
Olfactory Strategies in the Defensive Behaviour of Insects


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
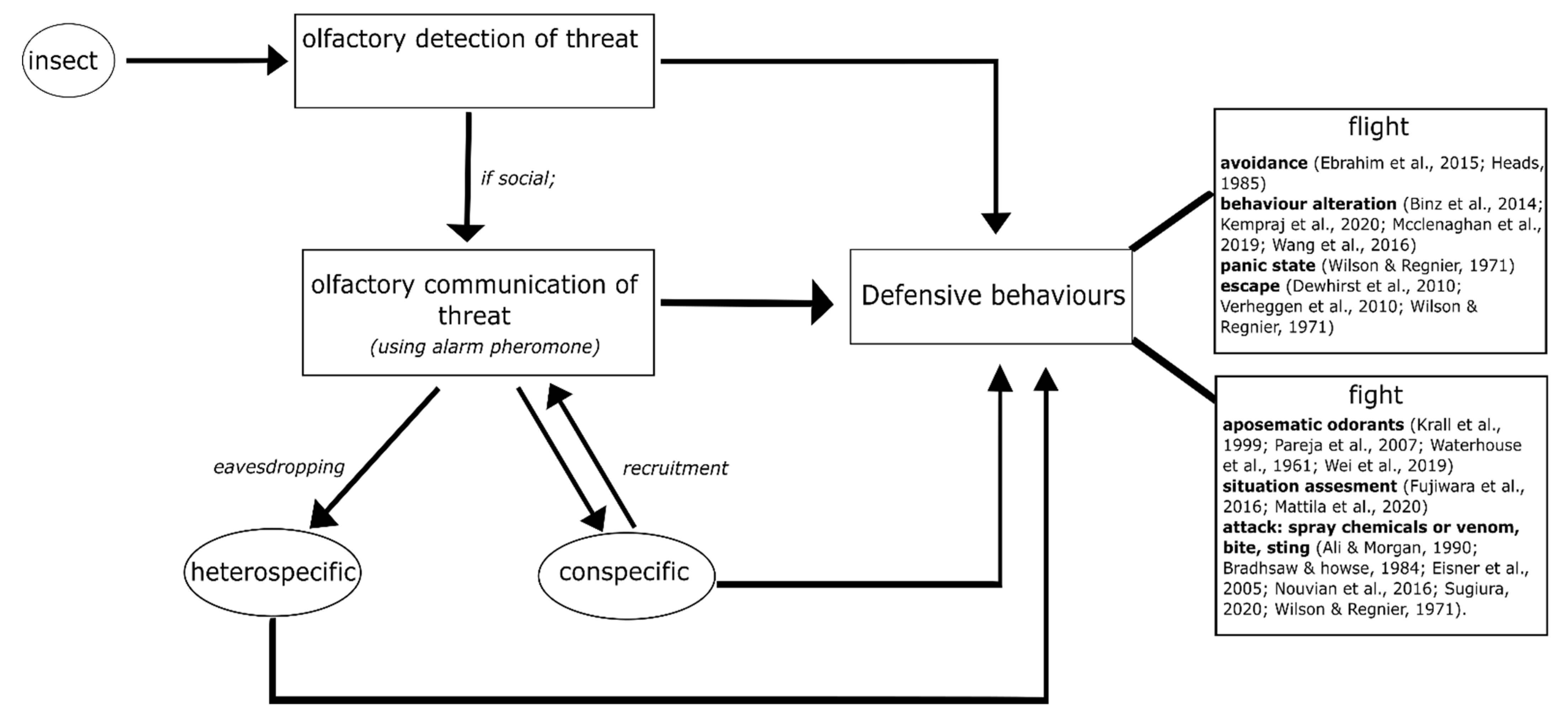
2. Olfaction in Defensive Behaviours of Insects
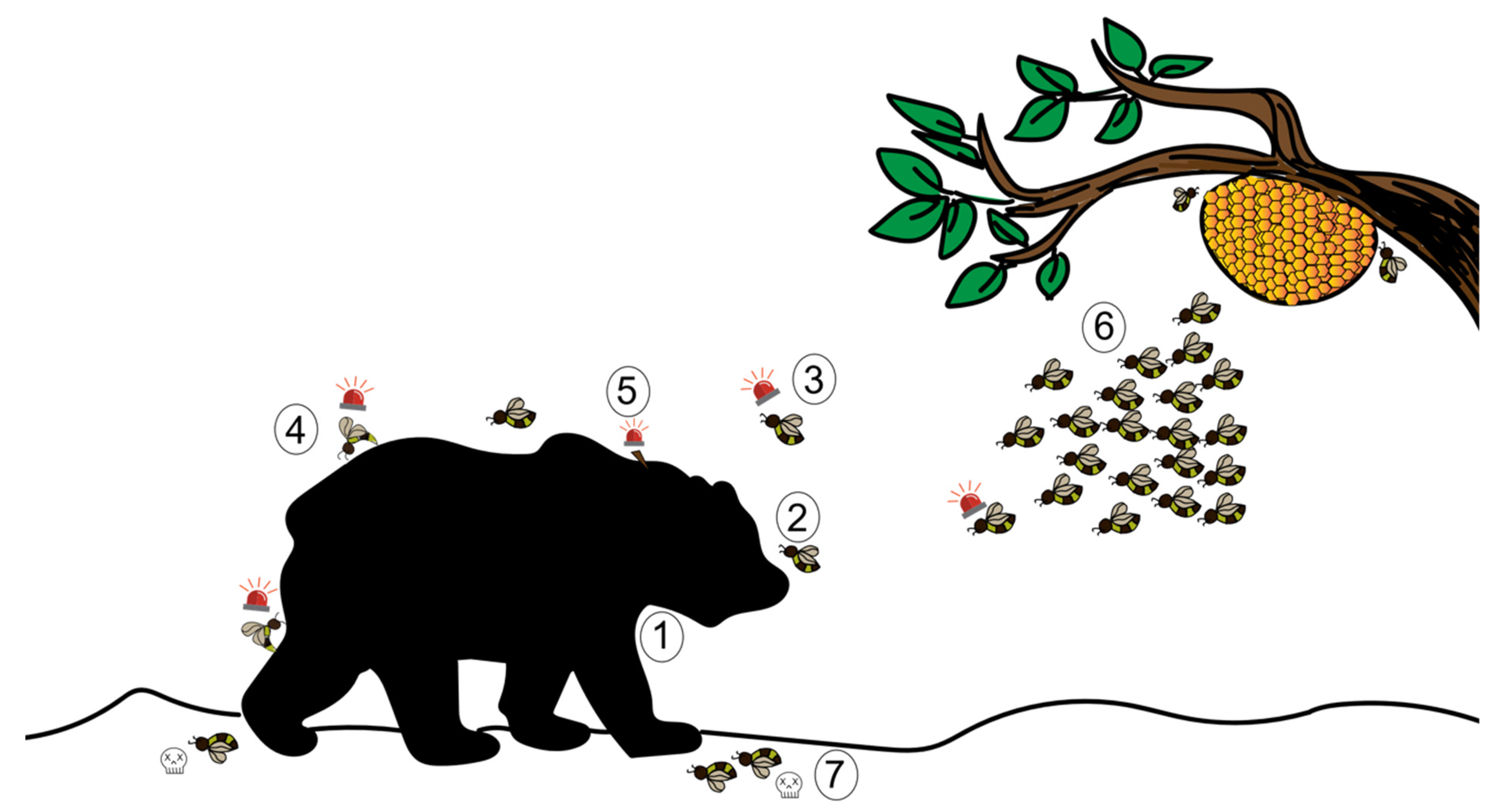
3. Defensive Behaviour Based on Olfactory Detection
4. Defensive Behaviour Based on Olfactory Communication
5. The Sting and Mandibular Alarm Pheromones
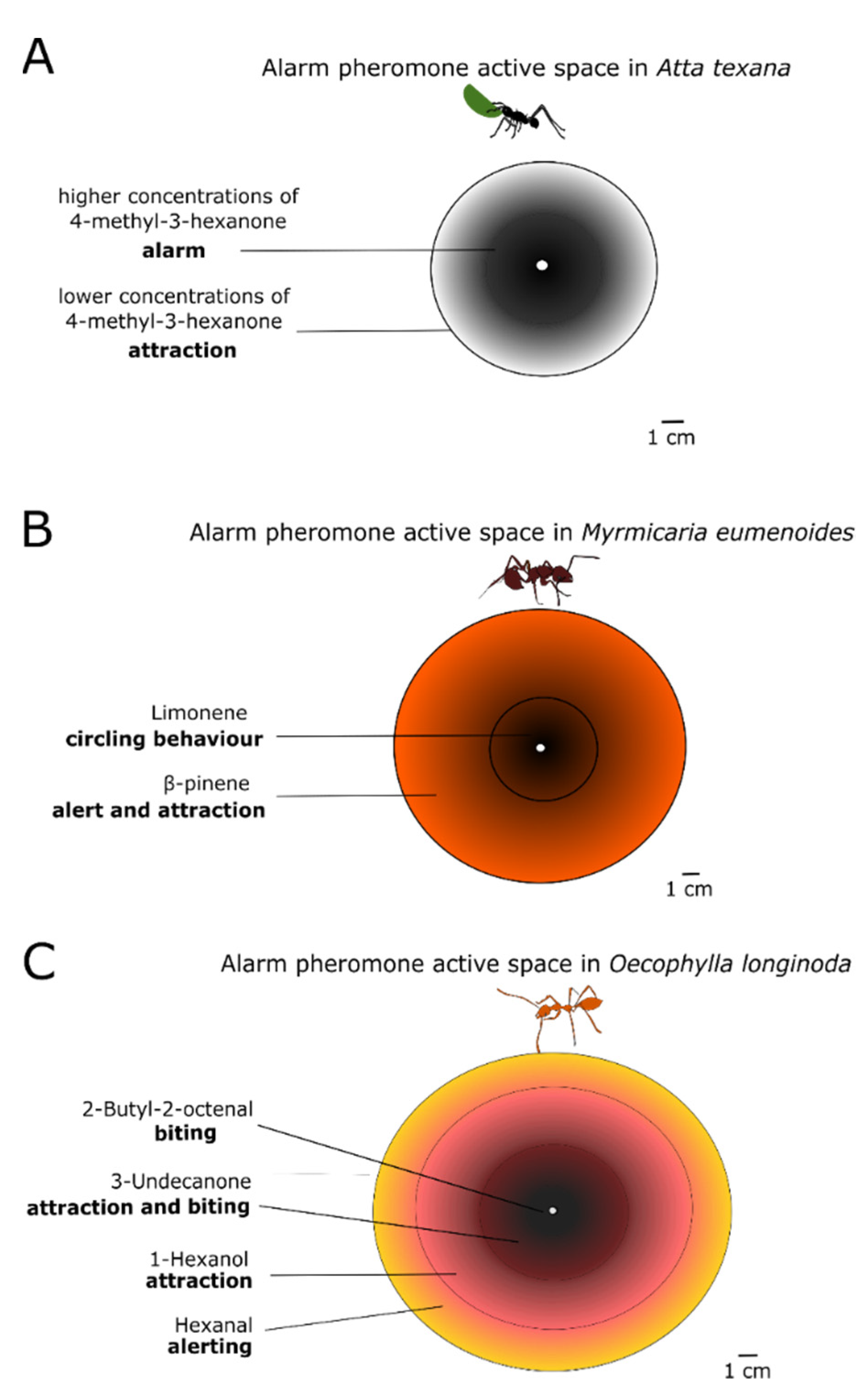
6. Chemical Properties of Alarm Pheromones
7. Effects of the Alarm Pheromone in Different Behavioural Contexts
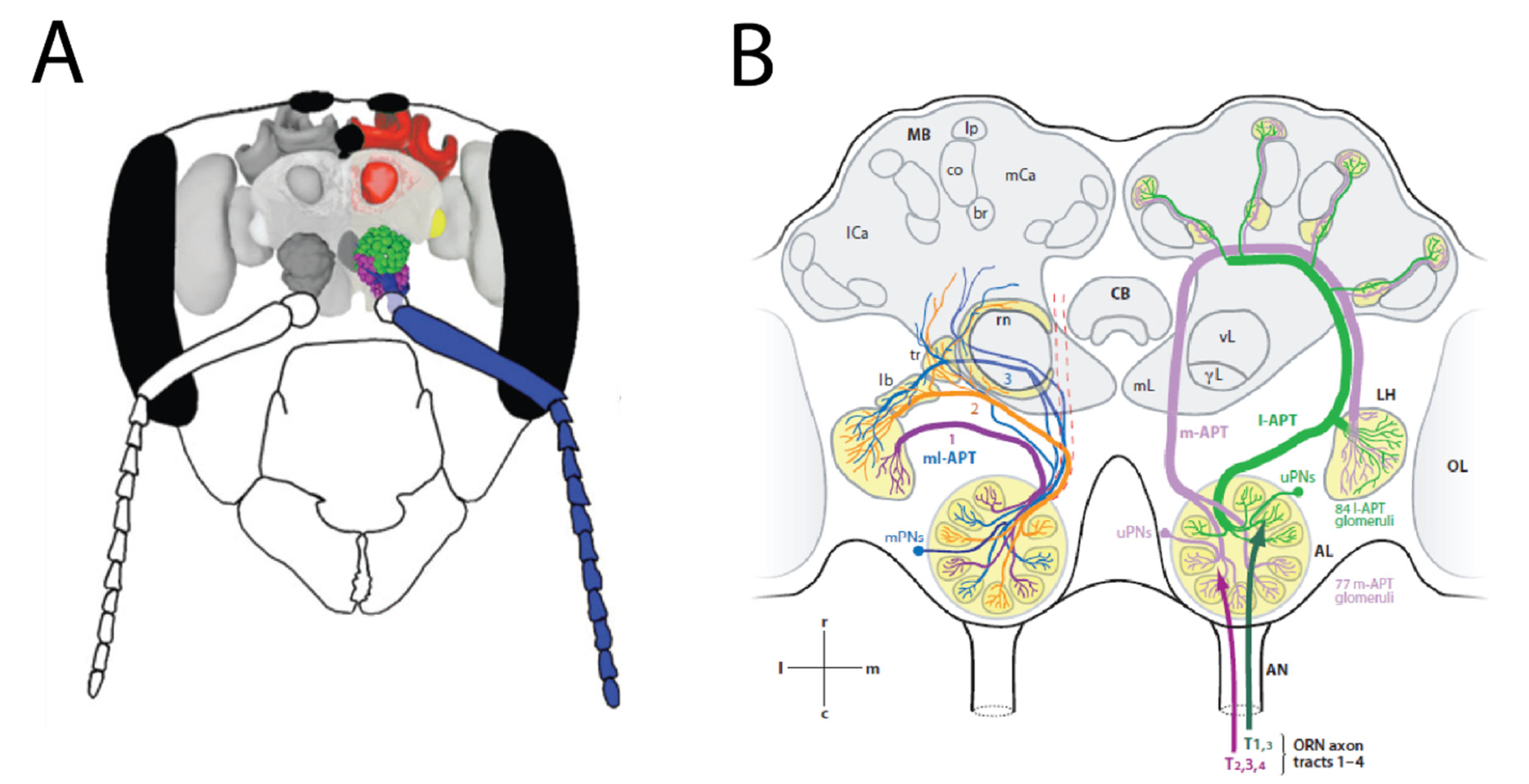
8. Neurobiology of Olfactory Coding for Defensive Mechanisms in Insects
9. Outlook to Future Research
Author Contributions
Funding
Conflicts of Interest
References
- Lima, S.L.; Dill, L.M. Behavioral decisions made under the risk of predation: A review and prospectus. Can. J. Zool. 1990, 68, 619–640. [Google Scholar] [CrossRef]
- Heads, P.A. The effect of invertebrate and vertebrate predators on the foraging movements of Ischnura elegans larvae (Odonata: Zygoptera). Freshw. Biol. 1985, 15, 559–571. [Google Scholar] [CrossRef]
- Wirsing, A.J.; Heithaus, M.R.; Brown, J.S.; Kotler, B.P.; Schmitz, O.J. The context dependence of non-consumptive predator effects. Ecol. Lett. 2021, 24, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Kempraj, V.; Park, S.J.; Taylor, P.W. Forewarned is forearmed: Queensland fruit flies detect olfactory cues from predators and respond with predator-specific behaviour. Sci. Rep. 2020, 10, 7297. [Google Scholar] [CrossRef] [PubMed]
- Binz, H.; Bucher, R.; Entling, M.H.; Menzel, F. Knowing the Risk: Crickets Distinguish between Spider Predators of Different Size and Commonness. Ethology 2014, 120, 99–110. [Google Scholar] [CrossRef]
- Ebrahim, S.A.M.; Dweck, H.K.M.; Stökl, J.; Hofferberth, J.E.; Trona, F.; Weniger, K.; Rybak, J.; Seki, Y.; Stensmyr, M.C.; Sachse, S.; et al. Drosophila Avoids Parasitoids by Sensing Their Semiochemicals via a Dedicated Olfactory Circuit. PLoS Biol. 2015, 13, e1002318. [Google Scholar] [CrossRef]
- Goodale, E.; Nieh, J.C. Public use of olfactory information associated with predation in two species of social bees. Anim. Behav. 2012, 84, 919–924. [Google Scholar] [CrossRef]
- Li, J.; Wang, Z.; Tan, K.; Qu, Y.; Nieh, J. Effects of natural and synthetic alarm pheromone and individual pheromone components on foraging behavior of the giant Asian honey bee, Apis dorsata. J. Exp. Biol. 2014, 217, 3512–3518. [Google Scholar] [CrossRef]
- Bitterman, M.; Menzel, R.; Fietz, A.; Schäfer, S. Classical conditioning of proboscis extension in honeybees (Apis mellifera). J. Comp. Psychol. 1983, 97, 107. [Google Scholar] [CrossRef]
- Urlacher, E.; Francés, B.; Giurfa, M.; Devaud, J.-M. An alarm pheromone modulates appetitive olfactory learning in the honeybee (Apis mellifera). Front. Behav. Neurosci. 2010, 4, 157. [Google Scholar] [CrossRef]
- Wang, Z.; Qu, Y.; Dong, S.; Wen, P.; Li, J.; Tan, K.; Menzel, R. Honey Bees Modulate Their Olfactory Learning in the Presence of Hornet Predators and Alarm Component. PLoS ONE 2016, 11, e0150399. [Google Scholar] [CrossRef] [PubMed]
- Mcclenaghan, B.; Schlaf, M.; Geddes, M.; Mazza, J.; Pitman, G.; Mccallum, K.; Rawluk, S.; Hand, K.; Otis, G.W. Behavioral responses of honey bees, Apis cerana and Apis mellifera, to Vespa mandarinia marking and alarm pheromones. J. Apic. Res. 2019, 58, 141–148. [Google Scholar] [CrossRef]
- Lucas, C.; Brossette, L.; Lefloch, L.; Dupont, S.; Christidès, J.-P.; Bagnères, A.-G. When predator odour makes groups stronger: Effects on behavioural and chemical adaptations in two termite species. Ecol. Entomol. 2018, 43, 513–524. [Google Scholar] [CrossRef]
- Fujiwara, A.; Sasaki, M.; Washitani, I. A scientific note on hive entrance smearing in Japanese Apis cerana induced by pre-mass attack scouting by the Asian giant hornet Vespa mandarinia. Apidologie 2016, 47, 789–791. [Google Scholar] [CrossRef]
- Mattila, H.R.; Otis, G.W.; Nguyen, L.T.P.; Pham, H.D.; Knight, O.M.; Phan, N.T. Honey bees (Apis cerana) use animal feces as a tool to defend colonies against group attack by giant hornets (Vespa soror). PLoS ONE 2020, 15, e0242668. [Google Scholar] [CrossRef]
- Hosseini, M.; Mehrparvar, M.; Zytynska, S.E.; Hatano, E.; Weisser, W.W. Aphid alarm pheromone alters larval behaviour of the predatory gall midge, Aphidoletes aphidimyza and decreases intraguild predation by anthocorid bug, Orius laevigatus. Bull. Entomol. Res. 2021, 111, 445–453. [Google Scholar] [CrossRef]
- Sugiura, S. Predators as drivers of insect defenses. Entomol. Sci. 2020, 23, 316–337. [Google Scholar] [CrossRef]
- Eisner, T.; Eisner, M.; Siegler, M. Secret Weapons: Defenses of Insects, Spiders, Scorpions, and Other Many-Legged Creatures; Harvard University Press: Cambridge, MA, USA, 2005. [Google Scholar]
- Dean, J.; Aneshansley, D.; Edgerton, H.; Eisner, T. Defensive spray of the bombardier beetle: A biological pulse jet. Science 1990, 248, 1219–1221. [Google Scholar] [CrossRef]
- Eisner, T.; Jones, T.H.; Aneshansley, D.J.; Tschinkel, W.R.; Silberglied, R.E.; Meinwald, J. Chemistry of defensive secretions of bombardier beetles (Brachinini, Metriini, Ozaenini, Paussini). J. Insect Physiol. 1977, 23, 1383–1386. [Google Scholar] [CrossRef]
- Meinwald, J.; Chadha, M.S.; Hurst, J.J.; Eisner, I. Defense mechanisms of arthropods-IX anisomorphal, the secretion of a phasmid insect. Tetrahedron Lett. 1962, 3, 29–33. [Google Scholar] [CrossRef]
- Blum, M.S.; Warter, S.L.; Monroe, R.S.; Chidester, J.C. Chemical releasers of social behaviour—I. Methyl-n-amyl ketone in Iridomyrmex pruinosus (Roger) (Formicidae: Dolichoderinae). J. Insect Physiol. 1963, 9, 881–885. [Google Scholar] [CrossRef]
- Wilson, E.O.; Regnier, F.E., Jr. The Evolution of the Alarm-Defense System in the Formicine Ants. Am. Nat. 1971, 105, 279–289. [Google Scholar] [CrossRef]
- James, C.C.; Sánchez, D.; Cruz-López, L.; Nieh, J. Fighting ability and the toxicity of raiding pheromone in an obligate kleptoparasite, the stingless bee Lestrimelitta niitkib. Behav. Ecol. Sociobiol. 2022, 76, 38. [Google Scholar] [CrossRef]
- Eisner, T.; Grant, R.P. Toxicity, Odor Aversion, and “Olfactory Aposematism”. Science 1981, 213, 476. [Google Scholar] [CrossRef] [PubMed]
- Krall, B.S.; Bartelt, R.J.; Lewis, C.J.; Whitman, D.W. Chemical defense in the stink bug Cosmopepla bimaculata. J. Chem. Ecol. 1999, 25, 2477–2494. [Google Scholar] [CrossRef]
- Pareja, M.; Borges, M.; Laumann, R.A.; Moraes, M.C.B. Inter- and intraspecific variation in defensive compounds produced by five neotropical stink bug species (Hemiptera: Pentatomidae). J. Insect Physiol. 2007, 53, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, D.F.; Forss, D.A.; Hackman, R.H. Characteristic odour components of the scent of stink bugs. J. Insect Physiol. 1961, 6, 113–121. [Google Scholar] [CrossRef]
- Wei, J.; Shao, W.; Cao, M.; Ge, J.; Yang, P.; Chen, L.; Wang, X.; Kang, L. Phenylacetonitrile in locusts facilitates an antipredator defense by acting as an olfactory aposematic signal and cyanide precursor. Sci. Adv. 2019, 5, eaav5495. [Google Scholar] [CrossRef]
- Vander Meer, R.K.; Morel, L. Nestmate recognition in ants. In Pheromone Communication in Social Insects; CRC Press: Boca Raton, FL, USA, 1998; Volume 79. [Google Scholar]
- Breed, M.D. Recognition Pheromones of the Honey Bee. BioScience 1998, 48, 463–470. [Google Scholar] [CrossRef]
- D’Ettorre, P.; Wenseleers, T.; Dawson, J.; Hutchinson, S.; Boswell, T.; Ratnieks, F.L.W. Wax combs mediate nestmate recognition by guard honeybees. Anim. Behav. 2006, 71, 773–779. [Google Scholar] [CrossRef]
- Dani, F.R.; Jones, G.R.; Corsi, S.; Beard, R.; Pradella, D.; Turillazzi, S. Nestmate Recognition Cues in the Honey Bee: Differential Importance of Cuticular Alkanes and Alkenes. Chem. Senses 2005, 30, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.W.; Blomquist, G.J. Ecological, Behavioural, and biochemical aspects of insect hydrocarbons. Annu. Rev. Entomol. 2005, 50, 371–393. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.W.; Mcdaniel, C.A.; Blomquist, G.J. Cuticular hydrocarbons of the eastern subterranean termite, Reticulitermes flavipes (Kollar) (Isoptera: Rhinotermitidae). J. Chem. Ecol. 1978, 4, 233–245. [Google Scholar] [CrossRef]
- Singer, T.L. Roles of Hydrocarbons in the Recognition Systems of Insects. Am. Zool. 1998, 38, 394–405. [Google Scholar] [CrossRef]
- Van Zweden, J.S.; d’Ettorre, P. Nestmate recognition in social insects and the role of hydrocarbons. Insect Hydrocarb. Biol. Biochem. Chem. Ecol. 2010, 11, 222–243. [Google Scholar]
- Guerrieri, F.J.; Nehring, V.; Jørgensen, C.G.; Nielsen, J.; Galizia, C.G.; D’Ettorre, P. Ants recognize foes and not friends. Proc. R. Soc. B Biol. Sci. 2009, 276, 2461–2468. [Google Scholar] [CrossRef]
- Lorenzi, M.C.; d’Ettorre, P. Nestmate Recognition in Social Insects: What Does It Mean to Be Chemically Insignificant? Front. Ecol. Evol. 2020, 7, 488. [Google Scholar] [CrossRef]
- Breed, M.D. Nestmate recognition assays as a tool for population and ecological studies in eusocial insects: A review. J. Kans. Entomol. Soc. 2003, 76, 539–550. [Google Scholar]
- Blomquist, G.J.; Bagnères, A.-G. Insect Hydrocarbons: Biology, Biochemistry, and Chemical Ecology; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Ozaki, M.; Hefetz, A. Neural Mechanisms and Information Processing in Recognition Systems. Insects 2014, 5, 722–741. [Google Scholar] [CrossRef]
- Wyatt, T.D. Pheromones and Animal Behavior: Chemical Signals and Signatures; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Karlson, P.; Lüscher, M. The Proposed Biological Term ‘Pheromone’. Nature 1959, 183, 1835. [Google Scholar] [CrossRef]
- Slessor, K.N.; Winston, M.L.; Le Conte, Y. Pheromone Communication in the Honeybee (Apis mellifera L.). J. Chem. Ecol. 2005, 31, 2731–2745. [Google Scholar] [CrossRef]
- Grüter, C.; Czaczkes, T.J. Communication in social insects and how it is shaped by individual experience. Anim. Behav. 2019, 151, 207–215. [Google Scholar] [CrossRef]
- Bossert, W.H.; Wilson, E.O. The analysis of olfactory communication among animals. J. Theor. Biol. 1963, 5, 443–469. [Google Scholar] [CrossRef]
- Ali, M.F.; Morgan, E.D. Chemical communication in insect communities: A guide to insect pheromones with special emphasis on social insects. Biol. Rev. 1990, 65, 227–247. [Google Scholar] [CrossRef]
- Basu, S.; Clark, R.E.; Fu, Z.; Lee, B.W.; Crowder, D.W. Insect alarm pheromones in response to predators: Ecological trade-offs and molecular mechanisms. Insect Biochem. Mol. Biol. 2021, 128, 103514. [Google Scholar] [CrossRef]
- Maschwitz, U.W. Alarm Substances and Alarm Behaviour in Social Hymenoptera. Nature 1964, 204, 324–327. [Google Scholar] [CrossRef]
- Wager, B.R.; Breed, M.D. Does Honey Bee Sting Alarm Pheromone Give Orientation Information to Defensive Bees? Ann. Entomol. Soc. Am. 2000, 93, 1329–1332. [Google Scholar] [CrossRef]
- Verheggen, F.J.; Haubruge, E.; Mescher, M.C. Alarm Pheromones—Chemical Signaling in Response to Danger. In Vitamins & Hormones; Elsevier: Amsterdam, The Netherlands, 2010; pp. 215–239. [Google Scholar]
- Dewhirst, S.Y.; Pickett, J.A.; Hardie, J. Aphid pheromones. Vitam. Horm. 2010, 83, 551–574. [Google Scholar]
- Han, S.; Chen, W.; Elgar, M.A. An ambiguous function of an alarm pheromone in the collective displays of the Australian meat ant, Iridomyrmex purpureus. Ethology 2022, 128, 70–76. [Google Scholar] [CrossRef]
- Hughes, W.O.H.; Howse, P.E.; Vilela, E.F.; Knapp, J.J.; Goulson, D. Field Evaluation of Potential of Alarm Pheromone Compounds to Enhance Baits for Control of Grass-Cutting Ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2002, 95, 537–543. [Google Scholar] [CrossRef]
- Wang, Z.; Wen, P.; Qu, Y.; Dong, S.; Li, J.; Tan, K.; Nieh, J.C. Bees eavesdrop upon informative and persistent signal compounds in alarm pheromones. Sci. Rep. 2016, 6, 25693. [Google Scholar] [CrossRef]
- Wen, P.; Cheng, Y.; Qu, Y.; Zhang, H.; Li, J.; Bell, H.; Tan, K.; Nieh, J. Foragers of sympatric Asian honey bee species intercept competitor signals by avoiding benzyl acetate from Apis cerana alarm pheromone. Sci. Rep. 2017, 7, 6721. [Google Scholar] [CrossRef]
- Wells, R.L.; Frost, C.J.; Yanoviak, S.P. Effects of Azteca trigona alarm pheromones on heterospecific ant behavior. Insectes Sociaux 2021, 68, 359–365. [Google Scholar] [CrossRef]
- Sharma, K.; Vander Meer, R.K.; Fadamiro, H.Y. Phorid fly, Pseudacteon tricuspis, response to alkylpyrazine analogs of a fire ant, Solenopsis invicta, alarm pheromone. J. Insect Physiol. 2011, 57, 939–944. [Google Scholar] [CrossRef]
- Du, Y.; Grodowitz, M.J.; Chen, J. Electrophysiological Responses of Eighteen Species of Insects to Fire Ant Alarm Pheromone. Insects 2019, 10, 403. [Google Scholar] [CrossRef]
- Dong, S.; Wen, P.; Zhang, Q.; Wang, Y.; Cheng, Y.; Tan, K.; Nieh, J.C. Olfactory eavesdropping of predator alarm pheromone by sympatric but not allopatric prey. Anim. Behav. 2018, 141, 115–125. [Google Scholar] [CrossRef]
- Free, J.B. The stimuli releasing the stinging response of honeybees. Anim. Behav. 1961, 9, 193–196. [Google Scholar] [CrossRef]
- Ghent, R.L.; Gary, N.E. A Chemical Alarm Releaser in Honey Bee Stings (Apis mellifera L.). Psyche J. Entomol. 1962, 69, 1–6. [Google Scholar] [CrossRef]
- Cassier, P.; Tel-Zur, D.; Lensky, Y. The sting sheaths of honey bee workers (Apis mellifera L.): Structure and alarm pheromone secretion. J. Insect Physiol. 1994, 40, 23–32. [Google Scholar] [CrossRef]
- Grandperrin, D.; Cassier, P. Anatomy and ultrastructure of the Koschewnikow’s gland of the honey bee, Apis mellifera L. (Hymenoptera: Apidae). Int. J. Insect Morphol. Embryol. 1983, 12, 25–42. [Google Scholar] [CrossRef]
- Nouvian, M.; Hotier, L.; Claudianos, C.; Giurfa, M.; Reinhard, J. Appetitive floral odours prevent aggression in honeybees. Nat. Commun. 2015, 6, 10247. [Google Scholar] [CrossRef]
- Nouvian, M.; Reinhard, J.; Giurfa, M. The defensive response of the honeybee Apis mellifera. J. Exp. Biol. 2016, 219, 3505–3517. [Google Scholar] [CrossRef]
- Millor, J.; Pham-Delegue, M.; Deneubourg, J.L.; Camazine, S. Self-organized defensive behavior in honeybees. Proc. Natl. Acad. Sci. USA 1999, 96, 12611–12615. [Google Scholar] [CrossRef]
- Butler, C.G.; Free, J.B. The behaviour of worker honeybees at the hive entrance. Behaviour 1952, 4, 262–292. [Google Scholar] [CrossRef]
- Cheng, Y.-N.; Wen, P.; Dong, S.-H.; Tan, K.; Nieh, J.C. Poison and alarm: The Asian hornet Vespa velutina uses sting venom volatiles as alarm pheromone. J. Exp. Biol. 2016, 220, 645–651. [Google Scholar] [CrossRef]
- Santoro, D.; Hartley, S.; Suckling, D.M.; Lester, P.J. The stinging response of the common wasp (Vespula vulgaris): Plasticity and variation in individual aggressiveness. Insectes Sociaux 2015, 62, 455–463. [Google Scholar] [CrossRef]
- Ono, M.; Terabe, H.; Hori, H.; Sasaki, M. Components of giant hornet alarm pheromone. Nature 2003, 424, 637–638. [Google Scholar] [CrossRef]
- Riley, R.; Silverstein, R.; Moser, J.C. Isolation, identification, synthesis and biological activity of volatile compounds from the heads of Atta ants. J. Insect Physiol. 1974, 20, 1629–1637. [Google Scholar] [CrossRef]
- Butler, C.G. Mandibular Gland Pheromone of Worker Honeybees. Nature 1966, 212, 530. [Google Scholar] [CrossRef]
- Shearer, D.A.; Boch, R. 2-Heptanone in the Mandibular Gland Secretion of the Honey-bee. Nature 1965, 206, 530. [Google Scholar] [CrossRef]
- Alavez-Rosas, D.; Sánchez-Guillén, D.; Malo, E.A.; Cruz-López, L. (S)-2-Heptanol, the alarm pheromone of the stingless bee Melipona solani (Hymenoptera, Meliponini). Apidologie 2019, 50, 277–287. [Google Scholar] [CrossRef]
- Smith, B.; Roubik, D. Mandibular glands of stingless bees (Hymenoptera: Apidae): Chemical analysis of their contents and biological function in two species of Melipona. J. Chem. Ecol. 1983, 9, 1465–1472. [Google Scholar] [CrossRef]
- Fortunato, A.; Maile, R.; Turillazzi, S.; Morgan, E.D.; Moneti, G.; Jones, G.R.; Pieraccini, G. Defensive role of secretion of ectal mandibular glands of the wasp Polistes dominulus. J. Chem. Ecol. 2001, 27, 569–579. [Google Scholar] [CrossRef]
- Wilson, E.O. Chemical communication in the social insects. Science 1965, 149, 1064–1071. [Google Scholar] [CrossRef]
- Hölldobler, B.; Wilson, E.O. The Ants; Harvard University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Bradshaw, J.W.S.; Baker, R.; Howse, P.E. Multicomponent alarm pheromones of the weaver ant. Nature 1975, 258, 230–231. [Google Scholar] [CrossRef]
- Bradshaw, J.W.S.; Howse, P.E. Sociochemicals of Ants. In Chemical Ecology of Insects; Springer: Berlin/Heidelberg, Germany, 1984; pp. 429–473. [Google Scholar]
- Maccaro, J.J.; Whyte, B.A.; Tsutsui, N.D. The Ant Who Cried Wolf? Short-Term Repeated Exposure to Alarm Pheromone Reduces Behavioral Response in Argentine Ants. Insects 2020, 11, 871. [Google Scholar] [CrossRef]
- López-Incera, A.; Nouvian, M.; Ried, K.; Müller, T.; Briegel, H.J. Honeybee communication during collective defence is shaped by predation. BMC Biol. 2021, 19, 106. [Google Scholar] [CrossRef]
- Hu, L.; Balusu, R.R.; Zhang, W.-Q.; Ajayi, O.S.; Lu, Y.-Y.; Zeng, R.-S.; Fadamiro, H.Y.; Chen, L. Intra- and inter-specific variation in alarm pheromone produced by Solenopsis fire ants. Bull. Entomol. Res. 2018, 108, 667–673. [Google Scholar] [CrossRef]
- Núñez, J. Alarm Pheromone Induces Stress Analgesia via an Opioid System in the Honeybee. Physiol. Behav. 1997, 63, 75–80. [Google Scholar] [CrossRef]
- Alaux, C.; Robinson, G.E. Alarm Pheromone Induces Immediate–Early Gene Expression and Slow Behavioral Response in Honey Bees. J. Chem. Ecol. 2007, 33, 1346–1350. [Google Scholar] [CrossRef]
- Rogers, L.J.; Vallortigara, G. Complementary Specializations of the Left and Right Sides of the Honeybee Brain. Front. Psychol. 2019, 10, 280. [Google Scholar] [CrossRef] [PubMed]
- Rossi, N.; Baracchi, D.; Giurfa, M.; D’Ettorre, P. Pheromone-Induced Accuracy of Nestmate Recognition in Carpenter Ants: Simultaneous Decrease in Type I and Type II Errors. Am. Nat. 2019, 193, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Baracchi, D.; Cabirol, A.; Devaud, J.-M.; Haase, A.; D’Ettorre, P.; Giurfa, M. Pheromone components affect motivation and induce persistent modulation of associative learning and memory in honey bees. Commun. Biol. 2020, 3, 447. [Google Scholar] [CrossRef]
- Jernigan, C.M.; Birgiolas, J.; Mchugh, C.; Roubik, D.W.; Wcislo, W.T.; Smith, B.H. Colony-level non-associative plasticity of alarm responses in the stingless honey bee, Tetragonisca angustula. Behav. Ecol. Sociobiol. 2018, 72, 58. [Google Scholar] [CrossRef]
- London, K.B.; Jeanne, R.L. Effects of colony size and stage of development on defense response by the swarm-founding wasp Polybia occidentalis. Behav. Ecol. Sociobiol. 2003, 54, 539–546. [Google Scholar] [CrossRef]
- Sun, C.-X.; Li, Z.-X. Biosynthesis of aphid alarm pheromone is modulated in response to starvation stress under regulation by the insulin, glycolysis and isoprenoid pathways. J. Insect Physiol. 2021, 128, 104174. [Google Scholar] [CrossRef]
- Westwick, R.R.; Rittschof, C.C. Insects Provide Unique Systems to Investigate How Early-Life Experience Alters the Brain and Behavior. Front. Behav. Neurosci. 2021, 15, 61. [Google Scholar] [CrossRef]
- Breed, M.D.; Guzmán-Novoa, E.; Hunt, G.J. Defensive behaviour of Honeybees: Organization, Genetics, and Comparisons with Other Bees. Annu. Rev. Entomol. 2004, 49, 271–298. [Google Scholar] [CrossRef]
- Breed, M.D.; Rogers, K.B. The behavioral genetics of colony defense in honeybees: Genetic variability for guarding behavior. Behav. Genet. 1991, 21, 295–303. [Google Scholar] [CrossRef]
- Pokorny, T.; Sieber, L.-M.; Hofferberth, J.E.; Bernadou, A.; Ruther, J. Age-dependent release of and response to alarm pheromone in a ponerine ant. J. Exp. Biol. 2020, 223, jeb218040. [Google Scholar] [CrossRef]
- Steinbrecht, R.A. Structure and function of insect olfactory sensilla. In Proceedings of the Ciba Foundation Symposium 200-Olfaction in Mosquito-Host Interactions: Olfaction in Mosquito-Host Interactions: Ciba Foundation Symposium 200; John Wiley & Sons, Ltd.: Chichester, UK, 2007; pp. 158–183. [Google Scholar]
- Steinbrecht, R.A. Pore structures in insect olfactory sensilla: A review of data and concepts. Int. J. Insect Morphol. Embryol. 1997, 26, 229–245. [Google Scholar] [CrossRef]
- Keil, T.A. Comparative morphogenesis of sensilla: A review. Int. J. Insect Morphol. Embryol. 1997, 26, 151–160. [Google Scholar] [CrossRef]
- Grabe, V.; Sachse, S. Fundamental principles of the olfactory code. Biosystems 2018, 164, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Galizia, C.G. Olfactory coding in the insect brain: Data and conjectures. Eur. J. Neurosci. 2014, 39, 1784–1795. [Google Scholar] [CrossRef] [PubMed]
- Galizia, C.G.; Rössler, W. Parallel Olfactory Systems in Insects: Anatomy and Function. Annu. Rev. Entomol. 2010, 55, 399–420. [Google Scholar] [CrossRef]
- Kirschner, S.; Kleineidam, C.J.; Zube, C.; Rybak, J.; Grünewald, B.; Rössler, W. Dual olfactory pathway in the honeybee, Apis mellifera. J. Comp. Neurol. 2006, 499, 933–952. [Google Scholar] [CrossRef]
- Zube, C.; Kleineidam, C.J.; Kirschner, S.; Neef, J.; Rössler, W. Organization of the olfactory pathway and odor processing in the antennal lobe of the ant Camponotus floridanus. J. Comp. Neurol. 2008, 506, 425–441. [Google Scholar] [CrossRef]
- Kreissl, S.; Bicker, G. Histochemistry of acetylcholinesterase and immunocytochemistry of an acetylcholine receptor-like antigen in the brain of the honeybee. J. Comp. Neurol. 1989, 286, 71–84. [Google Scholar] [CrossRef]
- Schäfer, S.; Bicker, G. Distribution of GABA-like immunoreactivity in the brain of the honeybee. J. Comp. Neurol. 1986, 246, 287–300. [Google Scholar] [CrossRef]
- Galán, R.F.; Sachse, S.; Galizia, C.G.; Herz, A.V.M. Odor-Driven Attractor Dynamics in the Antennal Lobe Allow for Simple and Rapid Olfactory Pattern Classification. Neural Comput. 2004, 16, 999–1012. [Google Scholar] [CrossRef][Green Version]
- Sachse, S.; Beshel, J. The good, the bad, and the hungry: How the central brain codes odor valence to facilitate food approach in Drosophila. Curr. Opin. Neurobiol. 2016, 40, 53–58. [Google Scholar] [CrossRef]
- Sandoz, J.-C. Understanding the logics of pheromone processing in the honeybee brain: From labeled-lines to across-fiber patterns. Front. Behav. Neurosci. 2007, 1, 5. [Google Scholar] [CrossRef]
- Suh, G.S.B.; Wong, A.M.; Hergarden, A.C.; Wang, J.W.; Simon, A.F.; Benzer, S.; Axel, R.; Anderson, D.J. A single population of olfactory sensory neurons mediates an innate avoidance behaviour in Drosophila. Nature 2004, 431, 854–859. [Google Scholar] [CrossRef]
- Du, Y.; Chen, J. The Odorant Binding Protein, SiOBP5, Mediates Alarm Pheromone Olfactory Recognition in the Red Imported Fire Ant, Solenopsis invicta. Biomolecules 2021, 11, 1595. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, N.; Nishino, H.; Mizunami, M. Pheromone-sensitive glomeruli in the primary olfactory centre of ants. Proc. R. Soc. B Biol. Sci. 2006, 273, 2219–2225. [Google Scholar] [CrossRef]
- Keesey, I.W.; Hansson, B.S. The neuroethology of labeled lines in insect olfactory systems. In Insect Pheromone Biochemistry and Molecular Biology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 285–327. [Google Scholar]
- Galizia, C.; Menzel, R.; Hölldobler, B. Optical Imaging of Odor-Evoked Glomerular Activity Patterns in the Antennal Lobes of the Ant Camponotus rufipes. Naturwissenschaften 1999, 86, 533–537. [Google Scholar] [CrossRef] [PubMed]
- Galizia, C.G.; Sachse, S.; Rappert, A.; Menzel, R. The glomerular code for odor representation is species specific in the honeybee Apis mellifera. Nat. Neurosci. 1999, 2, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Joerges, J.; Küttner, A.; Galizia, C.G.; Menzel, R. Representations of odours and odour mixtures visualized in the honeybee brain. Nature 1997, 387, 285–288. [Google Scholar] [CrossRef]
- Yamagata, N.; Nishino, H.; Mizunami, M. Neural pathways for the processing of alarm pheromone in the ant brain. J. Comp. Neurol. 2007, 505, 424–442. [Google Scholar] [CrossRef]
- Mizunami, M.; Yamagata, N.; Nishino, H. Alarm pheromone processing in the ant brain: An evolutionary perspective. Front. Behav. Neurosci. 2010, 4, 28. [Google Scholar] [CrossRef]
- Mauelshagen, J. Neural correlates of olfactory learning paradigms in an identified neuron in the honeybee brain. J. Neurophysiol. 1993, 69, 609–625. [Google Scholar] [CrossRef] [PubMed]
- Sachse, S.; Rappert, A.; Galizia, C.G. The spatial representation of chemical structures in the antennal lobe of honeybees: Steps towards the olfactory code. Eur. J. Neurosci. 1999, 11, 3970–3982. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Sato, K.; Giurfa, M.; Zhang, S. Processing of sting pheromone and its components in the antennal lobe of the worker honeybee. J. Insect Physiol. 2008, 54, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Carcaud, J.; Giurfa, M.; Sandoz, J.-C. Differential Combinatorial Coding of Pheromones in Two Olfactory Subsystems of the Honey Bee Brain. J. Neurosci. 2015, 35, 4157–4167. [Google Scholar] [CrossRef]
- Kiyose, K.; Katsuki, M.; Suzaki, Y.; Okada, K.; Okada, Y. Octopaminergic system orchestrates combat and mating behaviors: A potential regulator of alternative male mating tactics in an armed beetle. J. Insect Physiol. 2021, 131, 104211. [Google Scholar] [CrossRef]
- Stevenson, P.A. Octopamine and Experience-Dependent Modulation of Aggression in Crickets. J. Neurosci. 2005, 25, 1431–1441. [Google Scholar] [CrossRef]
- Alekseyenko, O.V.; Chan, Y.-B.; Li, R.; Kravitz, E.A. Single dopaminergic neurons that modulate aggression in Drosophila. Proc. Natl. Acad. Sci. USA 2013, 110, 6151–6156. [Google Scholar] [CrossRef]
- Alekseyenko, O.V.; Chan, Y.-B.; de la Paz Fernandez, M.; Bülow, T.; Pankratz, M.J.; Kravitz, E.A. Single Serotonergic Neurons that Modulate Aggression in Drosophila. Curr. Biol. 2014, 24, 2700–2707. [Google Scholar] [CrossRef]
- Dierick, H.A.; Greenspan, R.J. Serotonin and neuropeptide F have opposite modulatory effects on fly aggression. Nat. Genet. 2007, 39, 678–682. [Google Scholar] [CrossRef]
- Galizia, C.G.; Kreissl, S. Neuropeptides in Honey Bees. In Honeybee Neurobiology and Behavior; Springer: Berlin/Heidelberg, Germany, 2012; pp. 211–226. [Google Scholar]
- Harrison, J.W.; Palmer, J.H.; Rittschof, C.C. Altering social cue perception impacts honey bee aggression with minimal impacts on aggression-related brain gene expression. Sci. Rep. 2019, 9, 14642. [Google Scholar] [CrossRef]
- Kravitz, E.A.; Huber, R. Aggression in invertebrates. Curr. Opin. Neurobiol. 2003, 13, 736–743. [Google Scholar] [CrossRef] [PubMed]
- Scheiner, R.; Baumann, A.; Blenau, W. Aminergic Control and Modulation of Honeybee Behaviour. Curr. Neuropharmacol. 2006, 4, 259–276. [Google Scholar] [CrossRef] [PubMed]
- Nouvian, M.; Mandal, S.; Jamme, C.; Claudianos, C.; D’Ettorre, P.; Reinhard, J.; Barron, A.B.; Giurfa, M. Cooperative defence operates by social modulation of biogenic amine levels in the honey bee brain. Proc. R. Soc. B Biol. Sci. 2018, 285, 20172653. [Google Scholar] [CrossRef] [PubMed]
- Homberg, U. Processing of antennal information in extrinsic mushroom body neurons of the bee brain. J. Comp. Physiol. A 1984, 154, 825–836. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kannan, K.; Galizia, C.G.; Nouvian, M. Olfactory Strategies in the Defensive Behaviour of Insects. Insects 2022, 13, 470. https://doi.org/10.3390/insects13050470
Kannan K, Galizia CG, Nouvian M. Olfactory Strategies in the Defensive Behaviour of Insects. Insects. 2022; 13(5):470. https://doi.org/10.3390/insects13050470
Chicago/Turabian StyleKannan, Kavitha, C. Giovanni Galizia, and Morgane Nouvian. 2022. "Olfactory Strategies in the Defensive Behaviour of Insects" Insects 13, no. 5: 470. https://doi.org/10.3390/insects13050470
APA StyleKannan, K., Galizia, C. G., & Nouvian, M. (2022). Olfactory Strategies in the Defensive Behaviour of Insects. Insects, 13(5), 470. https://doi.org/10.3390/insects13050470