
Climatic Niche, Altitudinal Distribution, and Vegetation Type Preference of the Flea Beetle Genus Arsipoda in New Caledonia (Coleoptera Chrysomelidae)




Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
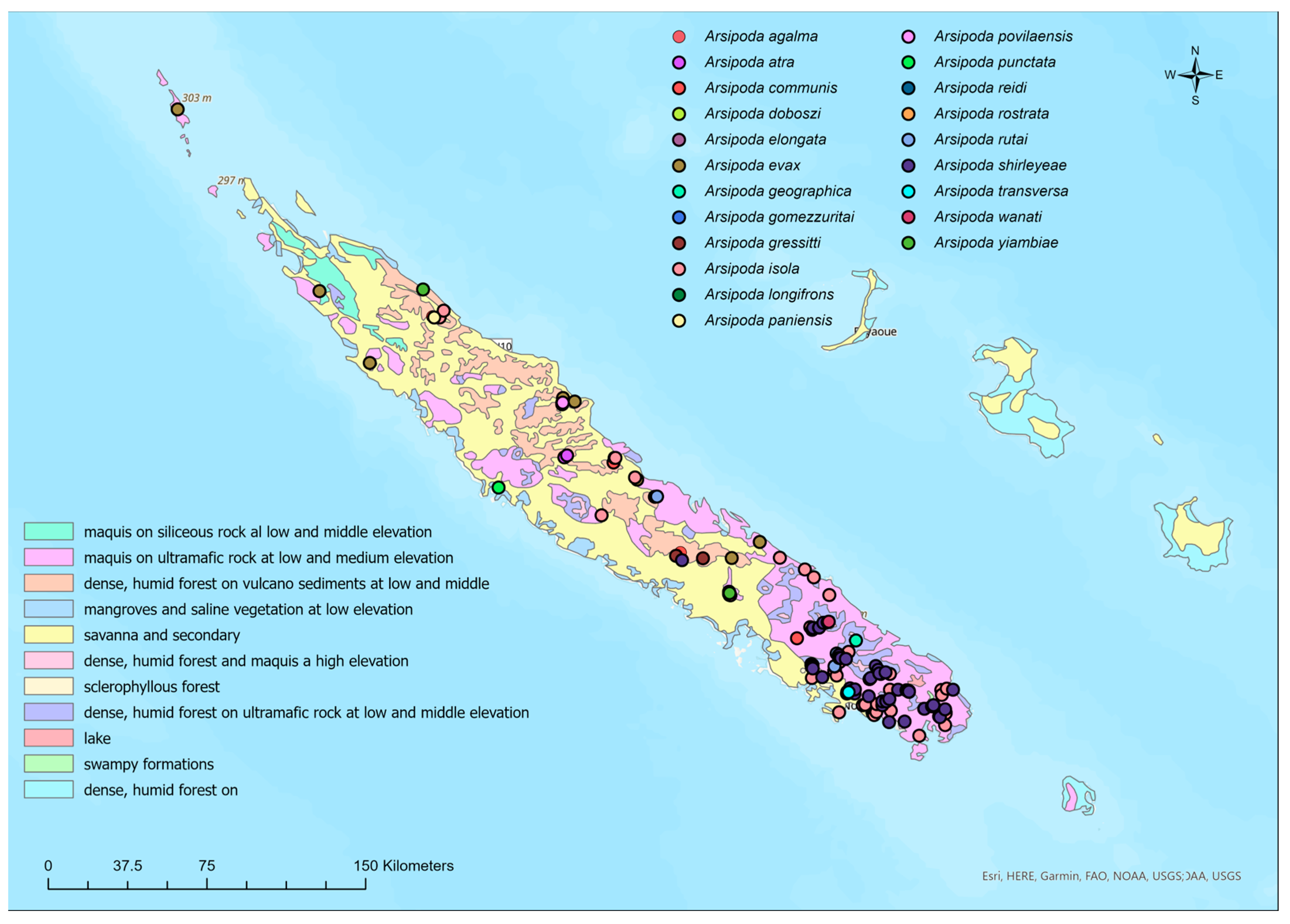
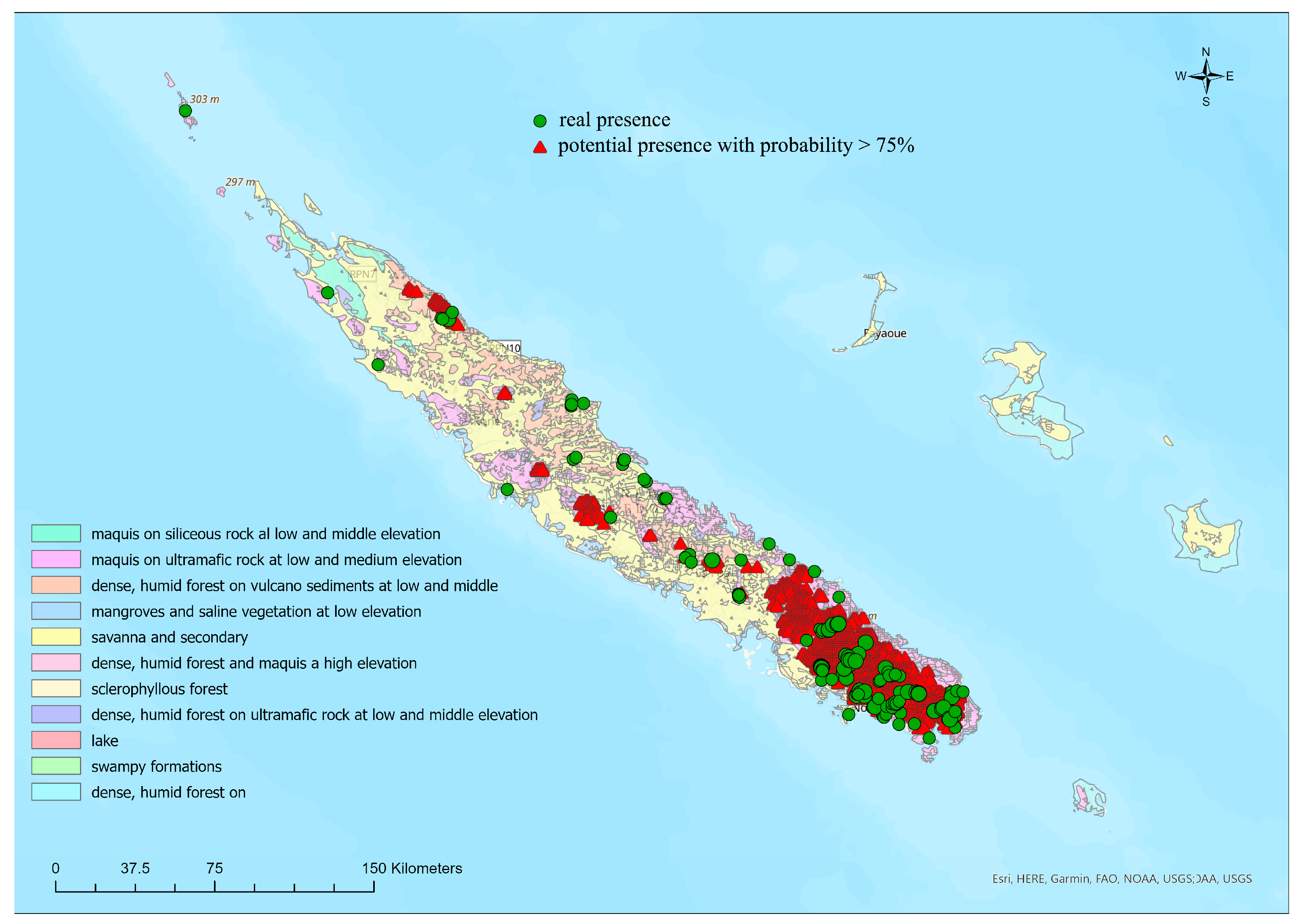
2.1. Study Area, Species Database, and Vegetation Formations
2.2. Model Building and Evaluation
2.3. Spatial and Statistical Analyses
3. Results
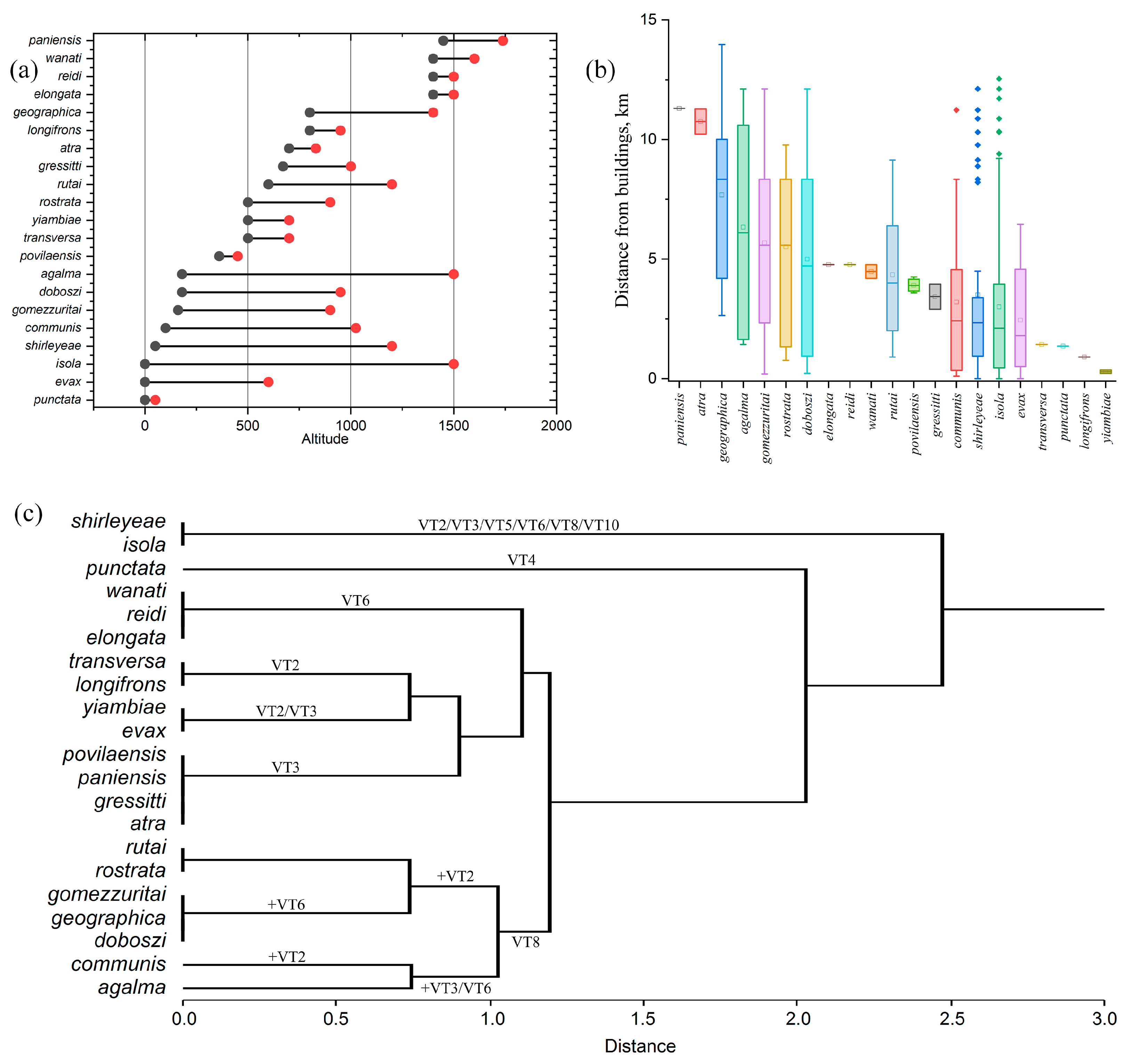
3.1. Habitat Preference, and Altitudinal Distribution
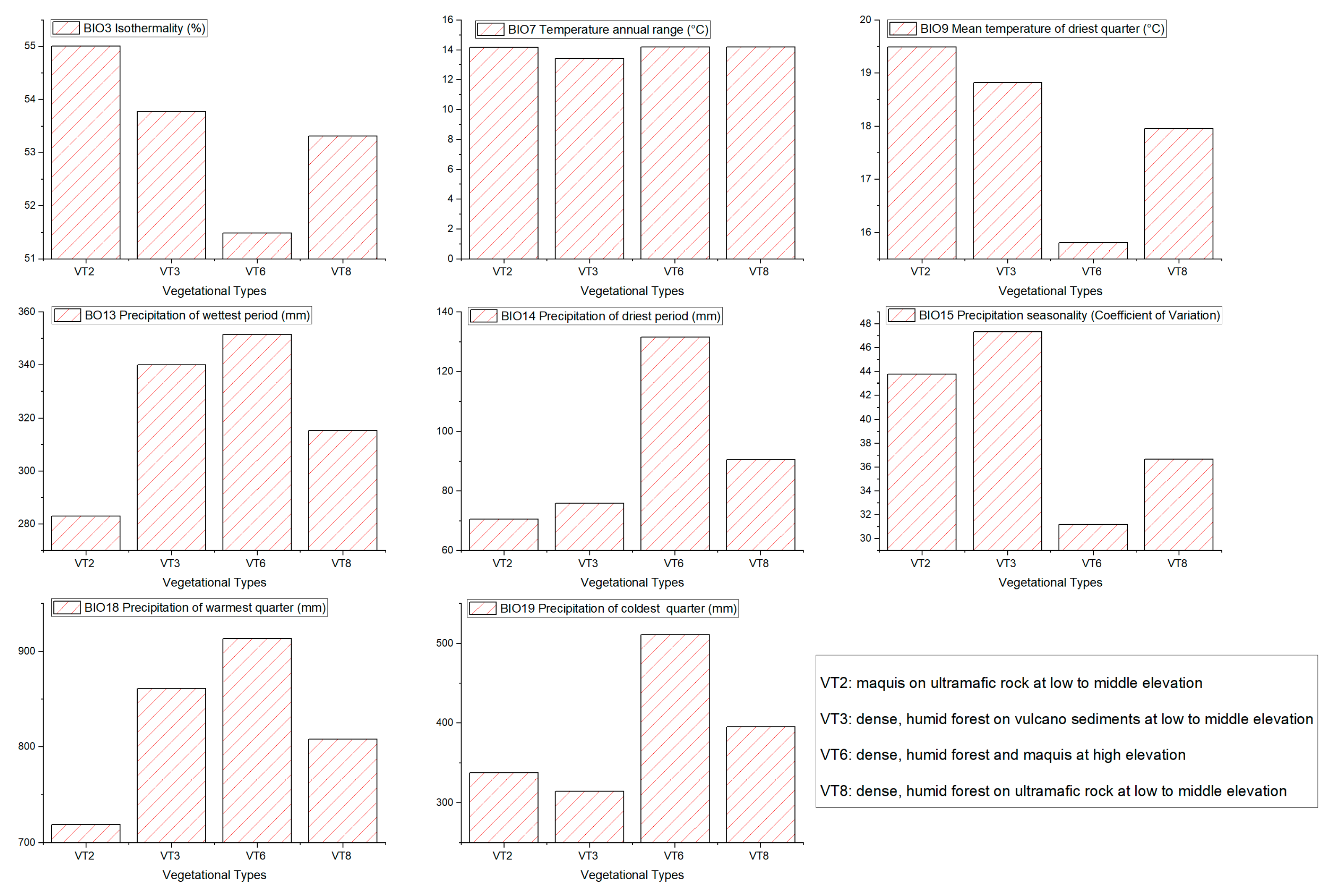
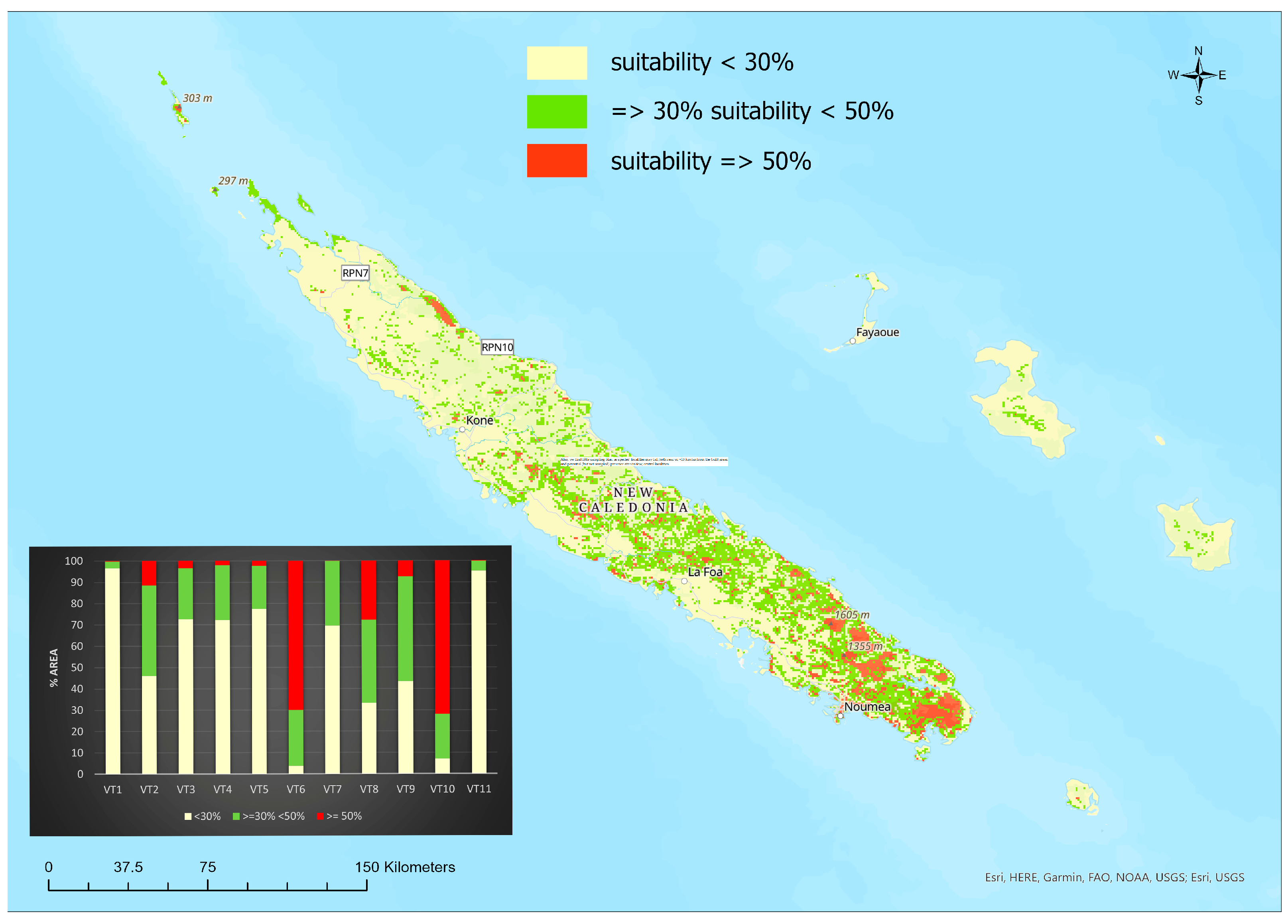
3.2. Ecological Niche Modelling and Vegetation Types
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pellens, R.; Grandcolas, P. Conservation and Management of the Biodiversity in a Hotspot Characterized by Short Range Endemism and Rarity: The Challenge of New Caledonia. In Biodiversity Hotspots; Rescigno, V., Maletta, S., Eds.; Nova Science Publishers: Hauppauge, NY, USA, 2009; pp. 139–151. [Google Scholar]
- Pillon, Y.; Jaffré, T.; Birnbaum, P.; Bruy, D.; Cluzel, D.; Ducousso, M.; Fogliani, B.; Ibanez, T.; Jourdan, H.; Lagarde, L. Infertile Landscapes on an Old Oceanic Island: The Biodiversity Hotspot of New Caledonia. Biol. J. Linn. Soc. 2021, 133, 317–341. [Google Scholar] [CrossRef]
- New Caledonia Ecosystem Profile Working Group. Delineation of Key Biodiversity Areas, New Caledonia; New Caledonia Ecosystem Profile Working Group: Arlington, VA, USA, 2011. [Google Scholar]
- Wulff, A.S.; Hollingsworth, P.M.; Ahrends, A.; Jaffré, T.; Veillon, J.-M.; L’Huillier, L.; Fogliani, B. Conservation Priorities in a Biodiversity Hotspot: Analysis of Narrow Endemic Plant Species in New Caledonia. PLoS ONE 2013, 8, e73371. [Google Scholar] [CrossRef] [PubMed]
- Caesar, M.; Grandcolas, P.; Pellens, R. Outstanding Micro-Endemism in New Caledonia: More than One out of Ten Animal Species Have a Very Restricted Distribution Range. PLoS ONE 2017, 12, e0181437. [Google Scholar] [CrossRef] [PubMed]
- Nattier, R.; Pellens, R.; Robillard, T.; Jourdan, H.; Legendre, F.; Caesar, M.; Nel, A.; Grandcolas, P. Updating the Phylogenetic Dating of New Caledonian Biodiversity with a Meta-Analysis of the Available Evidence. Sci. Rep. 2017, 7, 3705. [Google Scholar] [CrossRef]
- Giribet, G.; Baker, C.M. Further Discussion on the Eocene Drowning of New Caledonia: Discordances from the Point of View of Zoology. J. Biogeogr. 2019, 46, 1912–1918. [Google Scholar] [CrossRef]
- Grandcolas, P. The Origin of Diversity in Insects: Speciation, Adaptation and the Earth Dynamics. Comptes Rendus. Biol. 2019, 342, 252–253. [Google Scholar] [CrossRef]
- Jolivet, P.; Verma, K. On the Origin of the Chrysomelid Fauna of New Caledonia. Res. Chrysomelidae 2008, 1, 309–319. [Google Scholar]
- Jolivet, P.; Verma, K.K. Biogeography and Biology of the New Caledonian Chrysomelidae (Coleoptera). Res. Chrysomelidae 2009, 2, 211–223. [Google Scholar]
- Beenen, R. Contribution to the Knowledge of Galerucinae of New Caledonia (Coleoptera: Chrysomelidae). Genus 2008, 19, 65–87. [Google Scholar]
- Beenen, R. Contribution to the Knowledge of Galerucinae of New Caledonia 2 (Coleoptera: Chrysomelidae). Genus 2013, 24, 65–108. [Google Scholar]
- Gòmez-Zurita, J.; Cardoso, A. Systematics of the New Caledonian Endemic Genus Taophila Heller (Coleoptera: Chrysomelidae, Eumolpinae) Combining Morphological, Molecular and Ecological Data, with Description of Two New Species. Syst. Entomol. 2014, 39, 111–126. [Google Scholar] [CrossRef]
- Gómez-Zurita, J.; Cardoso, A.; Jurado-Rivera, J.A.; Jolivet, P.; Cazères, S.; Mille, C. Discovery of New Species of New Caledonian Arsipoda Erichson, 1842 (Coleoptera: Chrysomelidae) and Insights on Their Ecology and Evolution Using DNA Markers. J. Nat. Hist. 2010, 44, 2557–2579. [Google Scholar] [CrossRef]
- Samuelson, G.A. Review of Taophila, a Genus Endemic to New Caledonia (Coleoptera: Chrysomelidae: Eumolpinae). Zootaxa 2010, 2621, 45–62. [Google Scholar] [CrossRef]
- D’Alessandro, P.; Samuelson, A.; Biondi, M. Taxonomic Revision of the Genus Arsipoda Erichson, 1842 (Coleoptera, Chrysomelidae) in New Caledonia. Eur. J. Taxon. 2016, 230, 1–61. [Google Scholar] [CrossRef]
- Borowiec, L.; Świętojańska, J.; Sekerka, L. Revision of the Tribe Cryptonychini (Coleoptera: Chrysomelidae: Cassidinae) of New Caledonia. Zootaxa 2019, 4690, 1. [Google Scholar] [CrossRef]
- Platania, L.; Cardoso, A.; Gómez-Zurita, J. Diversity and Evolution of New Caledonian Endemic Taophila Subgenus Lapita (Coleoptera: Chrysomelidae: Eumolpinae). Zool. J. Linn. Soc. 2020, 189, 1123–1154. [Google Scholar] [CrossRef]
- Gómez-Zurita, J. Integrative Systematic Revision of a New Genus of Eumolpinae (Coleoptera: Chrysomelidae) Endemic to New Caledonia: Dematotrichus gen. nov. and Its Numerous New Hairy Species. Syst. Biodivers. 2022, 20, 1–28. [Google Scholar] [CrossRef]
- Platania, L.; Gómez-Zurita, J. Integrative Taxonomic Revision of the New Caledonian Endemic Genus Taophila Heller (Coleoptera: Chrysomelidae, Eumolpinae). Insect Syst. Evol. 2021, 53, 111–184. [Google Scholar] [CrossRef]
- Bouchard, P.; Bousquet, Y.; Davies, A.E.; Alonso-Zarazaga, M.A.; Lawrence, J.F.; Lyal, C.H.; Newton, A.F.; Reid, C.A.; Schmitt, M.; Ślipiński, S.A. Family-Group Names in Coleoptera (Insecta). ZooKeys 2011, 88, 1–972. [Google Scholar]
- Nadein, K.; Betz, O. Jumping Mechanisms and Performance in Beetles. I. Flea Beetles (Coleoptera: Chrysomelidae: Alticini). J. Exp. Biol. 2016, 219, 2015–2027. [Google Scholar] [CrossRef]
- Biondi, M.; Iannella, M.; D’Alessandro, P. Unravelling the Taxonomic Assessment of an Interesting New Species from Socotra Island: Blepharidina socotrana sp. nov. (Coleoptera: Chrysomelidae). Acta Entomol. Musei Natl. Pragae 2019, 59, 499–505. [Google Scholar] [CrossRef]
- Nadein, K.S. Catalogue of Alticini Genera of the World (Coleoptera: Chrysomelidae). Available online: http://www.zin.ru/Animalia/Coleoptera/eng/alticinw.htm (accessed on 11 November 2022).
- Nadein, K.S.; Beždek, J. Galerucinae Latreille 1802. In Coleoptera, Beetles, Volume 3: Morphology and Systematics (Phytophaga); Leschen, R.A.B., Beutel, R.G., Eds.; De Gruyter: Berlin, Germany, 2014; Volume 4, ISBN 3-11-027446-9. [Google Scholar]
- Biondi, M.; D’Alessandro, P. Longitarsus doeberli, a Wingless New Species from Socotra Island (Coleoptera: Chrysomelidae). Acta Entomol. Musei Natl. Pragae 2017, 57, 165–172. [Google Scholar] [CrossRef][Green Version]
- Biondi, M.; D’Alessandro, P. Taxonomical Revision of the Longitarsus capensis Species-Group: An Example of Mediterranean-Southern African Disjunct Distributions (Coleoptera: Chrysomelidae). Eur. J. Entomol. 2008, 105, 719–736. [Google Scholar] [CrossRef]
- Biondi, M.; D’Alessandro, P. Genus-Group Names of Afrotropical Flea Beetles (Coleoptera: Chrysomelidae: Alticinae): Annotated Catalogue and Biogeographical Notes. Eur. J. Entomol. 2010, 107, 401–424. [Google Scholar] [CrossRef]
- Biondi, M.; D’Alessandro, P. Afrotropical Flea Beetle Genera: A Key to Their Identification, Updated Catalogue and Biogeographical Analysis (Coleoptera, Chrysomelidae, Galerucinae, Alticini). ZooKeys 2012, 253, 1–158. [Google Scholar] [CrossRef]
- Biondi, M.; Urbani, F.; D’Alessandro, P. Relationships between the Geographic Distribution of Phytophagous Insects and Different Types of Vegetation: A Case Study of the Flea Beetle Genus Chaetocnema (Coleoptera: Chrysomelidae) in the Afrotropical Region. Eur. J. Entomol. 2015, 112, 311–327. [Google Scholar] [CrossRef]
- D’Alessandro, P.; Iannella, M.; Frasca, R.; Biondi, M. Distribution Patterns and Habitat Preference for the Genera-Group Blepharida Sl in Sub-Saharan Africa (Coleoptera: Chrysomelidae: Galerucinae: Alticini). Zool. Anz. 2018, 277, 23–32. [Google Scholar] [CrossRef]
- Jolivet, P.; Verma, K.K. Biology of Leaf Beetles; Intercept Limited: Hampshire, UK, 2002; ISBN 1-898298-86-6. [Google Scholar]
- Urbani, F.; D’Alessandro, P.; Frasca, R.; Biondi, M. Maximum Entropy Modeling of Geographic Distributions of the Flea Beetle Species Endemic in Italy (Coleoptera: Chrysomelidae: Galerucinae: Alticini). Zool. Anz. 2015, 258, 99–109. [Google Scholar] [CrossRef]
- Iannella, M.; D’Alessandro, P.; Biondi, M. Forecasting the Spread Associated with Climate Change in Eastern Europe of the Invasive Asiatic Flea Beetle, Luperomorpha xanthodera (Coleoptera: Chrysomelidae). Eur. J. Entomol. 2020, 117, 130–138. [Google Scholar] [CrossRef]
- De Simone, W.; Iannella, M.; D’Alessandro, P.; Biondi, M. Assessing Influence in Biofuel Production and Ecosystem Services When Environmental Changes Affect Plant—Pest Relationships. GCB Bioenergy 2020, 12, 864–877. [Google Scholar] [CrossRef]
- Iannella, M.; De Simone, W.; D’Alessandro, P.; Biondi, M. Climate Change Favours Connectivity between Virus-Bearing Pest and Rice Cultivations in Sub-Saharan Africa, Depressing Local Economies. PeerJ 2021, 9, e12387. [Google Scholar] [CrossRef] [PubMed]
- Scherer, G. Die Alticinae des Indischen Subkontinentes (Coleoptera-Chrysomelidae); Pacific Insects Monograph: Honolulu, Hawaii, 1969; Volume 22. [Google Scholar]
- Seeno, T.N.; Wilcox, J.A. Leaf Beetle Genera (Coleoptera, Chrysomelidae); Entomography; Entomography Publications: Sacramento, CA, USA, 1982; Volume 1. [Google Scholar]
- Biondi, M.; D’Alessandro, P. Arsipoda reidi, a New Replacement Name for Arsipoda montana D’Alessandro, Samuelson & Biondi (Coleoptera: Chrysomelidae). Fragm. Entomol. 2022, 54, 171–172. [Google Scholar]
- Mohamedsaid, M. Catalogue of the Malaysian Chrysomelidae (Insecta: Coleoptera); Pensoft Publishers: Sofia, Bulgaria, 2004; ISBN 954-642-201-0. [Google Scholar]
- Samuelson, G.A. Alticinae of Oceania (Coleoptera: Chrysomelidae); Pacific Insects Monograph: Honolulu, Hawaii, 1973; Volume 30, ISBN 9798659868010. [Google Scholar]
- Heikertinger, F.; Csiki, E. Chrysomelidae: Halticinae I. In Coleopterorum Catalogus; No. 166; Springer: Berlin/Heidelberg, Germany, 1940; pp. 52–56. [Google Scholar]
- Jaffré, T.; Rigault, F.; Munzinger, J. La Végétation. In Atlas de la Nouvelle-Calédonie; IRD-Congrès de la Nouvelle-Calédonie: Nouméa, France, 2012. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models: With Applications in R; Cambridge University Press: Cambridge, UK, 2017; ISBN 0-521-76513-7. [Google Scholar]
- Dormann, C.F. Effects of Incorporating Spatial Autocorrelation into the Analysis of Species Distribution Data. Glob. Ecol. Biogeogr. 2007, 16, 129–138. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel Methods Improve Prediction of Species’ Distributions from Occurrence Data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Naimi, B. Usdm: Uncertainty Analysis for Species Distribution Models. R Package Version 1.1–15. R Documentation. 2017. Available online: http://www.rdocumentation.org/packages/usdm (accessed on 15 June 2021).
- Smith, A.B.; Godsoe, W.; Rodríguez-Sánchez, F.; Wang, H.-H.; Warren, D. Niche Estimation above and below the Species Level. Trends Ecol. Evol. 2019, 34, 260–273. [Google Scholar] [CrossRef] [PubMed]
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. SpThin: An R Package for Spatial Thinning of Species Occurrence Records for Use in Ecological Niche Models. Ecography 2015, 38, 541–545. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: http://www.R-project.org/ (accessed on 20 July 2021).
- Thuiller, W.; Georges, D.; Engler, R. Biomod2: Ensemble Platform for Species Distribution Modeling. R Package Version 3.3-7. 2016. Available online: https://cran.microsoft.com/snapshot/2016-08-05/web/packages/biomod2/biomod2.pdf (accessed on 7 March 2022).
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the Accuracy of Species Distribution Models: Prevalence, Kappa and the True Skill Statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Console, G.; Iannella, M.; Cerasoli, F.; D’Alessandro, P.; Biondi, M. A European Perspective of the Conservation Status of the Threatened Meadow Viper Vipera ursinii (BONAPARTE, 1835) (Reptilia, Viperidae). Wildl. Biol. 2020, 2, 1–12. [Google Scholar] [CrossRef]
- Biondi, M.; D’Alessandro, P.; De Simone, W.; Iannella, M. DBSCAN and GIE, Two Density-Based “Grid-Free” Methods for Finding Areas of Endemism: A Case Study of Flea Beetles (Coleoptera, Chrysomelidae) in the Afrotropical Region. Insects 2021, 12, 1115. [Google Scholar] [CrossRef]
- Iannella, M.; D’Alessandro, P.; Biondi, M. Entomological Knowledge in Madagascar by GBIF Datasets: Estimates on the Coverage and Possible Biases (Insecta). Fragm. Entomol. 2019, 51, 1–10. [Google Scholar] [CrossRef]
- ESRI Inc. ArcGIS Pro 3.0; ESRI Inc.: Redlands, CA, USA, 2022. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | VT1 | VT2 | VT3 | VT4 | VT5 | VT6 | VT7 | VT8 | VT9 | VT10 | VT11 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Arsipoda agalma Samuelson, 1973 | 0 | 0 | 4 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 |
| Arsipoda atra D’Alessandro, Samuelson, and Biondi, 2016 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Arsipoda communis D’Alessandro, Samuelson, and Biondi, 2016 | 0 | 19 | 2 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 |
| Arsipoda doboszi D’Alessandro, Samuelson, and Biondi, 2016 | 0 | 11 | 0 | 0 | 0 | 2 | 0 | 1 | 0 | 0 | 0 |
| Arsipoda elongata D’Alessandro, Samuelson, and Biondi, 2016 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| Arsipoda evax Samuelson, 1973 | 0 | 9 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Arsipoda geographica Gómez-Zurita, 2010 | 0 | 2 | 0 | 0 | 0 | 5 | 0 | 3 | 0 | 0 | 0 |
| Arsipoda gomezzuritai D’Alessandro, Samuelson, and Biondi, 2016 | 0 | 9 | 0 | 0 | 0 | 2 | 0 | 2 | 0 | 0 | 0 |
| Arsipoda gressitti D’Alessandro, Samuelson, and Biondi, 2016 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Arsipoda isola Samuelson, 1973 | 0 | 64 | 1 | 0 | 11 | 3 | 0 | 12 | 0 | 2 | 0 |
| Arsipoda longifrons D’Alessandro, Samuelson, and Biondi, 2016 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Arsipoda paniensis D’Alessandro, Samuelson, and Biondi, 2016 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Arsipoda povilaensis D’Alessandro, Samuelson, and Biondi, 2016 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Arsipoda punctata D’Alessandro, Samuelson, and Biondi, 2016 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Arsipoda reidi Biondi and D’Alessandro, 2022 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| Arsipoda rostrata Gómez-Zurita, 2010 | 0 | 6 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Arsipoda rutai D’Alessandro, Samuelson, and Biondi, 2016 | 0 | 9 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 |
| Arsipoda shirleyeae Samuelson, 1973 | 0 | 33 | 1 | 0 | 2 | 2 | 0 | 16 | 0 | 3 | 0 |
| Arsipoda transversa D’Alessandro, Samuelson, and Biondi, 2016 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Arsipoda wanati D’Alessandro, Samuelson, and Biondi, 2016 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 |
| Arsipoda yiambiae Samuelson, 1973 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vegetation Type | Area (km2) | No. of Species | No. of Occurrences |
|---|---|---|---|
| VT1—maquis on siliceous rock at low to middle elevation | 530 | 0 | 0 |
| VT2—maquis on ultramafic rock at low to middle elevation | 4451 | 12 | 165 |
| VT3—dense, humid forest on volcano sediments at low to middle elevation | 2275 | 10 | 22 |
| VT4—mangroves and saline vegetation at low elevation | 628 | 1 | 1 |
| VT5—savanna and secondary brushwood | 8411 | 2 | 13 |
| VT6—dense, humid forest and maquis at high elevation | 132 | 10 | 20 |
| VT7—sclerophyllous forest | 16 | 0 | 0 |
| VT8—dense, humid forest on ultramafic rock at low to middle elevation | 1097 | 9 | 40 |
| VT9—lake | 23 | 0 | 0 |
| VT10—swampy formations | 43 | 2 | 5 |
| VT11—dense, humid forest on calcareous rock | 1254 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biondi, M.; D’Alessandro, P.; Iannella, M. Climatic Niche, Altitudinal Distribution, and Vegetation Type Preference of the Flea Beetle Genus Arsipoda in New Caledonia (Coleoptera Chrysomelidae). Insects 2023, 14, 19. https://doi.org/10.3390/insects14010019
Biondi M, D’Alessandro P, Iannella M. Climatic Niche, Altitudinal Distribution, and Vegetation Type Preference of the Flea Beetle Genus Arsipoda in New Caledonia (Coleoptera Chrysomelidae). Insects. 2023; 14(1):19. https://doi.org/10.3390/insects14010019
Chicago/Turabian StyleBiondi, Maurizio, Paola D’Alessandro, and Mattia Iannella. 2023. "Climatic Niche, Altitudinal Distribution, and Vegetation Type Preference of the Flea Beetle Genus Arsipoda in New Caledonia (Coleoptera Chrysomelidae)" Insects 14, no. 1: 19. https://doi.org/10.3390/insects14010019
APA StyleBiondi, M., D’Alessandro, P., & Iannella, M. (2023). Climatic Niche, Altitudinal Distribution, and Vegetation Type Preference of the Flea Beetle Genus Arsipoda in New Caledonia (Coleoptera Chrysomelidae). Insects, 14(1), 19. https://doi.org/10.3390/insects14010019