
Effects of Repeated Short-Term Heat Exposure on Life History Traits of Colorado Potato Beetle


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects Rearing
2.2. Treatments and Data Collection
2.3. Data Analyses
3. Results
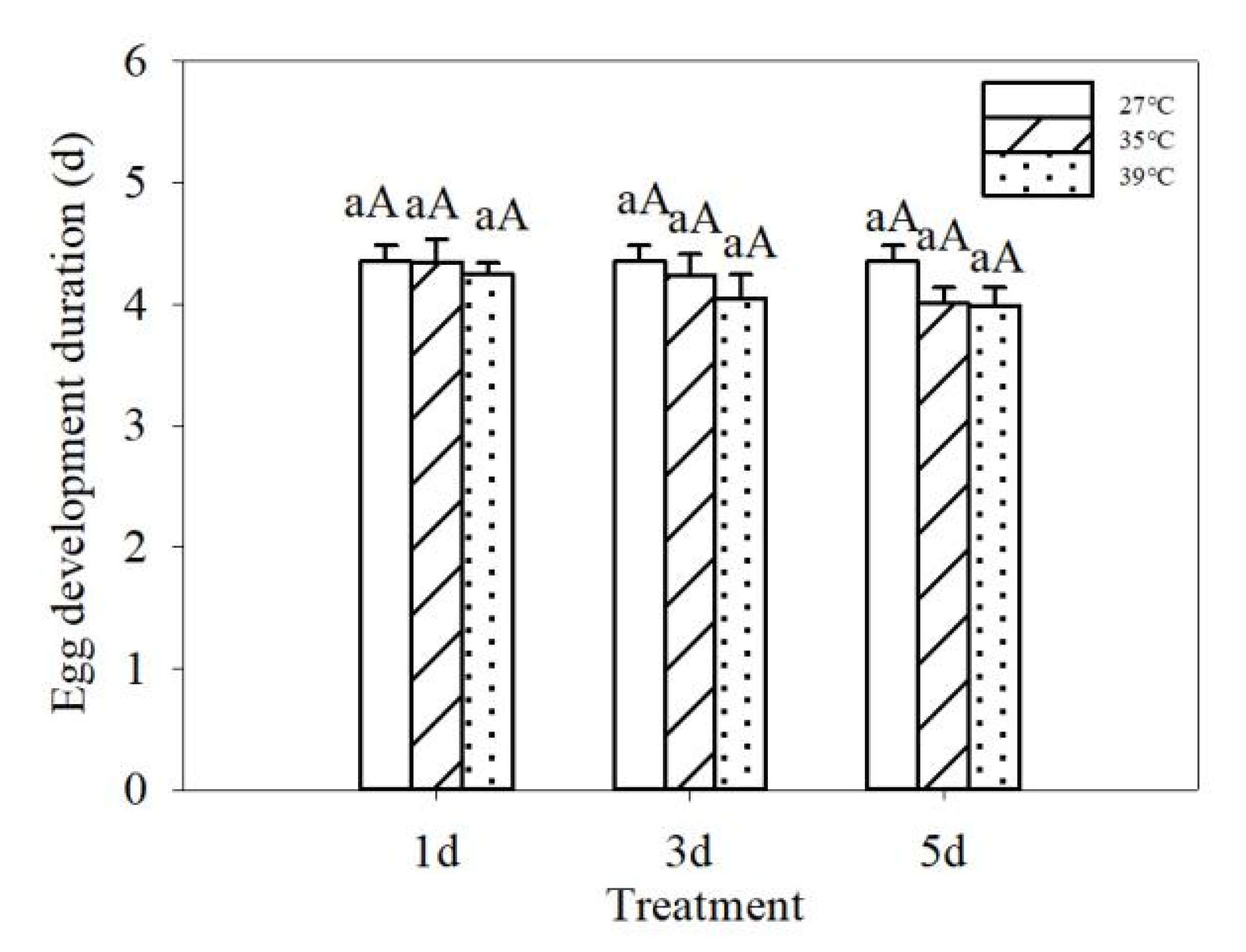
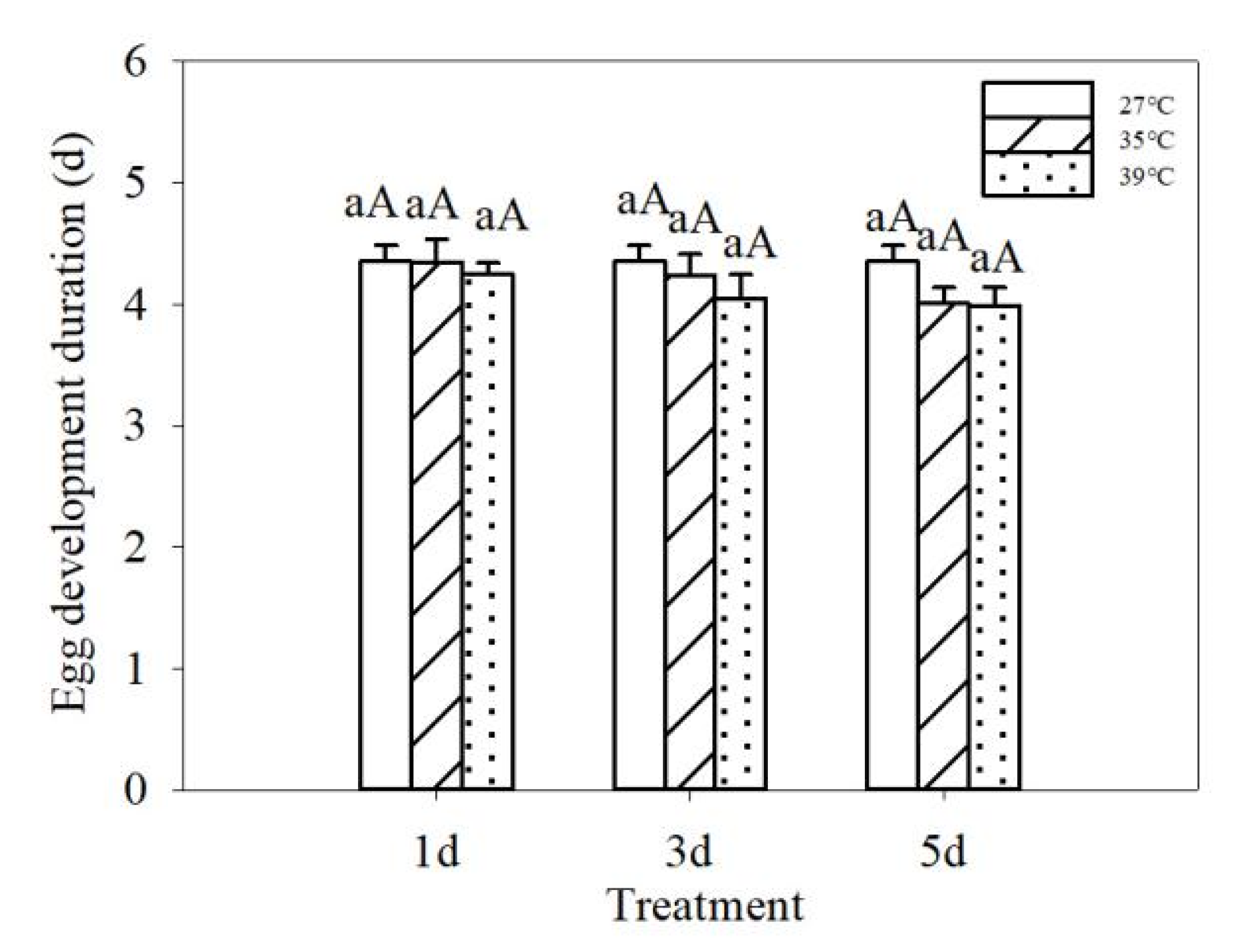
3.1. Effects of Repeated Short-Term Heat Exposure on the Hatching of Colorado Potato Beetles Eggs
3.2. Effects of Repeated Short-Term Heat Exposure on the Reproduction of Adults Colorado Potato Beetles
3.3. Effect of Repeated Short-Term Heat Exposure on the Demographic Parameters of Adults Colorado Potato Beetle
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- IPCC. Climate Change 2013: The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: New York, NY, USA, 2013; pp. 123–147. [Google Scholar]
- Barriopedro, D.; Fischer, E.M.; Luterbacher, J.; Trigo, R.M.; García, H.R. The hot summer of 2010: Redrawing the temperature record map of Europe. Science 2011, 332, 220–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.; Li, S.; Zhang, B.; Li, H.G.; Wan, F.H.; Zheng, C.Y. Effects of extreme high temperature on survival rate, reproduction, trehalose and sorbitol of Frankliniella occidentalis. Sci. Agric. Sin. 2016, 49, 2310–2321. [Google Scholar]
- Terada, K.; Matsumura, K.; Miyatake, T. Effects of temperature during successive generations on life-history traits in a seed beetle Callosobruchus chinensis (Chrysomelidae: Coleoptera). Appl. Entomol. Zool. 2019, 54, 459. [Google Scholar] [CrossRef]
- Wang, X.D.; Ji, S.X.; Shen, X.N.; Liu, W.X.; Wan, F.H.; Lv, Z.C. Research advance of epigenetic mechanism in temperature stress. Chin. J. Biol. Control 2021, 37, 598–608. [Google Scholar]
- Yang, Y.T.; Liang, L.; Shi, C.H.; Xie, W.; Zhang, Y.J. Age-stage, two-sex life table of laboratory populations of Bradisia odoriphaga under different temperatures. Plant Prot. 2019, 45, 51–57. [Google Scholar]
- Gao, G.Z.; Lv, Z.Z.; Xia, D.P.; Sun, P. Effects of pattern and timing of high temperature exposure on the mortality and fecundity of Aphis gossypii Glover on cotton. Acta Ecol. Sin. 2012, 32, 7568–7575. [Google Scholar]
- Zhang, H.; Lin, J.T. Responses of insects to global warming. J. Environ. Entomol. 2015, 37, 1280–1286. [Google Scholar]
- Giordanengo, P.; Vincent, C.; Alyokhin, A. Insect Pests of Potato: Global Perspectives on Biology and Management; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Hiiesaar, K.; Karise, R.; Williams, I.H.; Luik, A.; Metspalu, L.; Jõgar, K.; Eremeev, V.; Ploomi, A.; Kruus, E.; Mänd, M. Cold tolerance of Colorado potato beetle (Leptinotarsa decemlineata Say) adults and eggs. Zemdirbyste-Agriculture 2014, 101, 431–436. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Hawthorne, D.; Qin, Y.; Pan, X.; Li, Z.; Zhu, S. Impact of climate and host availability on future distribution of Colorado potato beetle. Sci. Rep. 2017, 7, 4489. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.J.; You, Q.L.; Wang, P.L.; Kong, L. Analysis on heat waves variation features in China during 1961–2014. J. Meteorol. Res. 2018, 38, 28–36. [Google Scholar]
- Han, Z.L.; Tan, X.L.; Chen, J.L. Effect of environmental temperature variations on flight and locomotory behavior of Harmonia axyridis (Coleoptera: Coccinellidae). Chin. J. Biol. Control 2017, 33, 433–441. [Google Scholar]
- Li, G.J.; Benoit, F.; Ceustermans, N. Influence of day and night temperature on the growth, development and yield of greenhouse sweet pepper. J. Zhejiang Univ.-Agric. Life Sci. 2004, 30, 19–23. [Google Scholar]
- Du, P.H.; Schlemmer, M.L.; Van, D.B.J. The effect of temperature on the development of Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2020, 11, 228. [Google Scholar]
- Milosavljevic, I.; McCalla, K.A.; Morgan, D.; Hoddle, M.S. The effects of constant and fluctuating temperatures on development of Diaphorina citri (Hemiptera: Liviidae), the Asian Citrus Psyllid. J. Econ. Entomol. 2020, 113, 633–645. [Google Scholar] [CrossRef]
- Gu, X.P.; Huang, Y.Y.; Zhang, J.Y.; Zhang, X.M.; Chen, G.H. Effects of short-term high-temperature stress on growth, development and reproduction of melon fly. J. Environ. Entomol. 2020, 42, 391–399. [Google Scholar]
- Zhang, Y.H.; Zhang, Z.; He, J.; Tuerxun, A.; Cheng, D.F. Cold hardiness of natural populations of the Colorado potato beetle, Leptinotarsa decemlineata. Plant Prot. 2012, 38, 64–67. [Google Scholar]
- Ferro, D.N.; Logan, J.A.; Voss, R.H.; Elkinton, J.S. Colorado potato beetle (Coleoptera: Chrysomelidae) temperature-dependent growth and feeding rates. Environ. Entomol. 1985, 14, 343–348. [Google Scholar] [CrossRef]
- He, J.; Ding, X.H.; Yue, R.Q.; Li, X.H.; Tuerxun, A.; Guo, W.C. Preliminary study on the influence of temperature and water content of soil on relieving the Colorado potato beetles’ hibernation. Xinjiang Agric. Sci. 2014, 51, 1306–1311. [Google Scholar]
- Li, C.; Cheng, D.F.; Liu, H.; Zhang, Y.H. Effects of temperature on the distribution of the Colorado potato beetle (Leptinotarsa decemlineata)—Effect of high temperature on its emergence in Turpan, Xinjiang. Sci. Agric. Sin. 2013, 46, 737–744. [Google Scholar]
- Zhang, Z.; Li, C.; Cheng, D.F.; Zhang, Y.H.; Sun, J.R.; Wei, Z. Preliminary study on the lethal effect of high temperature on Colorado potato beetle. In Public Plant Protection and Green Prevention and Control, Proceedings of the 2010 Annual Academic Meeting of Chinese Society of Plant Protection, Hebi, Henan, China, 28 October 2010; China Academic Journal Electronic Publishing House: Beijing, China, 2010. [Google Scholar]
- Li, C.; Liu, H.; Huang, F.N.; Cheng, D.F.; Wang, J.J.; Zhang, Y.H.; Sun, J.R.; Guo, W.C. Effect of temperature on the occurrence and distribution of Colorado potato beetle (Coleoptera: Chrysomelidae) in China. Environ. Entomol. 2014, 43, 511–519. [Google Scholar] [CrossRef]
- Wang, L.; Ma, C.S. Impacts of periodic repeated heat events on ecological performance in insects. Chin. J. Appl. Entomol. 2013, 50, 1499–1508. [Google Scholar]
- Liao, J.H.; Liu, J.; Guan, Z.J.; Li, C. Duration of low temperature exposure affects egg hatching of the Colorado potato beetle and emergence of overwintering adults. Insects 2021, 12, 609. [Google Scholar] [CrossRef] [PubMed]
- Logan, P.A.; Casagrande, R.A.; Faubert, H.H.; Drummond, F.A. Temperature-dependent development and feeding of immature Colorado potato beetles, Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae). Environ. Entomol. 1985, 14, 275–283. [Google Scholar] [CrossRef]
- Zheng, H.Y.; Gao, Y.L.; Zhang, F.M.; Yang, C.X.; Jiang, J.; Zhu, X.; Zhang, Y.H.; Li, X.R. Cloning of heat shock protein gene Ld-hsp70 in Leptinotarsa decemlineata and its expression characteristics under temperature stress. Sci. Agric. Sin. 2021, 54, 1163–1175. [Google Scholar]
- Atashi, N.; Shishehbor, P.; Seraj, A.A.; Rasekh, A.; Hemmati, S.A.; Riddick, E.W. Effects of Helicoverpa armigera egg age on development, reproduction, and life table parameters of Trichogramma euproctidis. Insects 2021, 12, 569. [Google Scholar] [CrossRef]
- Ahn, J.J.; Cho, J.R.; Kim, J.H.; Seo, B.Y. Thermal effects on the population parameters and growth of Acyrthosiphon pisum (Harris) (Hemiptera: Aphididae). Insects 2020, 11, 481. [Google Scholar] [CrossRef]
- Ying, C.J.; Guo, H.L.; Cheng, G.Z.; Shu, Y.Y. Influence of temperature on the development and reproduction of Cinara cedri (Hemiptera: Aphidoidea: Lachninae). Bull. Entomol. Res. 2021, 111, 579–584. [Google Scholar]
- Zhou, Z.X.; Luo, J.C.; Lv, H.P.; Guo, W.C. Influence of temperature on development and reproduction of experimental populations of the Colorado potato beetle, Leptinotarsa decemlineata(Say) (Coleoptera: Chrysomelidae). Acta Ecol. Sin. 2010, 53, 926–931. [Google Scholar]
- Guo, W.C.; Tuerxun, A.; Cheng, D.F.; Tan, W.Z.; Zhang, Z.K.; Li, G.Q.; Jiang, W.H.; Deng, J.Y.; Wu, J.H.; Deng, C.S.; et al. Main progress on biology &ecology of Colorado potato beetle and counter measures of its monitoring and controlling in China. Plant Prot. 2014, 40, 1–11. [Google Scholar]
- Du, J.L.; Wu, G.D.; Lv, Y.; Liu, C.Z. Population parameters of two pea aphid (Acyrthosiphon pium) color morphs at different temperatures. Acta Prataculturae Sin. 2015, 24, 91–99. [Google Scholar]
- Cui, J.; Qiao, F.; Hu, Y.L.; Tian, X.Y.; Zhao, L.N.; Shi, S.S. Antioxidant responses of Megacopta cribraria (Hemiptera: Plataspidae) adults exposed to high temperature stress. J. Environ. Entomol. 2021, 43, 1244–1249. [Google Scholar]
- Wei, C.P.; Jiang, J.; Li, R.X.; Zhang, Y.H.; Zhang, F.M.; Deng, C.F.; Guo, W.C.; Liu, H. Cloning and sequence analysis of heat shock protein gene Ld-HSP60 in Leptinotarsa decemlineata (Coleoptera: Chrysomelidae) and its expression under temperature stress. Acta Ecol. Sin. 2017, 60, 523–532. [Google Scholar]
- Zhou, Z.S.; Guo, J.Y.; Min, L.; Wan, F.H. Effect of short-term high temperature stress on the development and fecundity of Ophraella communa (Coleoptera: Chrysomelidae). Biocontrol 2011, 21, 809–819. [Google Scholar] [CrossRef]
- Liang, F.; Wang, Z.; He, K.; Bai, S.; Zhang, T. Effects of brief exposure to high temperatures on the survival and fecundity of Athetis lepigone (Lepidoptera: Noctuidae). J. Therm. Biol. 2021, 100, 103066. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.J.; Liu, C.; Zhou, Z.X. A comparative study of development and reproduction of the Colorado potato beetle, Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae), on different host plants. Acta Ecol. Sin. 2012, 55, 84–90. [Google Scholar]
- Xu, Y.M.; Guo, W.C.; Xie, X.W.; Xia, Z.H.; Xu, J.J.; Zhang, D.M.; He, J.; Fu, W.J.; Ban, X.L. Response of Colorado potato beetle quantity in Xinjiang under different variety and fertilizer. Acta Agric. Boreali-Occident. Sin. 2011, 20, 179–185. [Google Scholar]
- Tian, T.; Zhang, Y.; Liu, T.X.; Zhang, S.Z. Effects of periodically repeated high-temperature exposure on the immediate and subsequent fitness of different developmental stages of Propylaea japonica. Pest. Manag. Sci. 2022, 78, 1649–1656. [Google Scholar] [CrossRef]
- Cao, Y.; Li, C.; Yang, W.J.; Meng, Y.L.; Wang, L.J.; Shang, B.Z.; Gao, Y.L. Effects of temperature on the development and reproduction of Thrips hawaiiensis (Thysanoptera: Thripidae). J. Econ. Entomol. 2018, 111, 755–760. [Google Scholar] [CrossRef]
- Dumas, P.; Morin, M.D.; Boquel, S.; Moffat, C.E.; Morin, P. Expression status of heat shock proteins in response to cold, heat, or insecticide exposure in the Colorado potato beetle Leptinotarsa decemlineata. Cell Stress Chaperon. 2019, 24, 539–547. [Google Scholar] [CrossRef]
- Grafius, E. Effects of temperature on pyrethroid toxicity to Colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1986, 79, 588–591. [Google Scholar] [CrossRef]
- Li, M.; Li, X.; Lü, J.; Huo, M. The effect of acclimation on heat tolerance of Lasioderma serricorne (Fabricius) (Coleoptera: Anobiidae). J. Therm. Biol. 2018, 71, 153–157. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Independent Variable | Egg Development Duration | Hatching Rate | ||||
|---|---|---|---|---|---|---|
| Degree Freedom | F | p | Degree Freedom | Wald χ2 | p | |
| Treatment temperature | 2 | 1.278 | 0.287 | 2 | 192.883 | 0.000 |
| Treatment days | 2 | 0.714 | 0.494 | 2 | 3.282 | 0.194 |
| Treatment temperature × Treatment days | 4 | 0.223 | 0.924 | 4 | 1.652 | 0.799 |
| Treatment Temperatures | Hatching Rate (%) |
|---|---|
| 27 °C (CK) | 91.00 ± 1.20 a |
| 35 °C | 63.00 ± 1.90 b |
| 39 °C | 52.00 ± 1.80 c |
| Independent Variable | Pre-Oviposition Duration | Fecundity | Oviposition Periods | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Degree Freedom | F | p | Degree Freedom | F | p | Degree Freedom | F | p | |
| Treatment temperature | 2 | 0.162 | 0.852 | 2 | 171.889 | 0.000 | 2 | 59.377 | 0.000 |
| Treatment days | 2 | 0.790 | 0.471 | 2 | 1.847 | 0.190 | 2 | 3.497 | 0.055 |
| Treatment temperature × Treatment days | 4 | 0.473 | 0.755 | 4 | 0.964 | 0.454 | 4 | 1.045 | 0.415 |
| Treatment Temperature | Pre-Oviposition Duration | Fecundity | Oviposition Periods |
|---|---|---|---|
| 27 °C (CK) | 10.92 ± 0.77 a | 315.96 ± 4.44 a | 14.01 ± 0.41 a |
| 35 °C | 10.80 ± 0.98 a | 134.10 ± 16.04 b | 8.62 ± 0.95 b |
| 39 °C | 11.69 ± 0.98 a | 50.25 ± 4.08 c | 3.90 ± 0.49 c |
| Independent Variable | Intrinsic Rate of Increase rm | Finite Rate of Increase λ | Net Reproductive Rate R0 | Mean Generation Time T | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Degree Freedom | F | p | Degree Freedom | F | p | Degree Freedom | F | p | Degree Freedom | F | p | |
| Treatment temperature | 2 | 119.392 | 0.000 | 2 | 114.602 | 0.000 | 2 | 175.484 | 0.000 | 2 | 3.118 | 0.072 |
| Treatment days | 2 | 1.045 | 0.375 | 2 | 0.984 | 0.395 | 2 | 1.338 | 0.290 | 2 | 0.115 | 0.892 |
| Treatment temperature × Treatment days | 4 | 0.304 | 0.871 | 4 | 0.289 | 0.881 | 4 | 0.915 | 0.479 | 4 | 0.081 | 0.987 |
| Treatment Temperature | Intrinsic Rate of Increase rm | Finite Rate of Increase λ | Net Reproductive Rate R0 | Mean Generation Time T |
|---|---|---|---|---|
| 27 °C (CK) | 0.1566 ± 0.0027 a | 1.1695 ± 0.0032 a | 160.1611 ± 2.6533 a | 32.5043 ± 0.6518 a |
| 35 °C | 0.1332 ± 0.0019 b | 1.1425 ± 0.0021 b | 68.4073 ± 7.8663 b | 31.3980 ± 0.2812 a |
| 39 °C | 0.1028 ± 0.0015 c | 1.1083 ± 0.0017 c | 23.5488 ± 1.3147 c | 30.6224 ± 0.1611 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, J.; Liu, J.; Li, C. Effects of Repeated Short-Term Heat Exposure on Life History Traits of Colorado Potato Beetle. Insects 2022, 13, 455. https://doi.org/10.3390/insects13050455
Liao J, Liu J, Li C. Effects of Repeated Short-Term Heat Exposure on Life History Traits of Colorado Potato Beetle. Insects. 2022; 13(5):455. https://doi.org/10.3390/insects13050455
Chicago/Turabian StyleLiao, Jianghua, Juan Liu, and Chao Li. 2022. "Effects of Repeated Short-Term Heat Exposure on Life History Traits of Colorado Potato Beetle" Insects 13, no. 5: 455. https://doi.org/10.3390/insects13050455
APA StyleLiao, J., Liu, J., & Li, C. (2022). Effects of Repeated Short-Term Heat Exposure on Life History Traits of Colorado Potato Beetle. Insects, 13(5), 455. https://doi.org/10.3390/insects13050455