
First Nationwide Monitoring Program for the Detection of Potentially Invasive Mosquito Species in Austria

 , , ,
, , ,  , ,
, ,  add
Show full author list
add
Show full author list
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Trap Sites and Catch Scheme
2.2. Sample Analysis
2.3. Data Analysis
3. Results
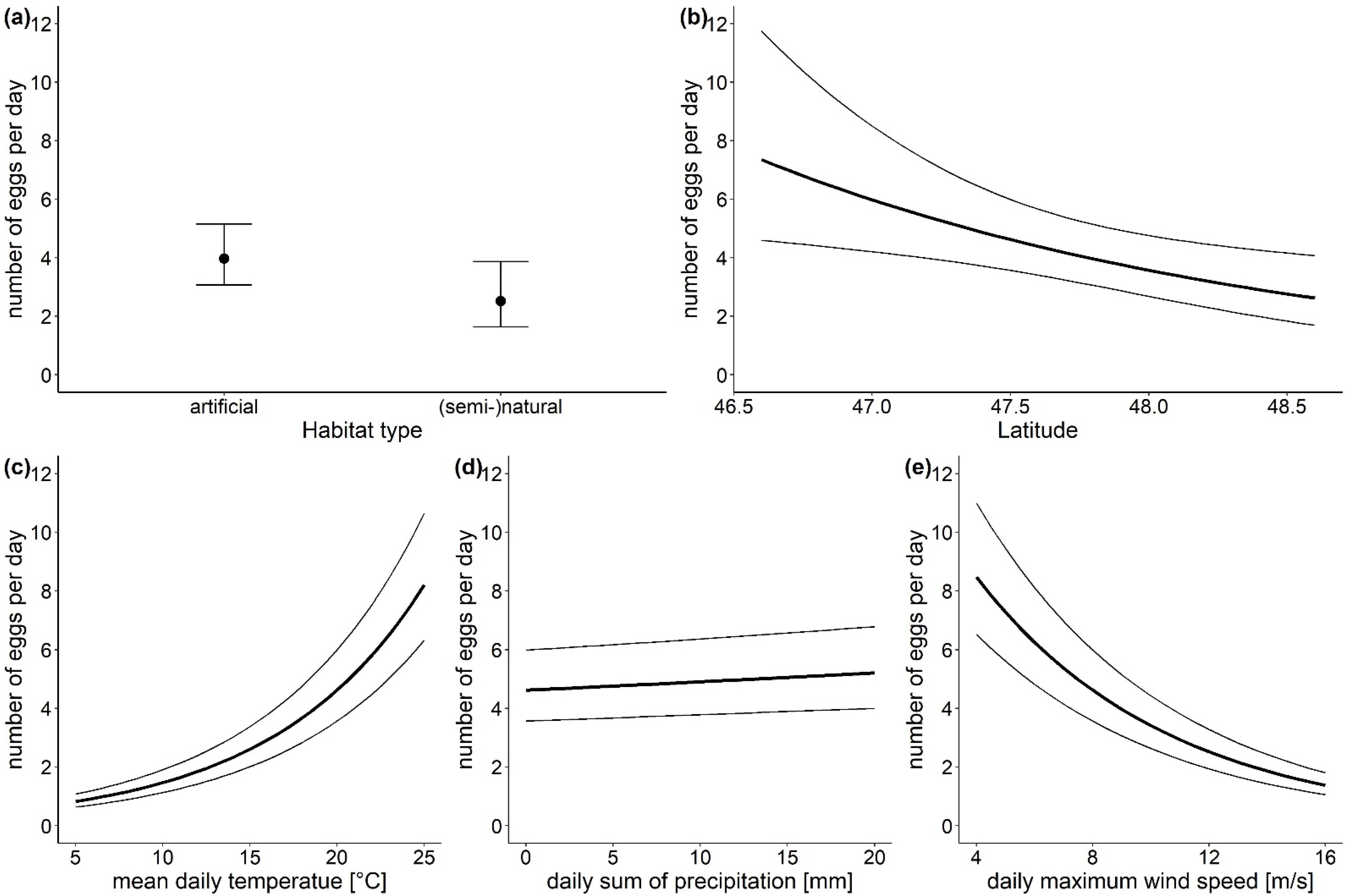
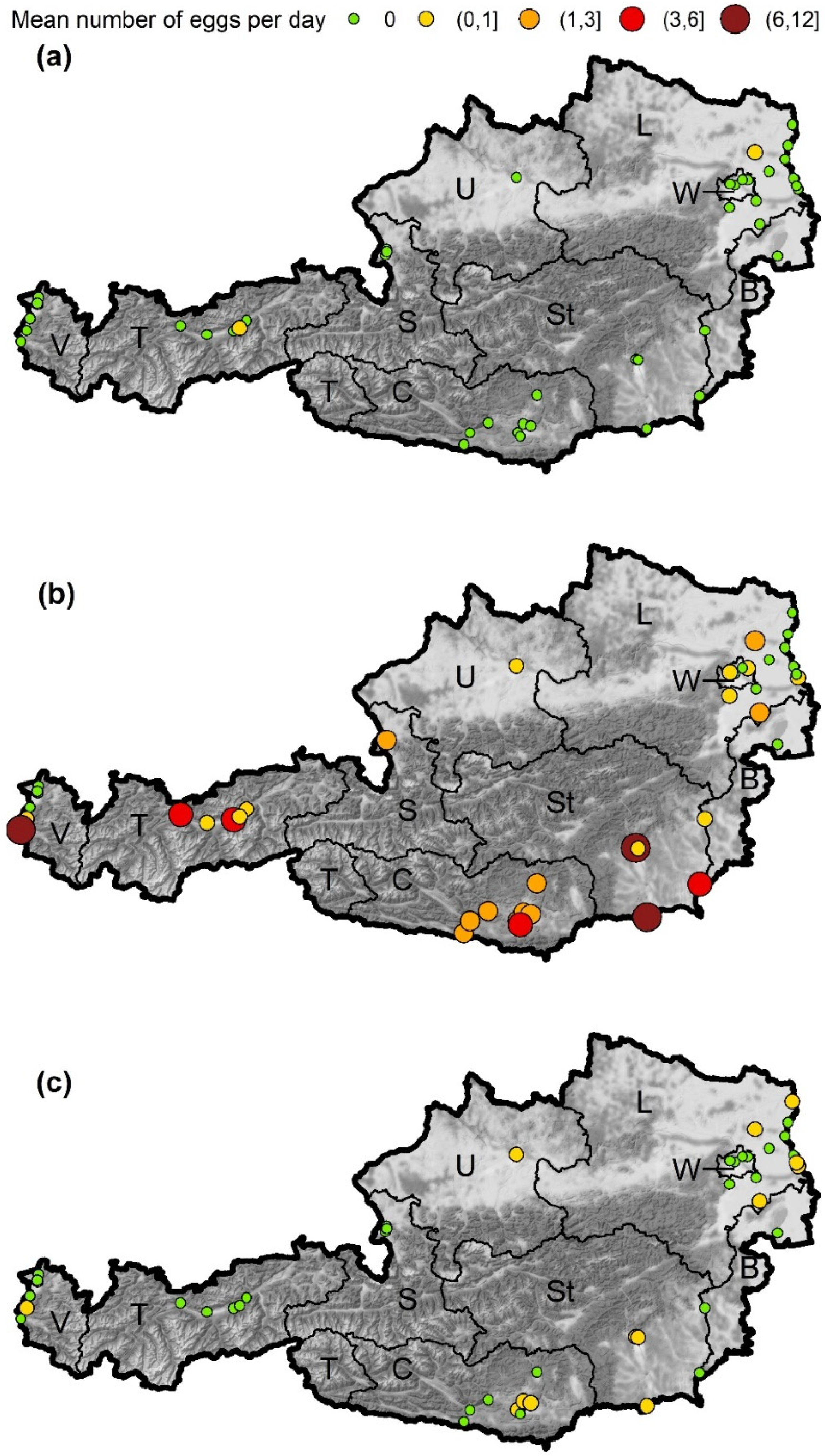
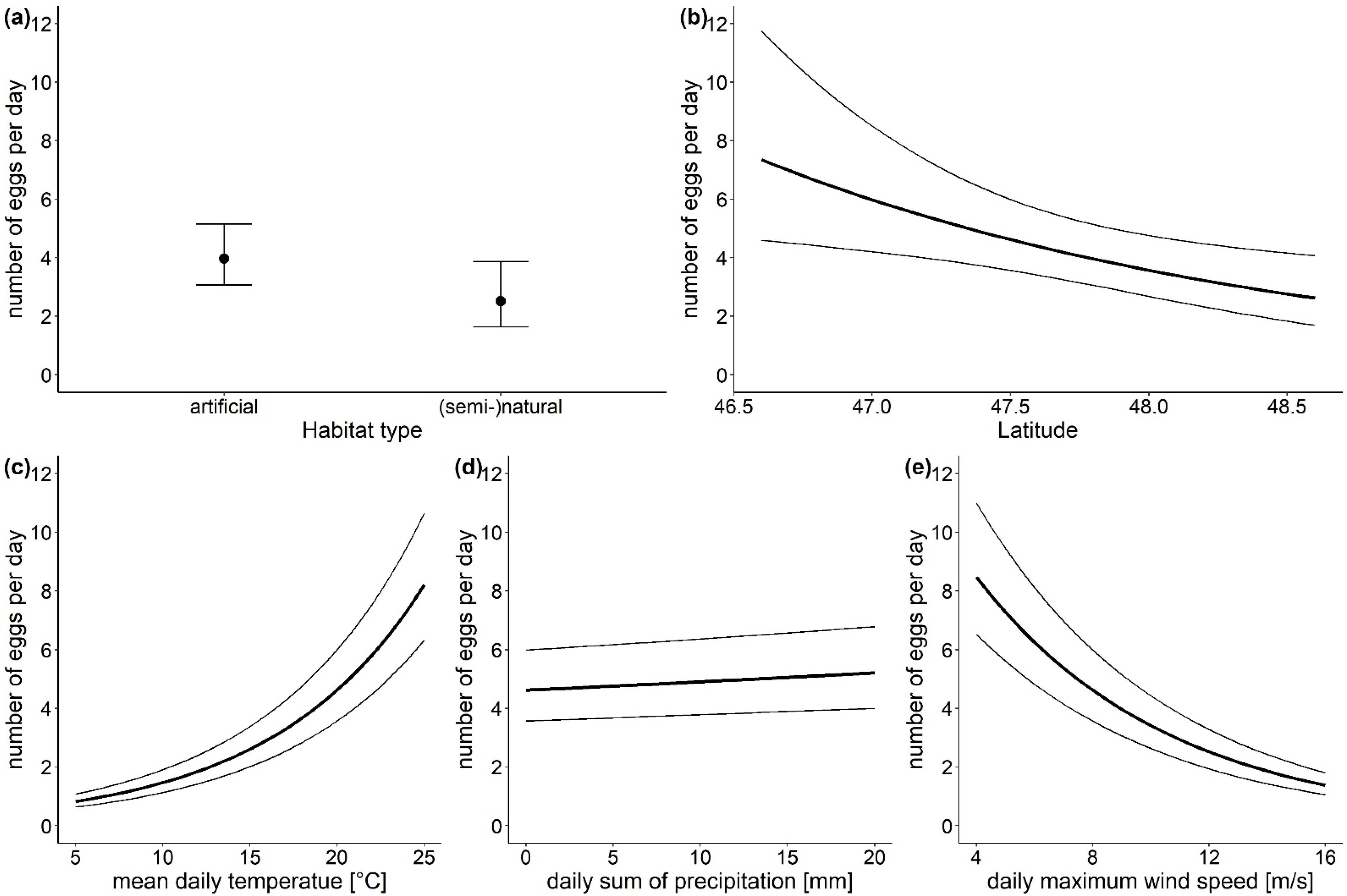
3.1. The Overall Number of Aedes-Eggs
3.2. Aedes albopictus
3.3. Aedes japonicus
3.4. Aedes geniculatus
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Medlock, J.; Hansford, K.M.; Versteirt, V.; Cull, B.; Kampen, H.; Fontenille, D.; Hendrickx, G.; Zeller, H.; Van Bortel, W.; Schaffner, F. An entomological review of invasive mosquitoes in Europe. Bull. Entomol. Res. 2015, 105, 637–663. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.; Hansford, K.M.; Schaffner, F.; Versteirt, V.; Hendrickx, G.; Zeller, H.; Van Bortel, W. A review of the invasive mosquitoes in Europe: Ecology, public health risks, and control options. Vector-Borne Zoonotic Dis. 2012, 12, 435–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholte, E.-J.; Schaffner, F. Waiting for the tiger—Establishment and spread of Aedes albopictus mosquito in Europe. In Emerging Pests and Vector-Borne Diseases in Europe. Volume 1: Ecology and Contro of Vector-Borne Diseases; Takken, W., Knols, B.G.J., Eds.; Wageningen Academic: Wageningen, The Netherlands, 2007; pp. 241–260. [Google Scholar]
- Eritja, R.; Palmer, J.R.B.; Roiz, D.; Sanpera-Calbet, I.; Bartumeus, F. Direct evidence of adult Aedes albopictus dispersal by car. Sci. Rep. 2017, 7, 14399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunze, S.; Kochmann, J.; Koch, L.K.; Klimpel, S. Aedes albopictus and its environmental limits in Europe. PLoS ONE 2016, 11, e0162116. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Kamara, F.; Zhou, G.; Puthiyakunnon, S.; Li, C.; Liu, Y.; Zhou, Y.; Yao, L.; Yan, G.; Chen, X.G. Urbanization increases Aedes albopictus larval habitats and accelerates mosquito development and survivorship. PLoS Negl. Trop. Dis. 2014, 8, e3301. [Google Scholar] [CrossRef] [Green Version]
- Caminade, C.; Medlock, J.M.; Ducheyne, E.; McIntyre, K.M.; Leach, S.; Baylis, M.; Morse, A.P. Suitability of European climate for the Asian tiger mosquito Aedes albopictus: Recent trends and future scenarios. J. R. Soc. Interface 2012, 9, 2708–2717. [Google Scholar] [CrossRef] [Green Version]
- Kraemer, M.U.G.; Reiner, R.C.; Brady, O.J.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; Yi, D.; Johnson, K.; Earl, L.; Marczak, L.B.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat. Microbiol. 2019, 4, 854–863. [Google Scholar] [CrossRef]
- Beck, K.G.; Zimmerman, K.; Schardt, J.D.; Stone, J.; Lukens, R.R.; Reichard, S.; Randall, J.; Cangelosi, A.A.; Cooper, D.; Thompson, J.P. Invasive species defined in a policy context: Recommendations from the Federal Invasive Species Advisory Committee. Invasive Plant Sci. Manag. 2008, 1, 414–421. [Google Scholar] [CrossRef]
- ECDC Guidelines for the Surveillance of Invasive Mosquitoes in Europe; European Centre for Disease Prevention and Control: Stockholm, Sweden, 2012; ISBN 9789291933785.
- Fuehrer, H.P.; Morelli, S.; Unterköfler, M.S.; Bajer, A.; Bakran-Lebl, K.; Dwużnik-Szarek, D.; Farkas, R.; Grandi, G.; Heddergott, M.; Jokelainen, P.; et al. Dirofilaria spp. and Angiostrongylus vasorum: Current risk of spreading in central and northern Europe. Pathogens 2021, 10, 1268. [Google Scholar] [CrossRef]
- Schoener, E.; Zittra, C.; Weiss, S.; Walder, G.; Barogh, B.S.; Weiler, S.; Fuehrer, H.P. Monitoring of alien mosquitoes of the genus Aedes (Diptera: Culicidae) in Austria. Parasitol. Res. 2019, 118, 1633–1638. [Google Scholar] [CrossRef] [Green Version]
- Seidel, B.; Duh, D.; Nowotny, N.; Allerberger, F. Erstnachweis der Stechmücken Aedes (Ochlerotatus) japonicus japonicus (Theobald, 1901) in Österreich und Slowenien in 2011 und für Aedes (Stegomyia) albopictus (Skuse, 1895) in Österreich 2012 (Diptera: Culicidae). Entomol. Z. 2012, 112, 223–226. [Google Scholar]
- Seidel, B.; Nowotny, N.; Bakonyi, T.; Allerberger, F.; Schaffner, F. Spread of Aedes japonicus japonicus (Theobald, 1901) in Austria, 2011-2015, and first records of the subspecies for Hungary, 2012, and the principality of Liechtenstein, 2015. Parasites Vectors 2016, 9, 356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuehrer, H.P.; Schoener, E.; Weiler, S.; Barogh, B.S.; Zittra, C.; Walder, G. Monitoring of alien mosquitoes in Western Austria (Tyrol, Austria, 2018). PLoS Negl. Trop. Dis. 2020, 14, e0008433. [Google Scholar] [CrossRef] [PubMed]
- Bakran-Lebl, K.; Zittra, C.; Harl, J.; Shahi-Barogh, B.; Grätzl, A.; Ebmer, D.; Schaffner, F.; Fuehrer, H.P. Arrival of the Asian tiger mosquito, Aedes albopictus (Skuse, 1895) in Vienna, Austria and initial monitoring activities. Transbound. Emerg. Dis. 2021, 68, 3145–3150. [Google Scholar] [CrossRef]
- Seidel, B.; Hufnagl, P.; Nowotny, N.; Allerberger, F.; Indra, A. Long-term field study on mosquitoes in Austria, in particular the invasive Korean bush-mosquito Aedes (Finlaya) koreicus (Edwards, 1917). Beitr. Entomofaunist. 2020, 21, 237–240. [Google Scholar]
- Gloria-Soria, A.; Payne, A.F.; Bialosuknia, S.M.; Stout, J.; Mathias, N.; Eastwood, G.; Ciota, A.T.; Kramer, L.D.; Armstrong, P.M. Vector competence of Aedes albopictus populations from the Northeastern United States for Chikungunya, Dengue, and Zika Viruses. Am. J. Trop. Med. Hyg. 2021, 104, 1123–1130. [Google Scholar] [CrossRef]
- McKenzie, B.A.; Wilson, A.E.; Zohdy, S. Aedes albopictus is a competent vector of Zika virus: A meta-analysis. PLoS ONE 2019, 14, e0216794. [Google Scholar] [CrossRef] [Green Version]
- Cancrini, G.; Romi, R.; Gabrielli, S.; Toma, L.; Di Paolo, M.; Scaramozzino, P. First finding of Dirofilaria repens in a natural population of Aedes albopictus. Med. Vet. Entomol. 2003, 17, 448–451. [Google Scholar] [CrossRef]
- Rezza, G.; Nicoletti, L.; Angelini, R.; Romi, R.; Finarelli, A.C.; Panning, M.; Cordioli, P.; Fortuna, C.; Boros, S.; Magurano, F.; et al. Infection with chikungunya virus in Italy: An outbreak in a temperate region. Lancet 2007, 370, 1840–1846. [Google Scholar] [CrossRef]
- Grandadam, M.; Caro, V.; Plumet, S.; Thiberge, J.M.; Souarès, Y.; Failloux, A.B.; Tolou, H.J.; Budelot, M.; Cosserat, D.; Leparc-Goffart, I.; et al. Chikungunya virus, Southeastern France. Emerg. Infect. Dis. 2011, 17, 910–913. [Google Scholar] [CrossRef]
- Marchand, E.; Prat, C.; Jeannin, C.; Lafont, E.; Bergmann, T.; Flusin, O.; Rizzi, J.; Roux, N.; Busso, V.; Deniau, J.; et al. Autochthonous case of dengue in France, October 2013. Eurosurveillance 2013, 18, 20661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veronesi, E.; Paslaru, A.; Silaghi, C.; Tobler, K.; Glavinic, U.; Torgerson, P.; Mathis, A. Experimental evaluation of infection, dissemination, and transmission rates for two West Nile virus strains in European Aedes japonicus under a fluctuating temperature regime. Parasitol. Res. 2018, 117, 1925–1932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turell, M.J.; Dohm, D.J.; Sardelis, M.R.; O’Guinn, M.L.; Andreadis, T.G.; Blow, J.A. An update on the potential of North American mosquitoes (Diptera: Culicidae) to transmit West Nile virus. J. Med. Entomol. 2005, 42, 57–62. [Google Scholar] [CrossRef]
- Lebl, K.; Zittra, C.; Silbermayr, K.; Obwaller, A.; Berer, D.; Brugger, K.; Walter, M.; Pinior, B.; Fuehrer, H.-P.; Rubel, F. Mosquitoes (Diptera: Culicidae) and their relevance as disease vectors in the city of Vienna, Austria. Parasitol. Res. 2015, 114, 707–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakran-Lebl, K.; Zittra, C.; Weiss, S.; Hodits, B.; Zechmeister, T.; Fuehrer, H.P. Range expansion of the alien mosquito species Aedes japonicus (Theobald, 1901) (Diptera: Culicidae) from 2014–2019 in Burgenland, Austria. Entomol. Austriaca 2021, 28, 107–118. [Google Scholar]
- Schaffner, F.; Kaufmann, C.; Pflüger, V.; Mathis, A. Rapid protein profiling facilitates surveillance of invasive mosquito species. Parasites Vectors 2014, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Bova, J.E.; Paulson, S.L.; Paulson, G. Morphological differentiation of the eggs of North American container-inhabiting Aedes mosquitoes. J. Am. Mosq. Control Assoc. 2016, 32, 244–246. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Lüdecke, D. ggeffects: Tidy data frames of marginal effects from regression models. J. Open Source Softw. 2018, 3, 772. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Kahle, D.; Wickham, H. ggmap: Spatial Visualization with ggplot2. R J. 2013, 5, 144–161. [Google Scholar] [CrossRef] [Green Version]
- Bivand, R.; Keitt, T.; Rowlingson, B. Rgdal: Bindings for the “Geospatial” Data Abstraction Library. Available online: https://cran.r-project.org/web/packages/rgdal/rgdal.pdf (accessed on 18 January 2022).
- Wilke, C.O. Cowplot: Streamlined Plot Theme and Plot Annotations for “ggplot2”. Available online: https://cran.r-project.org/web/packages/cowplot/cowplot.pdf (accessed on 18 January 2022).
- Gradoni, F.; Bertola, M.; Carlin, S.; Accordi, S.; Toniolo, F.; Visentin, P.; Patregnani, T.; Adami, S.; Terzo, L.; Dal Pont, M.; et al. Geographical data on the occurrence and spreading of invasive Aedes mosquito species in Northeast Italy. Data Brief 2021, 36, 107047. [Google Scholar] [CrossRef] [PubMed]
- Flacio, E.; Engeler, L.; Tonolla, M.; Müller, P. Spread and establishment of Aedes albopictus in southern Switzerland between 2003 and 2014: An analysis of oviposition data and weather conditions. Parasites Vectors 2016, 9, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, N.; Geier, M.; Balczun, C.; Bradersen, U.; Huber, K.; Kiel, E.; Krüger, A.; Lühken, R.; Orendt, C.; Plenge-Bönig, A.; et al. Repeated introduction of Aedes albopictus into Germany, July to October 2012. Parasitol. Res. 2013, 112, 1787–1790. [Google Scholar] [CrossRef] [PubMed]
- Müller, P.; Engeler, L.; Vavassori, L.; Suter, T.; Guidi, V.; Gschwind, M.; Tonolla, M.; Flacio, E. Surveillance of invasive Aedes mosquitoes along Swiss traffic axes reveals different dispersal modes for Aedes albopictus and Ae. japonicus. PLoS Negl. Trop. Dis. 2020, 14, 1–20. [Google Scholar] [CrossRef]
- Bundesminister für Soziales, Gesundheit, Pflege und Konsumentenschutz. 87. Verordnung: Maßnahmen bei der Einreise aus Italien; Bundesgesetzblatt für die Republick Österreich: Vienna, Austria, 2020; p. 1. [Google Scholar]
- Šebesta, O.; Rudolf, I.; Betášová, L.; Peško, J.; Hubálek, Z. An invasive mosquito species Aedes albopictus found in the Czech Republic, 2012. Euro Surveill. 2012, 17, 20301. [Google Scholar] [CrossRef]
- Rudolf, I.; Blažejová, H.; Straková, P.; Šebesta, O.; Peško, J.; Mendel, J.; Šikutová, S.; Hubálek, Z.; Kampen, H.; Schaffner, F. The invasive Asian tiger mosquito Aedes albopictus (Diptera: Culicidae) in the Czech Republic: Repetitive introduction events highlight the need for extended entomological surveillance. Acta Trop. 2018, 185, 239–241. [Google Scholar] [CrossRef]
- Chaves, L.F.; Friberg, M.D.; Moji, K. Synchrony of globally invasive Aedes spp. immature mosquitoes along an urban altitudinal gradient in their native range. Sci. Total Environ. 2020, 734, 139365. [Google Scholar] [CrossRef]
- Bartlett-Healy, K.; Unlu, I.; Obenauer, P.; Hughes, T.; Healy, S.; Crepeau, T.; Farajollahi, A.; Kesavaraju, B.; Fonseca, D.; Schoeler, G.; et al. Larval mosquito habitat utilization and community dynamics of Aedes albopictus and Aedes japonicus (Diptera: Culicidae). J. Med. Entomol. 2012, 49, 813–824. [Google Scholar] [CrossRef]
- Tanaka, K.; Mizusawa, K.; Saugstad, E.S. A revision of the adult and larval mosquitoes of Japan (including the Ryukyu Archipelago and the Ogasawara Islands) and Korea (Diptera: Culicidae). Contrib. Am. Entomol. Inst. 1979, 16, 1–987. [Google Scholar]
- Früh, L.; Kampen, H.; Koban, M.B.; Pernat, N.; Schaub, G.A.; Werner, D. Oviposition of Aedes japonicus japonicus (Diptera: Culicidae) and associated native species in relation to season, temperature and land use in western Germany. Parasites Vectors 2020, 13, 623. [Google Scholar] [CrossRef] [PubMed]
- Murrell, E.G.; Noden, B.H.; Juliano, S.A. Contributions of temporal segregation, oviposition choice, and non-additive effects of competitors to invasion success of Aedes japonicus (Diptera: Culicidae) in North America. Biol. Invasions 2015, 17, 1669–1681. [Google Scholar] [CrossRef] [PubMed]
- Krupa, E.; Henon, N.; Mathieu, B. Diapause characterisation and seasonality of Aedes japonicus japonicus (Diptera, Culicidae) in the northeast of France. Parasite 2021, 28, 45. [Google Scholar] [CrossRef] [PubMed]
- Reuss, F.; Wieser, A.; Niamir, A.; Bálint, M.; Kuch, U.; Pfenninger, M.; Müller, R. Thermal experiments with the Asian bush mosquito (Aedes japonicus japonicus) (Diptera: Culicidae) and implications for its distribution in Germany. Parasites Vectors 2018, 11, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrd, B.D.; Sither, C.B.; Goggins, J.A.; Kunze-Garcia, S.; Pesko, K.N.; Bustamante, D.M.; Sither, J.M.; Vonesh, J.R.; O’Meara, G.F. Aquatic thermal conditions predict the presence of native and invasive rock pool Aedes (Diptera: Culicidae) in the southern Appalachians, U.S.A. J. Vector Ecol. 2019, 44, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Kirik, H.; Burtin, V.; Tummeleht, L.; Kurina, O. Friends in all the green spaces: Weather dependent changes in urban mosquito (Diptera: Culicidae) abundance and diversity. Insects 2021, 12, 352. [Google Scholar] [CrossRef]
- Lebl, K.; Brugger, K.; Rubel, F. Predicting Culex pipiens/restuans population dynamics by interval lagged weather data. Parasites Vectors 2013, 6, 129. [Google Scholar] [CrossRef] [Green Version]
- Wong, G.K.L.; Jim, C.Y. Urban-microclimate effect on vector mosquito abundance of tropical green roofs. Build. Environ. 2017, 112, 63–76. [Google Scholar] [CrossRef]
- Hoffmann, E.J.; Miller, J.R. Reassessment of the role and utility of wind in Suppression of mosquito (Diptera: Culicidae) host finding: Stimulus dilution supported over flight limitation. J. Med. Entomol. 2003, 40, 607–614. [Google Scholar] [CrossRef]
- Rochlin, I.; Gaugler, R.; Williges, E.; Farajollahi, A. The rise of the invasives and decline of the natives: Insights revealed from adult populations of container-inhabiting Aedes mosquitoes (Diptera: Culicidae) in temperate North America. Biol. Invasions 2013, 15, 991–1003. [Google Scholar] [CrossRef] [Green Version]
- Chaves, L.F.; Kitron, U.D. Weather variability impacts on oviposition dynamics of the southern house mosquito at intermediate time scales. Bull. Entomol. Res. 2011, 101, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Strickman, D. Rate of oviposition by Culex quinquefasciatus in San Antonio, Texas, during three years. J. Am. Mosq. Control Assoc. 1988, 4, 339–344. [Google Scholar] [PubMed]
- Müller, G.C.; Kravchenko, V.D.; Junnila, A.; Schlein, Y. Tree-hole breeding mosquitoes in Israel. J. Vector Ecol. 2012, 37, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Zittra, C.; Vitecek, S.; Obwaller, A.G.; Rossiter, H.; Eigner, B.; Zechmeister, T.; Waringer, J.; Fuehrer, H.P. Landscape structure affects distribution of potential disease vectors (Diptera: Culicidae). Parasites Vectors 2017, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zittra, C.; Waringer, J. Species inventory, ecology, and seasonal distribution patterns of Culicidae (Insecta: Diptera) in the National Park Donau-Auen (Lower Austria). Aquat. Insects 2014, 36, 63–77. [Google Scholar] [CrossRef]
- Prudhomme, J.; Fontaine, A.; Lacour, G.; Gantier, J.C.; Diancourt, L.; Velo, E.; Bino, S.; Reiter, P.; Mercier, A. The native European Aedes geniculatus mosquito species can transmit chikungunya virus. Emerg. Microbes Infect. 2019, 8, 962–972. [Google Scholar] [CrossRef] [Green Version]
- Silaghi, C.; Beck, R.; Capelli, G.; Montarsi, F.; Mathis, A. Development of Dirofilaria immitis and Dirofilaria repens in Aedes japonicus and Aedes geniculatus. Parasites Vectors 2017, 10, 94. [Google Scholar] [CrossRef] [Green Version]
- Pernat, N.; Kampen, H.; Jeschke, J.M.; Werner, D. Citizen science versus professional data collection: Comparison of approaches to mosquito monitoring in Germany. J. Appl. Ecol. 2021, 58, 214–223. [Google Scholar] [CrossRef]
- Eritja, R.; Delacour-Estrella, S.; Ruiz-Arrondo, I.; González, M.A.; Barceló, C.; García-Pérez, A.L.; Lucientes, J.; Miranda, M.; Bartumeus, F. At the tip of an iceberg: Citizen science and active surveillance collaborating to broaden the known distribution of Aedes japonicus in Spain. Parasites Vectors 2021, 14, 375. [Google Scholar] [CrossRef]
- Eritja, R.; Ruiz-Arrondo, I.; Delacour-Estrella, S.; Schaffner, F.; Álvarez-Chachero, J.; Bengoa, M.; Puig, M.Á.; Melero-Alcíbar, R.; Oltra, A.; Bartumeus, F. First detection of Aedes japonicus in Spain: An unexpected finding triggered by citizen science. Parasites Vectors 2019, 12, 53. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Conditional Model | Estimate | Standard Error | z Value | p |
|---|---|---|---|---|
| intercept | 24.465 | 9.1326 | 2.679 | 0.0074 |
| habitat type—artificial surfaces | 0.457 | 0.2245 | 2.037 | 0.0417 |
| latitude | −0.515 | 0.1926 | −2.675 | 0.0075 |
| altitude—450–800 m a.s.l | 0.073 | 0.2320 | 0.316 | 0.7519 |
| temperature | 0.115 | 0.0018 | 64.725 | <0.0001 |
| precipitation | 0.006 | 0.0016 | 3.761 | 0.0002 |
| wind speed | −0.151 | 0.0043 | −35.093 | <0.0001 |
| random effects | ||||
| Variance | Std. Dev. | |||
| position:site | 0.4589 | 0.6774 | ||
| site | 0.1319 | 0.3632 | ||
| Zero-Inflated Model | Estimate | Standard Error | z Value | p |
| intercept | −126.355 | 37.4811 | −3.371 | 0.0007 |
| habitat type—artificial surfaces | −0.708 | 0.5001 | −1.415 | 0.1572 |
| latitude | 2.853 | 0.7872 | 3.625 | 0.0003 |
| altitude—450–800 m a.s.l | −0.937 | 1.0420 | −0.900 | 0.3683 |
| temperature | −0.329 | 0.0184 | −17.835 | <0.0001 |
| precipitation | −0.055 | 0.0181 | −3.057 | 0.0022 |
| wind speed | 0.119 | 0.0417 | 2.850 | 0.0044 |
| random effects | ||||
| Variance | Std. Dev. | |||
| position:site | 0.991 | 0.9955 | ||
| site | 5.101 | 2.2585 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakran-Lebl, K.; Pree, S.; Brenner, T.; Daroglou, E.; Eigner, B.; Griesbacher, A.; Gunczy, J.; Hufnagl, P.; Jäger, S.; Jerrentrup, H.; et al. First Nationwide Monitoring Program for the Detection of Potentially Invasive Mosquito Species in Austria. Insects 2022, 13, 276. https://doi.org/10.3390/insects13030276
Bakran-Lebl K, Pree S, Brenner T, Daroglou E, Eigner B, Griesbacher A, Gunczy J, Hufnagl P, Jäger S, Jerrentrup H, et al. First Nationwide Monitoring Program for the Detection of Potentially Invasive Mosquito Species in Austria. Insects. 2022; 13(3):276. https://doi.org/10.3390/insects13030276
Chicago/Turabian StyleBakran-Lebl, Karin, Stefanie Pree, Thomas Brenner, Eleni Daroglou, Barbara Eigner, Antonia Griesbacher, Johanna Gunczy, Peter Hufnagl, Stefanie Jäger, Hans Jerrentrup, and et al. 2022. "First Nationwide Monitoring Program for the Detection of Potentially Invasive Mosquito Species in Austria" Insects 13, no. 3: 276. https://doi.org/10.3390/insects13030276
APA StyleBakran-Lebl, K., Pree, S., Brenner, T., Daroglou, E., Eigner, B., Griesbacher, A., Gunczy, J., Hufnagl, P., Jäger, S., Jerrentrup, H., Klocker, L., Paill, W., Petermann, J. S., Barogh, B. S., Schwerte, T., Suchentrunk, C., Wieser, C., Wortha, L. N., Zechmeister, T., ... Fuehrer, H.-P. (2022). First Nationwide Monitoring Program for the Detection of Potentially Invasive Mosquito Species in Austria. Insects, 13(3), 276. https://doi.org/10.3390/insects13030276