiol. 2008, 54, 439–444. [Google Scholar] [CrossRef]
- Laughton, A.M.; Boots, M.; Siva-Jothy, M.T. The ontogeny of immunity in the honey bee, Apis mellifera L. following an immune challenge. J. Insect Physiol. 2011, 57, 1023–1032. [Google Scholar] [CrossRef] [PubMed]
- Chan, Q.W.; Mutti, N.S.; Foster, L.J.; Kocher, S.D.; Amdam, G.V.; Wolschin, F. The worker honeybee fat body proteome is extensively remodeled preceding a major life-history transition. PLoS ONE 2011, 6, e24794. [Google Scholar] [CrossRef] [PubMed]
- Toth, A.L.; Robinson, G.E. Worker nutrition and division of labour in honeybees. Anim. Behav. 2005, 69, 427–435. [Google Scholar] [CrossRef]
- Bitondi, M.M.G.; Simoes, Z.P. The relationship between level of pollen in the diet, vitellogenin and juvenile hormone titres in Africanized Apis mellifera workers. J. Apic. Res. 1996, 35, 27–36. [Google Scholar] [CrossRef]
- Sagona, S.; Coppola, F.; Nanetti, A.; Cardaio, I.; Tafi, E.; Palego, L.; Betti, L.; Giannaccini, G.; Felicioli, A. Queen Caging and Oxalic Acid Treatment: Combined Effect on Vitellogenin Content and Enzyme Activities in the First Post-Treatment Workers and Drones, Apis mellifera L. Animals 2022, 12, 3121. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Newly Emerged | Glucose Oxidase Activity (U/µg of Protein) | Phenoloxidase Activity (mU/mg of Protein) | Glutathione S Transferase Activity (U/mg of Protein) | |
|---|---|---|---|---|
| T0 | 29.92 SD 25.11 (27.48) | 14.78 SD 12.11 (11.07) b | 24.94 SD 29.20 (15.30) | |
| T5 | External control (nurse bees) | 37.31 SD 12.37 (39.27) | 41.34 SD 15.85 (44.34) a | 32.34 SD 8.10 (29.05) |
| Internal control | 32.75 SD 14.36 (31.85) | 29.61 SD 11.49 (29.48) a | 52.34 SD 27.73 (49.55) | |
| 0.5% coconut | 37.70 SD 18.55 (38.10) | 31.11 SD 10.51 (31.28) a | 33.92 SD 28.05 (26.26) | |
| 1% coconut | 30.00 SD 9.40 (30.84) | 11.64 SD 8.33 (9.89) b | 20.66 SD 12.98 (17.70) | |
| T20 | External control (guardian bees) | 43.68 SD 9.96 (40.45) | 28.06 SD 19.14 (26.49) ab | 30.20 SD 20.63 (23.25) |
| Internal control | 27.42 SD 8.51 (29.28) | 42.66 SD 23.48 (36.06) a | 30.93 SD 20.98 (26.85) | |
| 0.5% coconut | 29.62 SD 11.28 (28.13) | 26.93 SD 18.65 (16.96) ab | 32.93 SD 20.25 (31.98) | |
| 1% coconut | 31.21 SD 9.14 (32.62) | 33.63 SD 44.42 (16.77) ab | 38.76 SD 22.52 (41.47) | |
| p value, χ2 test or F, DF = 8 | p = 0.5922, F = 0.8163 | p = 0.0085, χ2 = 20.5449 | p = 0.4315, χ2 = 8.0215 | |
| Foragers | Glucose Oxidase Activity (U/mg of Protein) | Phenoloxidase Activity (mU/mg of Protein) | Glutathione S Transferase Activity (U/mg of Protein) | |
|---|---|---|---|---|
| T0 (External control) | 48.33 SD 21.93 (54.94) | 41.38 SD 23.77 (37.83) | 38.55 SD 21.30 (29.88) | |
| T5 | Internal Control | 49.37 SD 15.22 (46.71) | 33.33 SD 24.37 (31.55) | 35.19 SD 16.40 (35.39) |
| 0.5% coconut | 47.93 SD 16.31 (47.76) | 22.49 SD 13.66 (20.11) | 19.71 SD 11.64 (21.45) | |
| 1% coconut | 30.81 SD 18.76 (26.03) | 33.03 SD 32.75 (23.57) | 28.96 SD 9.18 (28.89) | |
| p value, χ2 test, DF = 3 | p = 0.2650, F = 1.4250 | p = 0.5049, χ2 = 2.3400 | p = 0.2360, χ2 = 4.2467 | |
| Comparisons | Mean SD (Median) | p Value |
|---|---|---|
| external control T5 vs. external control T20 | 0.103 SD 0.036 (0.099) vs. 0.123 SD 0.023 (0.125) | p > 0.05 |
| internal control T5 vs. internal control T20 | 0.135 SD 0.028 (0.141) vs. 0.083 SD 0.030 (0.088) | p = 0.0163 |
| 0.5% coconut T5 vs. 0.5% coconut T20 | 0.150 SD 0.054 (0.137) vs. 0.123 SD 0.037 (0.127) | p > 0.05 |
| 1% coconut T5 vs. 1% coconut T20 | 0.184 SD 0.027 (0.177) vs. 0.163 SD 0.077 (0.134) | p > 0.05 |
| Time | Honeybees | Diet | Fat Content mg/bee | Feed Intake mg/bee |
|---|---|---|---|---|
| T5 | Newly emerged | Internal control | 5.050 SD 0.802 | 0.016 SD 0.004 |
| T5 | Newly emerged | 0.5% coconut | 6.267 SD 2.614 | 0.017 SD 0.003 |
| T5 | Newly emerged | 1% coconut | 5.383 SD 1.422 | 0.010 SD 0.002 |
| T20 | Newly emerged | Internal control | 3.550 SD 1.232 | 0.017 SD 0.006 |
| T20 | Newly emerged | 0.5% coconut | 6.033 SD 2.661 | 0.017 SD 0.008 |
| T20 | Newly emerged | 1% coconut | 7.467 SD 3.189 | 0.010 SD 0.005 |
| T5 | Foragers | Internal control | 4.650 SD 1.183 | 0.031 SD 0.02 |
| T5 | Foragers | 0.5% coconut | 5.600 SD 1.787 | 0.020 SD 0.011 |
| T5 | Foragers | 1% coconut | 7.017 SD 3.089 | 0.025 SD 0.011 |
| Comparisons | Mean SD (Median) | p Value |
|---|---|---|
| external control T5 vs. external control T20 | 0.067 SD 0.014 (0.060) vs. 0.240 SD 0.066 (0.215) | p = 0.0495 |
| internal control T5 vs. internal control T20 | 0.239 SD 0.048 (0.217) vs. 0.245 SD 0.098 (0.206) | p > 0.05 |
| 0.5% coconut T5 vs. 0.5% coconut T20 | 0.138 SD 0.017 (0.144) vs. 0.243 SD 0.076 (0.152) | p = 0.0495 |
| 1% coconut T5 vs. 1% coconut T20 | 0.154 SD 0.066 (0.142) vs. 0.114 SD 0.058 (0.137) | p > 0.05 |
| Survival Rate | GOX Activity | PO Activity | Glutathione S Transferase Activity | Fat Content | Vitellogenin Content | |
|---|---|---|---|---|---|---|
| Newly emerged T5 Coconut oil intake | r = 0.4426 p = 0.0659 | r = −0.06749 p = 0.7902 | r = −0.5737 p = 0.0128 | r = −0.3796 p = 0.1202 | r = 0.5779 p = 0.0120 | r = −0.3725 p = 0.1279 |
| Newly emerged T20 Coconut oil intake | r = 0.03482 p = 0.8909 | r = 0.1668 p = 0.5084 | r = −0.2232 p = 0.3733 | r = 0.07580 p = 0.7650 | r = 0.5263 p = 0.0249 | r = −0.5483 p = 0.0185 |
| Foragers T5 Coconut oil intake | r = 0.2611 p = 0.2953 | r = −0.09746 p = 0.7004 | r = −0.05848 p = 0.8177 | r = −0.1906 p = 0.4487 | r = 0.4239 p = 0.0796 | r = −0.2239 p = 0.3718 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sagona, S.; Coppola, F.; Tafi, E.; Orlando, C.; D’Onofrio, C.; Boni, C.B.; Casini, L.; Palego, L.; Betti, L.; Giannaccini, G.; et al. Effects of Virgin Coconut Oil-Enriched Diet on Immune and Antioxidant Enzymatic Activity, Fat and Vitellogenin Contents in Newly Emerged and Forager Bees (Apis mellifera L.) Reared in Cages. Insects 2023, 14, 856. https://doi.org/10.3390/insects14110856
Sagona S, Coppola F, Tafi E, Orlando C, D’Onofrio C, Boni CB, Casini L, Palego L, Betti L, Giannaccini G, et al. Effects of Virgin Coconut Oil-Enriched Diet on Immune and Antioxidant Enzymatic Activity, Fat and Vitellogenin Contents in Newly Emerged and Forager Bees (Apis mellifera L.) Reared in Cages. Insects. 2023; 14(11):856. https://doi.org/10.3390/insects14110856
Chicago/Turabian StyleSagona, Simona, Francesca Coppola, Elena Tafi, Caterina Orlando, Chiara D’Onofrio, Chiara Benedetta Boni, Lucia Casini, Lionella Palego, Laura Betti, Gino Giannaccini, and et al. 2023. "Effects of Virgin Coconut Oil-Enriched Diet on Immune and Antioxidant Enzymatic Activity, Fat and Vitellogenin Contents in Newly Emerged and Forager Bees (Apis mellifera L.) Reared in Cages" Insects 14, no. 11: 856. https://doi.org/10.3390/insects14110856
APA StyleSagona, S., Coppola, F., Tafi, E., Orlando, C., D’Onofrio, C., Boni, C. B., Casini, L., Palego, L., Betti, L., Giannaccini, G., & Felicioli, A. (2023). Effects of Virgin Coconut Oil-Enriched Diet on Immune and Antioxidant Enzymatic Activity, Fat and Vitellogenin Contents in Newly Emerged and Forager Bees (Apis mellifera L.) Reared in Cages. Insects, 14(11), 856. https://doi.org/10.3390/insects14110856