Establishment, Genetic Diversity, and Habitat Suitability of Aedes albopictus Populations from Ecuador

 ,
,  , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction, Amplification, and Sequencing of the COI Gene
2.3. Phylogenetic Analyses
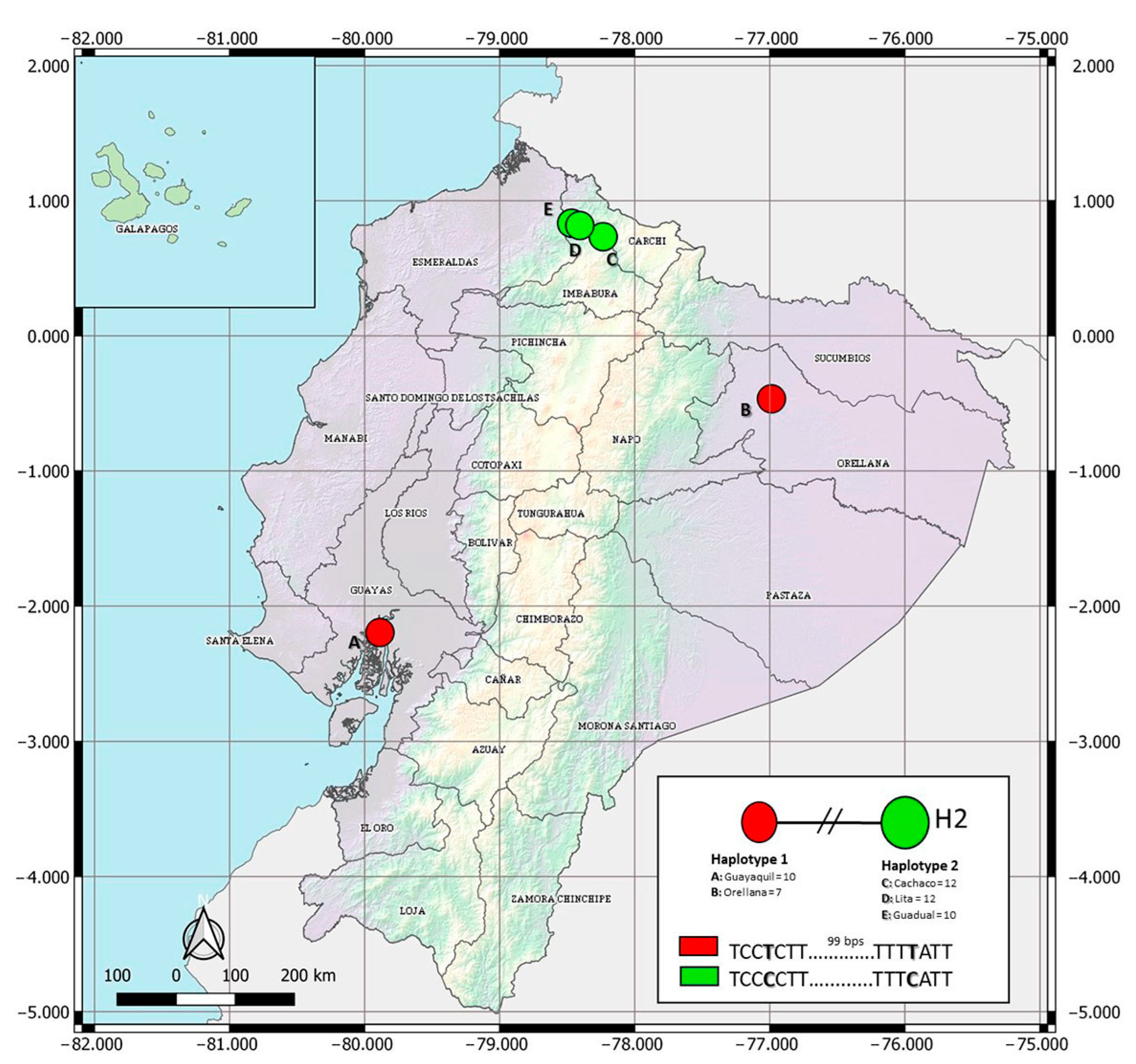
2.3.1. Haplotypes Network
2.3.2. Phylogenetic Tree
2.4. Genetic Diversity and Population Structure
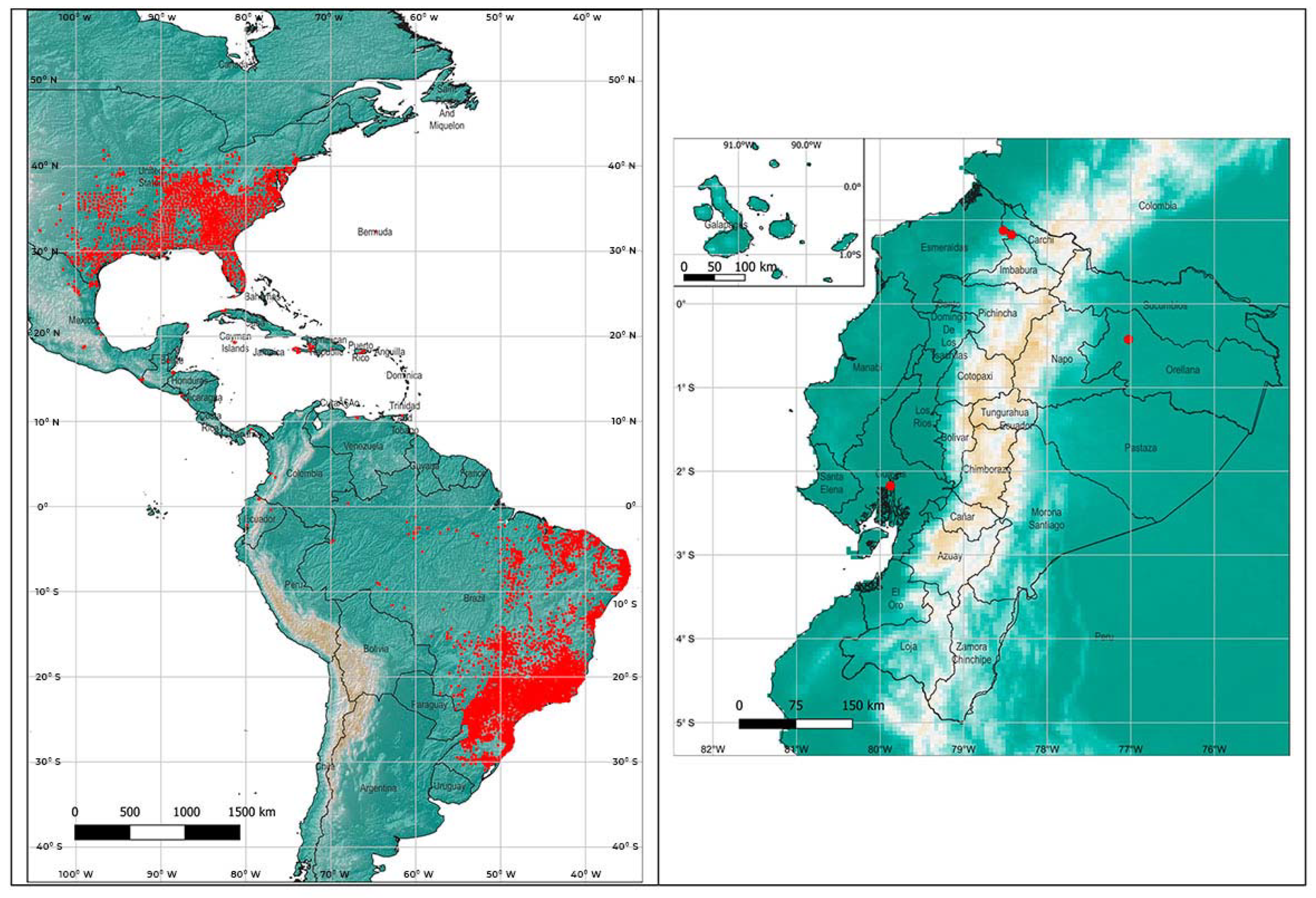
2.5. Maximum Entropy Model of the Asian Tiger Mosquito in Ecuador
3. Results
3.1. Phylogenetic Analyses
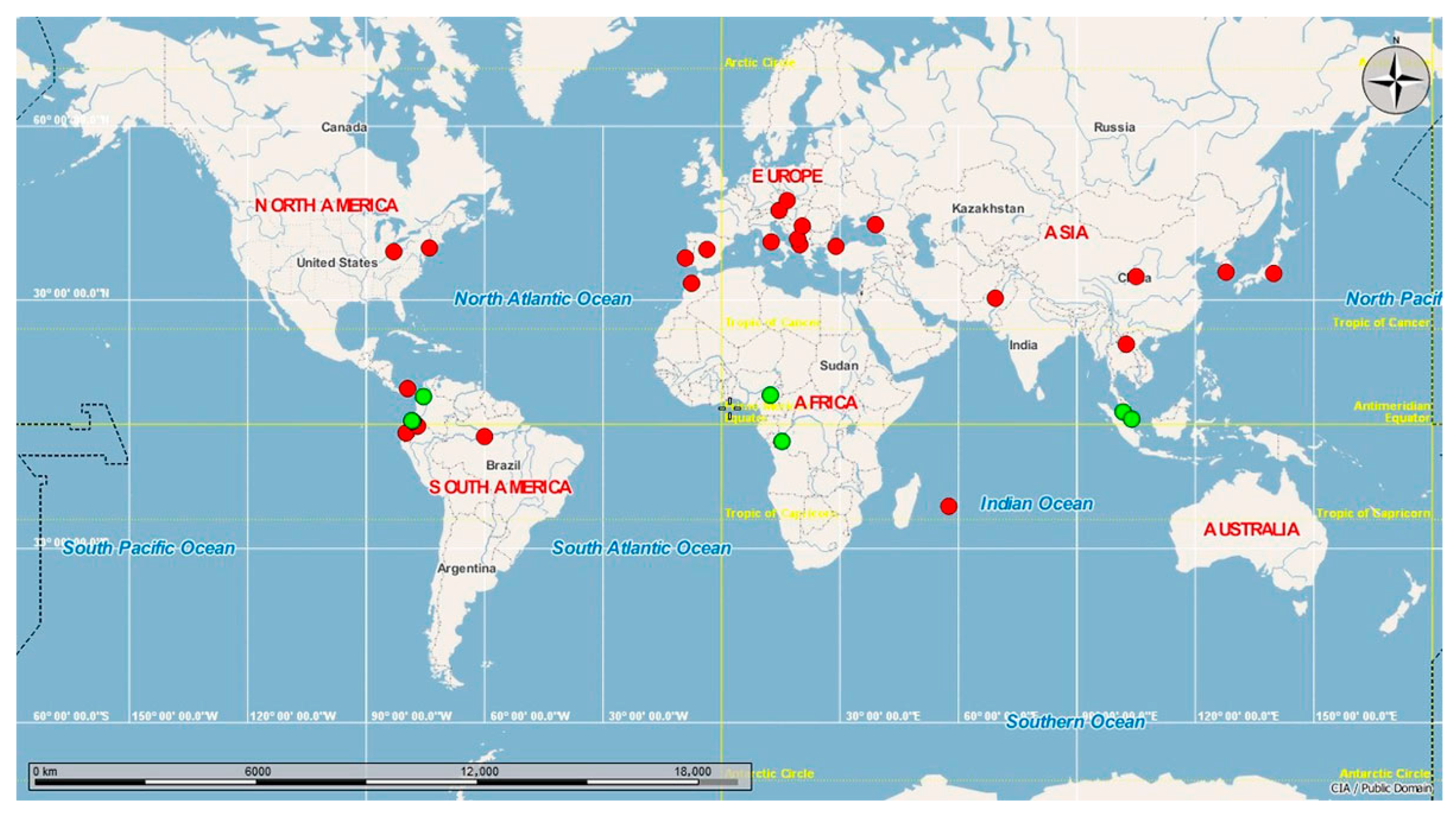
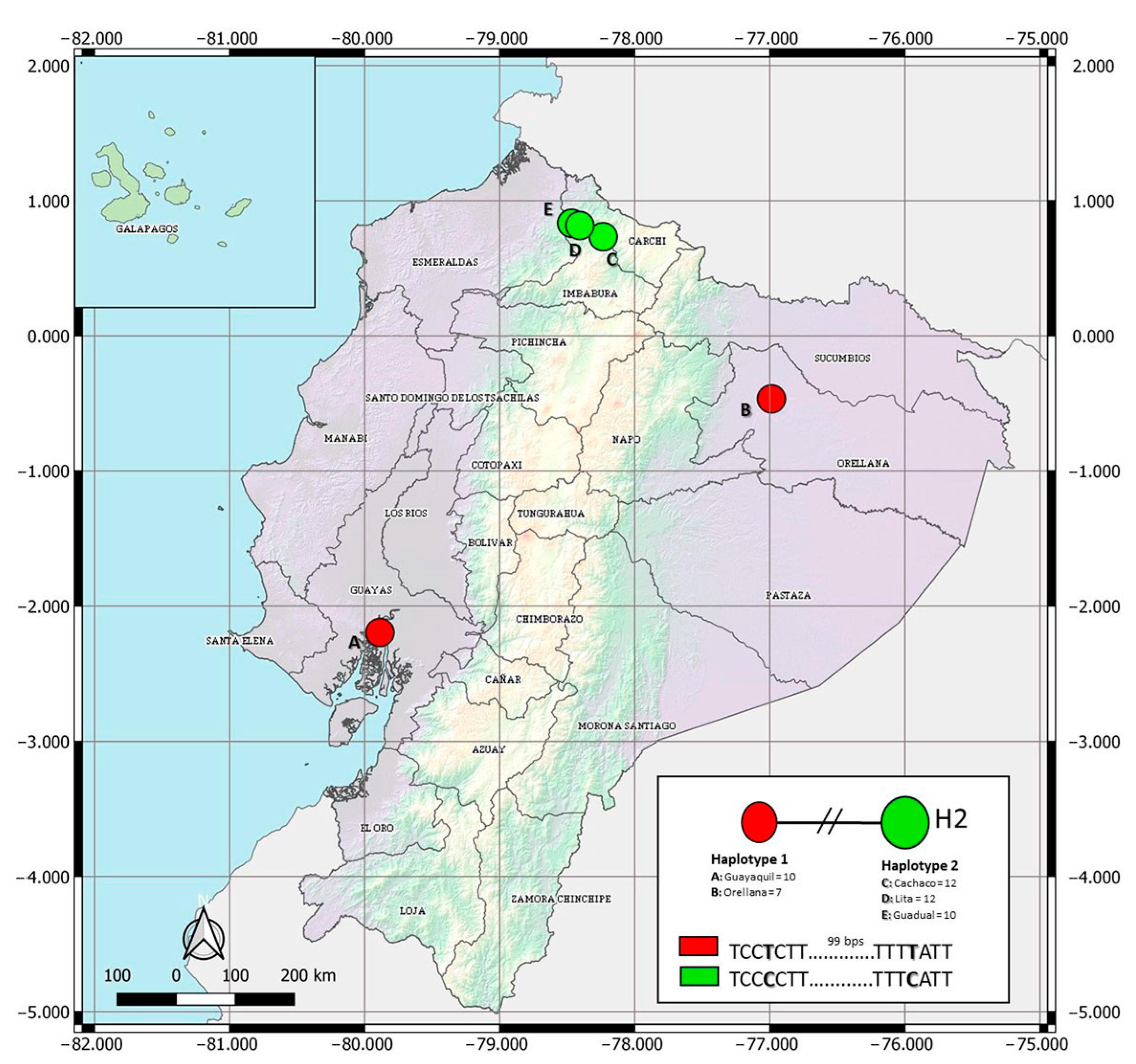
3.1.1. Haplotypes Network
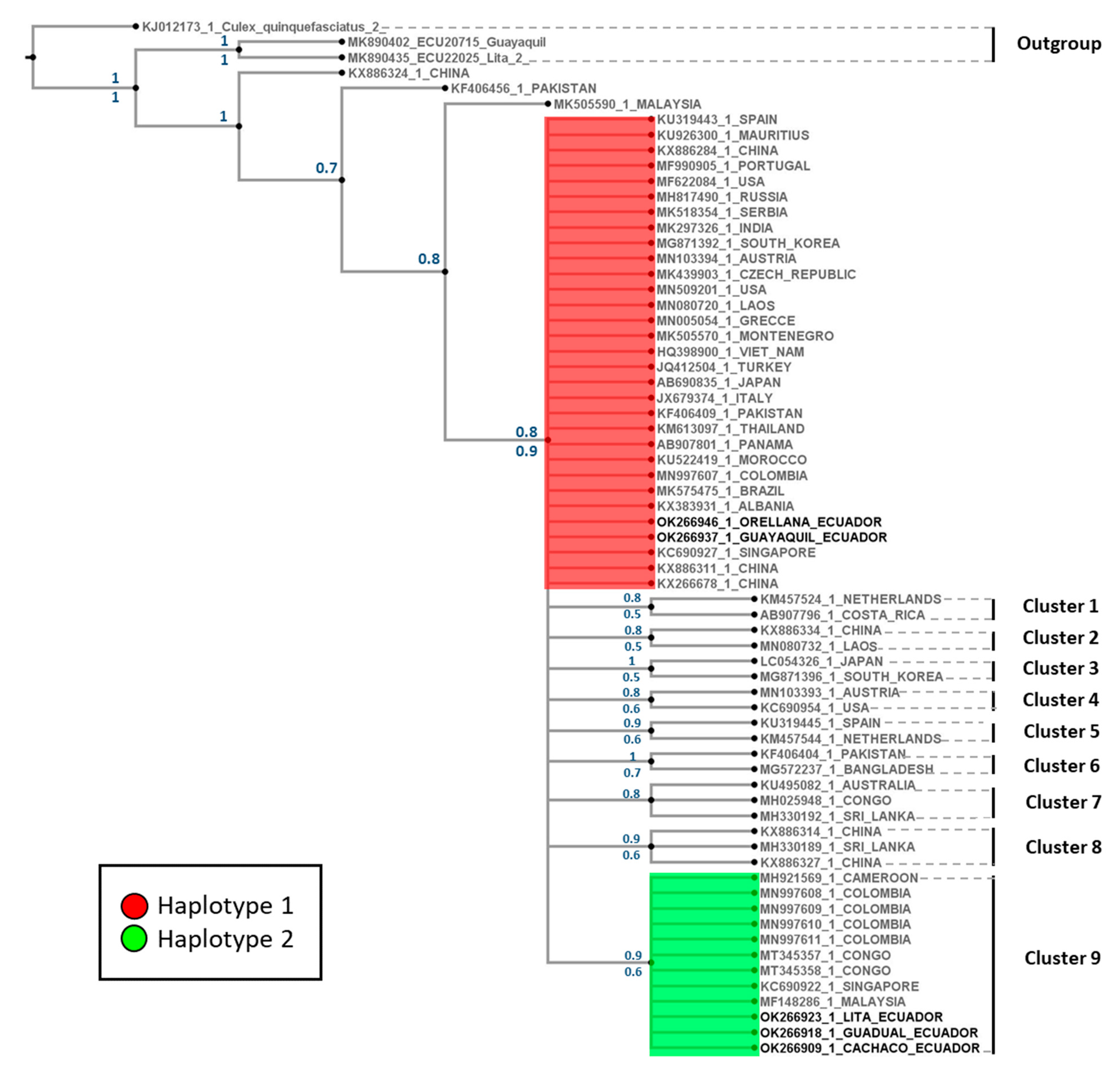
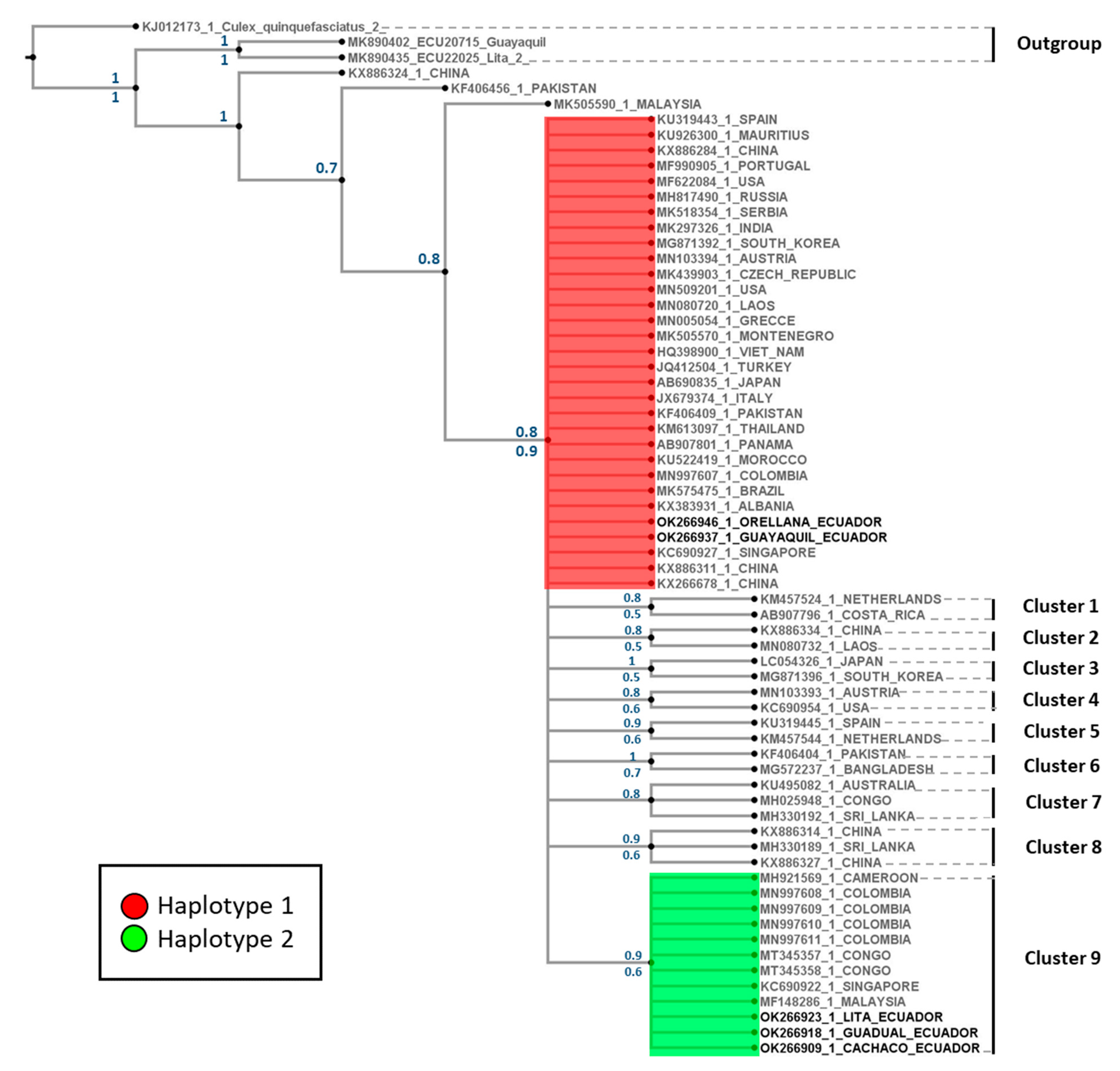
3.1.2. Phylogenetic Tree
3.2. Genetic Diversity and Population Structure
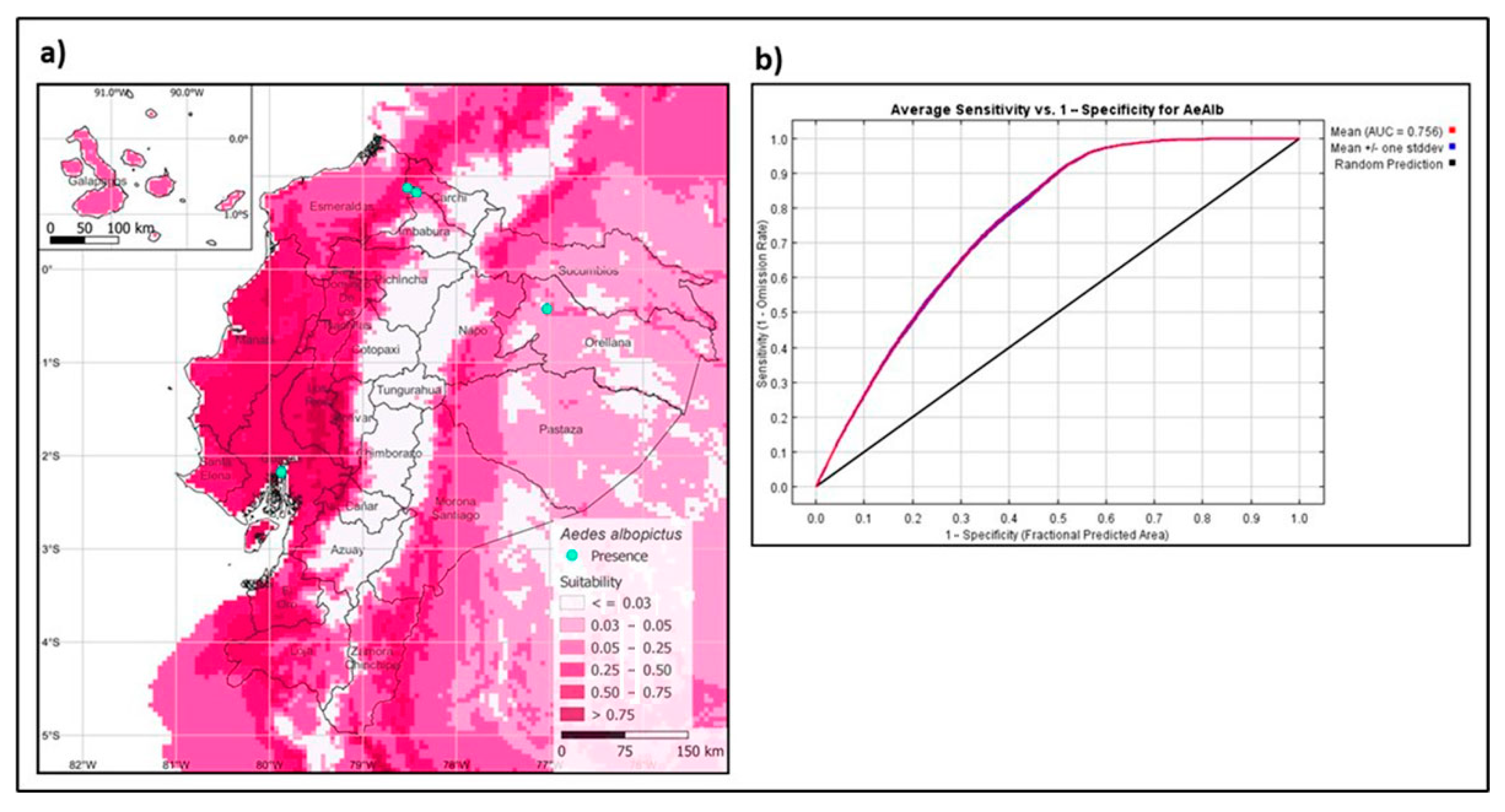
3.3. Maximum Entropy Model of the Asian Tiger Mosquito in Ecuador
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Medlock, J.M.; Hansford, K.M.; Schaffner, F.; Versteirt, V.; Hendrickx, G.; Zeller, H.; Van Bortel, W. A review of the invasive mosquitoes in Europe: Ecology, public health risks, and control options. Vector Borne Zoonotic Dis. 2012, 12, 435–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholte, E.J.; Schaffner, F. Waiting for the tiger: Establishment and spread of the Asian tiger mosquito in Europe. In Emerging Pests and Vector-Borne Diseases in Europe; Wageningen Academic Publishers: Wageningen, The Netherlands, 2007. [Google Scholar]
- Simberloff, D.W.; van der Weijden, R.; Leewis, P. Bol, Biological Globalisation. Bio-invasions and their impacts on nature, the economy, and public health. Biol. Invasions 2009, 11, 771–772. [Google Scholar] [CrossRef]
- Wong, P.-S.J.; Li, M.I.; Chong, C.-S.; Ng, L.-C.; Tan, C.-H. Aedes (Stegomyia) albopictus (Skuse): A potential vector of Zika virus in Singapore. PLoS Negl. Trop. Dis. 2013, 7, e2348. [Google Scholar] [CrossRef] [PubMed]
- Amraoui, F.; Vazeille, M.; Failloux, A.B. French Aedes albopictus are able to transmit yellow fever virus. Eurosurveillance 2016, 21, 30361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilder-Smith, A.; Gubler, D.J.; Weaver, S.C.; Monath, T.P.; Heymann, D.L.; Scott, T.W. Epidemic arboviral diseases: Priorities for research and public health. Lancet Infect. Dis. 2017, 17, e101–e106. [Google Scholar] [CrossRef] [Green Version]
- Ponce, P.; Morales, D.; Argoti, A.; Cevallos, V.E. First report of Aedes (Stegomyia) albopictus (Skuse) (Diptera: Culicidae), the asian tiger mosquito, in Ecuador. J. Med. Entomol. 2018, 55, 248–249. [Google Scholar] [CrossRef] [Green Version]
- Paduan, K.D.S.; Ribolla, P.E.M. Mitochondrial DNA polymorphism and heteroplasmy in populations of Aedes aegypti in Brazil. J. Med. Entomol. 2008, 45, 59–67. [Google Scholar] [CrossRef]
- Girard, E.B.; Langerak, A.; Jompa, J.; Wangensteen, O.S.; Macher, J.-N.; Renema, W. Mitochondrial cytochrome oxidase subunit 1: A Promising molecular marker for species identification in foraminifera. Front. Mar. Sci. 2022, 9, 809659. [Google Scholar] [CrossRef]
- Antich, A.; Palacin, C.; Wangensteen, O.S.; Turon, X. To denoise or to cluster, that is not the question: Optimizing pipelines for COI metabarcoding and metaphylogeography. BMC Bioinform. 2021, 22, 177. [Google Scholar] [CrossRef]
- Bracco, J.E.; Capurro, M.L.; Lourenço-de-Oliveira, R.; Sallum, M.A.M. Genetic variability of Aedes aegypti in the Americas using a mitochondrial gene: Evidence of multiple introductions. Mem. Inst. Oswaldo Cruz 2007, 102, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Fernando, H.S.D.; Hapugoda, M.; Perera, R.; Black Iv, W.C.; De Silva, B.G.D.N.K. Mitochondrial metabolic genes provide phylogeographic relationships of global collections of Aedes aegypti (Diptera: Culicidae). PLoS ONE 2020, 15, e0235430. [Google Scholar] [CrossRef] [PubMed]
- Avise, J.C.; Arnold, J.; Ball, R.M.; Bermingham, E.; Lamb, T.; Neigel, J.E.; Reeb, C.A.; Saunders, N.C. Intraspecific phylogeography: The mitochondrial DNA bridge between population genetics and systematics. Annu. Rev. Ecol. Syst. 1987, 18, 489–522. [Google Scholar] [CrossRef]
- Kang, S.; Jung, J.; Kim, W. Population genetic structure of the malaria vector Anopheles sinensis (Diptera: Culicidae) sensu stricto and evidence for possible introgression in the Republic of Korea. J. Med. Entomol. 2015, 52, 1270–1281. [Google Scholar] [CrossRef] [PubMed]
- Im, J.H.; Kim, T.-S.; Chung, M.-H.; Baek, J.H.; Kwon, H.Y.; Lee, J.-S. Current status and a perspective of mosquito-borne diseases in the Republic of Korea. Vector Borne Zoonotic Dis. 2021, 21, 69–77. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome C oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Sánchez-DelBarrio, J.C.; Messeguer, X.; Rozas, R. DnaSP, DNA polymorphism analyses by the coalescent and other methods. Bioinformatics 2003, 19, 2496–2497. [Google Scholar] [CrossRef]
- QGIS Development Team. QGIS Geographic Information System 2021. Available online: http://qgis.osgeo.org (accessed on 10 February 2022).
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Cummings, M.P. PAUP* (Phylogenetic Analysis Using Parsimony (and Other Methods)). In Dictionary of Bioinformatics and Computational Biology; John Wiley & Sons: Chichester, UK, 2004; Available online: https://onlinelibrary.wiley.com/doi/abs/10.1002/9780471650126.dob0522.pub2 (accessed on 1 March 2022).
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Muñoz-Tobar, S.I.; Caterino, M.S. The role of dispersal for shaping phylogeographical structure of flightless beetles from the Andes. PeerJ 2019, 7, e7226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckalew, J.O.; James, M.; Scott, L.; Reed, P. Water Resources Assessment of Ecuador; US Army Corps of Engineers: Mobile, AL, USA, 1988.
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. Maxent Software for Modeling Species Niches and Distributions, Version 3.4.1. 2017. Available online: https://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 27 February 2022).
- Org, O.G. GBIF Occurrence Download; GBIF: Copenhagen, Denmark, 2015; Available online: https://www.gbif.org/occurrence/download/0002031-170925132151566 (accessed on 10 February 2022).
- Futami, K.; Valderrama, A.; Baldi, M.; Minakawa, N.; Marín Rodríguez, R.; Chaves, L.F. New and common haplotypes shape genetic diversity in asian tiger mosquito populations from Costa Rica and Panamá. J. Econ. Entomol. 2015, 108, 761–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battaglia, V.; Gabrieli, P.; Brandini, S.; Capodiferro, M.R.; Javier, P.A.; Chen, X.-G.; Achilli, A.; Semino, O.; Gomulski, L.M.; Malacrida, A.R.; et al. The worldwide spread of the tiger mosquito as revealed by mitogenome haplogroup diversity. Front. Genet. 2016, 7, 208. [Google Scholar] [CrossRef] [Green Version]
- Walther, D.; Scheuch, D.E.; Kampen, H. The invasive Asian tiger mosquito Aedes albopictus (Diptera: Culicidae) in Germany: Local reproduction and overwintering. Acta Trop. 2017, 166, 186–192. [Google Scholar] [CrossRef]
- Lee, E.; Yang, S.-C.; Kim, T.-K.; Noh, B.-E.; Lee, H.S.; Kim, H.; Roh, J.Y.; Lee, W.-G. Geographical genetic variation and sources of Korean Aedes albopictus (Diptera: Culicidae) populations. J. Med. Entomol. 2020, 57, 1057–1068. [Google Scholar] [CrossRef]
- Zhong, D.; Lo, E.; Hu, R.; Metzger, M.E.; Cummings, R.; Bonizzoni, M.; Fujioka, K.K.; Sorvillo, T.E.; Kluh, S.; Healy, S.P.; et al. Genetic analysis of invasive Aedes albopictus populations in Los Angeles County, California and its potential public health impact. PLoS ONE 2013, 8, e68586. [Google Scholar] [CrossRef] [Green Version]
- Unal, I. Defining an optimal cut-point value in roc analysis: An alternative approach. Comput. Math. Methods Med. 2017, 2017, 3762651. [Google Scholar] [CrossRef]
- Fischer, D.; Thomas, S.M.; Neteler, M.; Tjaden, N.B.; Beierkuhnlein, C. Climatic suitability of Aedes albopictus in Europe referring to climate change projections: Comparison of mechanistic and correlative niche modelling approaches. Eurosurveillance 2014, 19, 20696. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Nihei, N.; Kurihara, T. Analysis of northern distribution of Aedes albopictus (Diptera: Culicidae) in Japan by geographical information system. J. Med. Entomol. 2002, 39, 4–11. [Google Scholar] [CrossRef] [Green Version]
- Stone, C.M.; Zuo, Z.; Li, B.; Ruiz, M.; Swanson, J.; Hunt, J.; Kim, C.-H.; Smith, R.L. Spatial, temporal, and genetic invasion dynamics of Aedes albopictus (Diptera: Culicidae) in Illinois. J. Med. Entomol. 2020, 57, 1488–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, T.L.; Rašić, G.; Zhang, D.; Zheng, X.; Xi, Z.; Hoffmann, A.A. Genome-wide SNPs reveal the drivers of gene flow in an urban population of the Asian Tiger Mosquito, Aedes albopictus. PLoS Negl. Trop. Dis. 2017, 11, e0006009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawley, W.A. The biology of Aedes albopictus. J. Am. Mosq. Control Assoc. Suppl. 1988, 1, 1–39. [Google Scholar] [PubMed]
- Wellmer, H. Dengue Haemorrhagic Fever in Thailand; Springer: Berlin/Heidelberg, Germany, 1983; ISBN 978-3-642-69154-6. [Google Scholar]
- Kraemer, M.U.G.; Reiner, R.C.; Brady, O.J.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; Yi, D.; Johnson, K.; Earl, L.; Marczak, L.B.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat. Microbiol. 2019, 4, 854–863. [Google Scholar] [CrossRef]
- Dutta, P.; Khan, S.A.; Sharma, C.K.; Doloi, P.; Hazarika, N.C.; Mahanta, J. Distribution of potential dengue vectors in major townships along the national highways and trunk roads of northeast India. Southeast Asian J. Trop. Med. Public Health 1998, 29, 173–176. [Google Scholar]
- Oliveira, S.; Rocha, J.; Sousa, C.A.; Capinha, C. Wide and increasing suitability for Aedes albopictus in Europe is congruent across distribution models. Sci. Rep. 2021, 11, 9916. [Google Scholar] [CrossRef]
- Kraemer, M.U.G.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.N.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. Elife 2015, 4, e08347. [Google Scholar] [CrossRef]
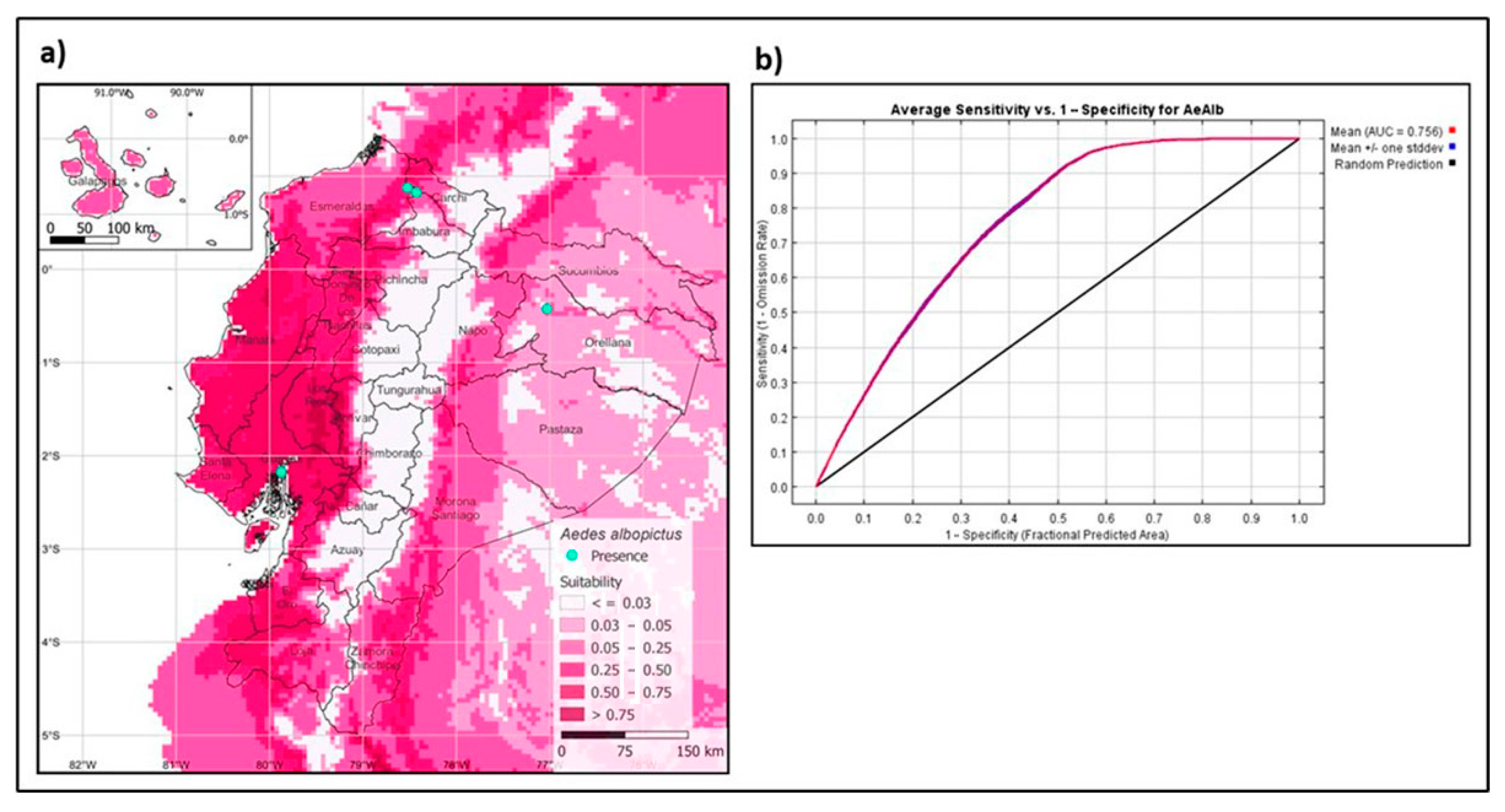

{kind=link}
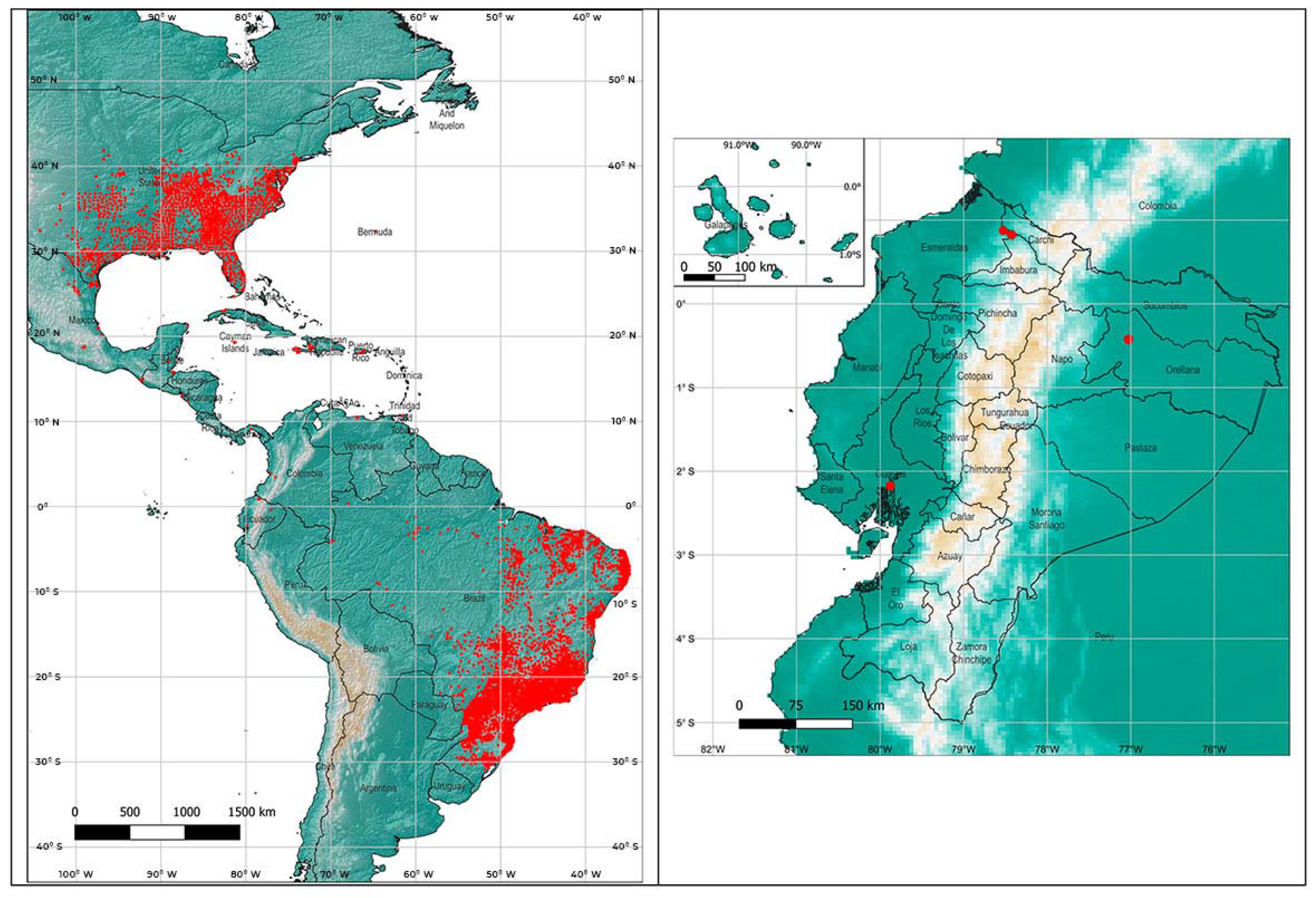
{kind=link}
{kind=link}
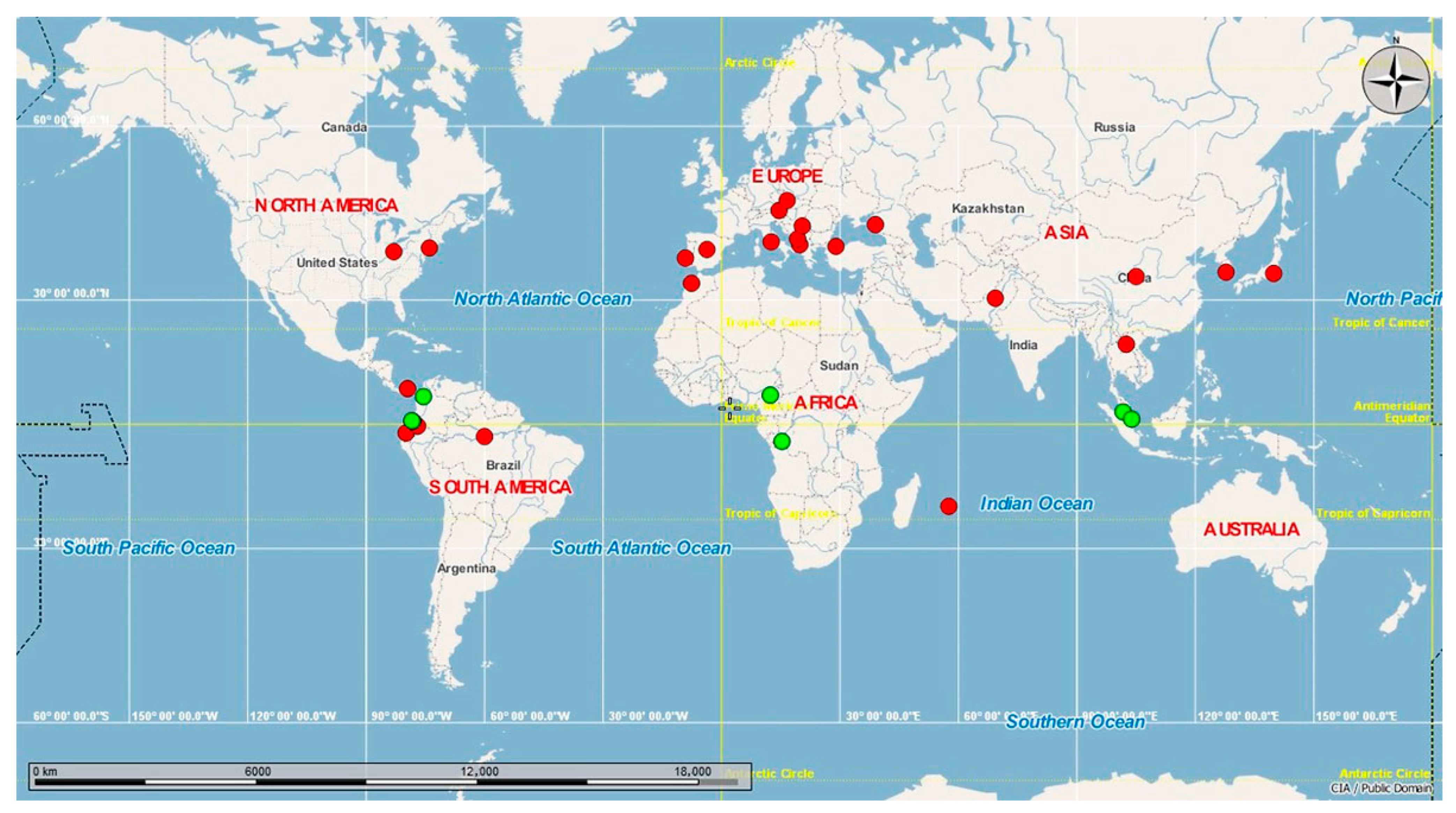
{kind=link}
{kind=link}
| Region | Province | Locality | Latitude | Longitude | Collection Year | Sample Size |
|---|---|---|---|---|---|---|
| Andean Lowland | Imbabura | Lita | 0.8337 | −78.4017 | 2018–2020 | 12 |
| Imbabura | Guadual | 0.8918 | −78.5022 | 2021 | 10 | |
| Imbabura | Cachaco | 0.8337 | −78.4023 | 2018–2019, 2021 | 12 | |
| Pacific coast | Guayas | Guayaquil | −2.1633 | −79.8938 | 2017 | 10 |
| Amazon basin | Orellana | Francisco de Orellana | −0.4412 | −77.0048 | 2018–2020 | 7 |
| Guayaquil | Puerto Francisco de Orellana | Lita | Cachaco | Guadual | |
|---|---|---|---|---|---|
| Guayaquil | 0 | ||||
| Puerto Francisco de Orellana | 0.20 | 0 | |||
| Lita | 0.29 a | 0.54 a | 0 | ||
| Cachaco | 0.33 a | 0.60 a | −0.01 | 0 | |
| Guadual | 0.30 a | 0.56 a | −0.02 | −0.11 | 0 |
| Number of Groups | Partitions | Test | Among Groups | Among Populations | Within Groups |
|---|---|---|---|---|---|
| 2 | (1,2,3,4) (5) | Coast vs. Amazon basin | 38.35 | 11.26 | 50.39 |
| 2 | (1,2,3) (4,5) | Haplotype 1 vs. haplotype 2 | 39.87 | 3.74 | 56.38 |
| 3 | (1,2,3) (4) (5) | NW coast, SW Coast and Amazonia | 45.46 | −2.30 | 56.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrazco-Montalvo, A.; Ponce, P.; Villota, S.D.; Quentin, E.; Muñoz-Tobar, S.; Coloma, J.; Cevallos, V. Establishment, Genetic Diversity, and Habitat Suitability of Aedes albopictus Populations from Ecuador. Insects 2022, 13, 305. https://doi.org/10.3390/insects13030305
Carrazco-Montalvo A, Ponce P, Villota SD, Quentin E, Muñoz-Tobar S, Coloma J, Cevallos V. Establishment, Genetic Diversity, and Habitat Suitability of Aedes albopictus Populations from Ecuador. Insects. 2022; 13(3):305. https://doi.org/10.3390/insects13030305
Chicago/Turabian StyleCarrazco-Montalvo, Andrés, Patricio Ponce, Stephany D. Villota, Emmanuelle Quentin, Sofía Muñoz-Tobar, Josefina Coloma, and Varsovia Cevallos. 2022. "Establishment, Genetic Diversity, and Habitat Suitability of Aedes albopictus Populations from Ecuador" Insects 13, no. 3: 305. https://doi.org/10.3390/insects13030305
APA StyleCarrazco-Montalvo, A., Ponce, P., Villota, S. D., Quentin, E., Muñoz-Tobar, S., Coloma, J., & Cevallos, V. (2022). Establishment, Genetic Diversity, and Habitat Suitability of Aedes albopictus Populations from Ecuador. Insects, 13(3), 305. https://doi.org/10.3390/insects13030305