

Temperature-Dependent Development Models Describing the Effects of Temperature on the Development of the Fall Armyworm Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae)

 ,
,  ,
,  ,
,  ,
, 

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Colony Maintenance
2.2. Effect of Temperature on the Fitness of S. frugiperda
2.3. Statistical Analysis
2.3.1. Effects of Temperature on the Fitness of S. frugiperda
2.3.2. Selection and Evaluation of Mathematical Models
3. Results
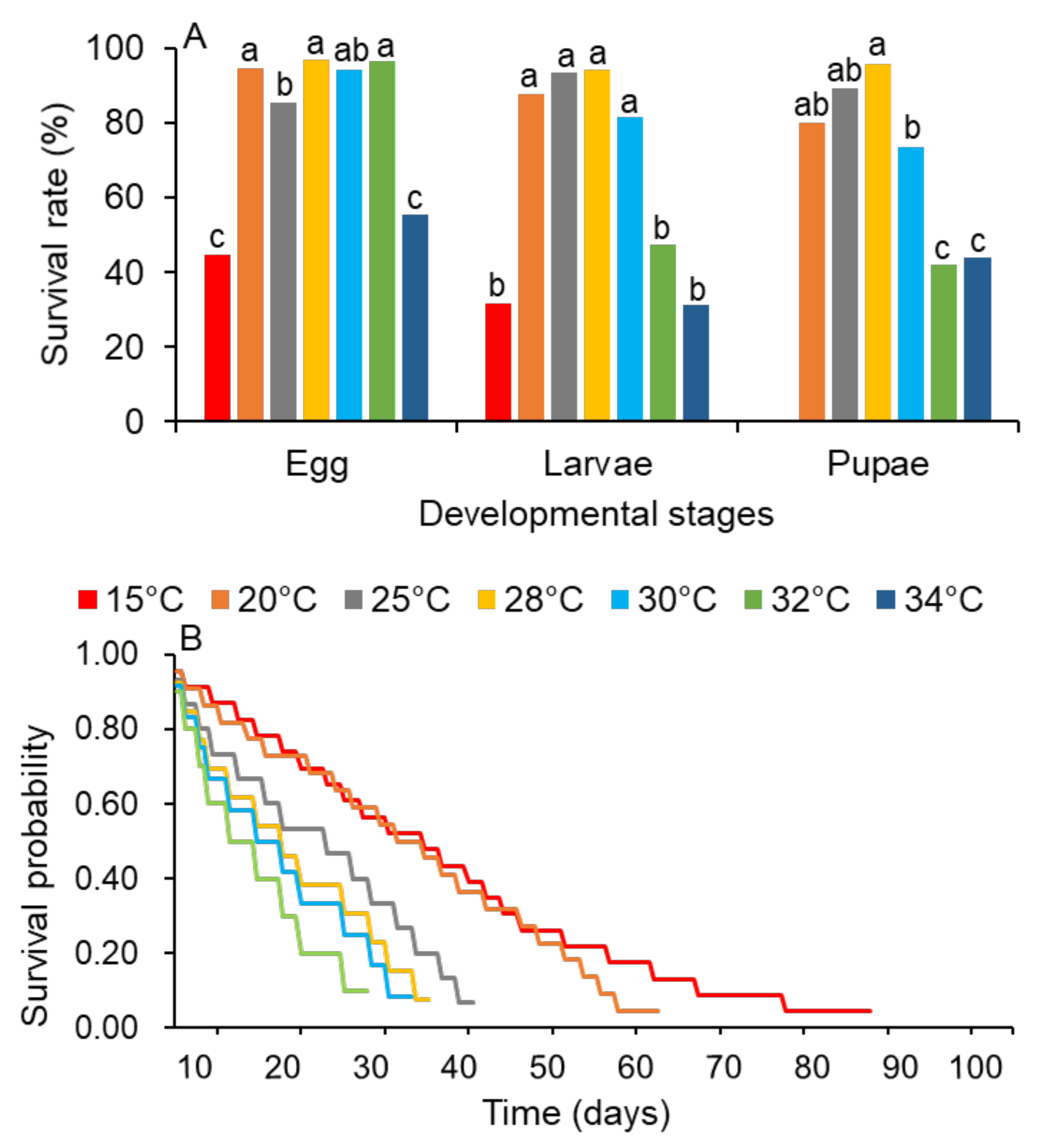
3.1. Effects of Temperature on the Fitness of S. frugiperda
3.2. Selection and Performance of Mathematical Models
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Nagoshi, R.N.; Koffi, D.; Agboka, K.; Tounou, K.A.; Banerjee, R.; Jurat-Fuentes, J.L.; Meagher, R.L. Comparative molecular analyses of invasive fall armyworm in Togo reveal strong similarities to populations from the eastern United States and the Greater Antilles. PLoS ONE 2017, 12, e0181982. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host Plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef]
- Acharya, R.; Akintola, A.A.; Malekera, M.J.; Kamulegeya, P.; Nyakunga, K.B.; Mutimbu, M.K.; Shrestha, Y.K.; Hemayet, J.S.M.; Hoat, T.X.; Dao, H.T.; et al. Genetic relationship of fall armyworm (Spodoptera frugiperda) populations that invaded Africa and Asia. Insects 2021, 12, 1–15. [Google Scholar] [CrossRef]
- Tambo, J.A.; Kansiime, M.K.; Rwomushana, I.; Mugambi, I.; Nunda, W.; Mloza Banda, C.; Nyamutukwa, S.; Makale, F.; Day, R. Impact of fall armyworm invasion on household income and food security in Zimbabwe. Food Energy Secur. 2021, 10, 299–312. [Google Scholar] [CrossRef]
- Early, R.; González-Moreno, P.; Murphy, S.T.; Day, R. Forecasting the global extent of invasion of the cereal pest Spodoptera frugiperda, the fall armyworm. NeoBiota 2018, 25–50. [Google Scholar] [CrossRef]
- Ge, S.S.; He, L.M.; He, W.; Yan, R.; Wyckhuys, K.A.G.; Wu, K.M. Laboratory-based flight performance of the fall armyworm, Spodoptera frugiperda. J. Integr. Agric. 2021, 20, 707–714. [Google Scholar] [CrossRef]
- Denlinger, D.L.; Yocum, G.D. Physiology of Heat Sensitivity. In Temperature Sensitivity in Insects and Application in Integrated Pest Management; West_View Press: Boulder, CO, USA; Oxford, UK, 1998; pp. 7–53. ISBN 0 8133 8990 9(HB). [Google Scholar]
- Ntiri, E.S.; Calatayud, P.A.; Berg, J.V.-D.; Schulthess, F.; Ru, B.P.L. Influence of temperature on intra- and interspecific resource utilization within a community of lepidopteran maize stemborers. PLoS ONE 2016, 11, e0148735. [Google Scholar] [CrossRef]
- Sampaio, F.; Krechemer, F.S.; Marchioro, C.A. Temperature-dependent development models describing the effects of temperature on the development of Spodoptera eridania. Pest Manag. Sci. 2021, 77, 919–929. [Google Scholar] [CrossRef]
- Logan, J.A.; Wollkind, D.J.; Hoyt, S.C.; Tanigoshi, L.K. An analytic model for description of temperature dependent rate phenomena in arthropods. Environ. Entomol. 1976, 5, 1133–1140. [Google Scholar] [CrossRef]
- Ikemoto, T. Intrinsic optimum temperature for development of insects and mites. Environ. Entomol. 2005, 34, 1377–1387. [Google Scholar] [CrossRef]
- Quinn, B.K. A critical review of the use and performance of different function types for modeling temperature-dependent development of arthropod larvae. J. Therm. Biol. 2017, 63, 65–77. [Google Scholar] [CrossRef]
- Régnier, B.; Legrand, J.; Rebaudo, F. Modeling temperature-dependent development rate in insects and implications of experimental design. Environ. Entomol. 2021, 51, 132–144. [Google Scholar] [CrossRef]
- Sevacherian, V.; Stern, V.M.; Mueller, A.J. Heat accumulation for timing Lygus control measures in a safflower-cotton complex. J. Econ. Entomol. 1976, 70, 399–402. [Google Scholar] [CrossRef]
- Harsimran, K.; Harsh, G. Pesticides: Environmental impacts and management strategies. In Pesticides-Toxic Aspects; Larramendy, M.L., Soloneski, S., Eds.; IntechOpen: London, UK, 2014. [Google Scholar]
- Joon, J.; Yeol, C.; Jung, C. Model of Grapholita molesta spring emergence in pear orchards based on statistical information criteria. J. Asia Pac. Entomol. 2012, 15, 589–593. [Google Scholar] [CrossRef]
- James, L.; Gary, L.; Street, G.; States, U.; Julie, C.; Carolina, N.; States, U. Degree-day model for timing insecticide applications to control Dioryctria amatella (Lepidoptera: Pyralidae ) in loblolly pine seed orchards. Can. Entomol. 2002, 134, 255–268. [Google Scholar]
- Rabhi, V. Modeling temperature-dependent development rate and phenology in insects: Review of major developments, challenges, and future directions. Entomol. Exp. Appl. 2018, 607–617. [Google Scholar] [CrossRef]
- Labatte, B.J.M. Modelling the larval development. J. Appl. Entomol. 1994, 118, 172–178. [Google Scholar] [CrossRef]
- Hai-, L.Z.; He, M.; Chen, C.; Guo, X. Effects of temperature on life-history traits of the newly invasive fall armyworm, Spodoptera frugiperda in Southeast China. Ecol. Evol. 2021, 5255–5264. [Google Scholar] [CrossRef]
- Prasad, T.V.; Srinivasa, R.M.; Rao, K.; Bal, S.K.; Muttapa, Y.; Choudhary, J.S.; Singh, V.K. Temperature-based phenology model for predicting the present and future establishment and distribution of recently invasive Spodoptera frugiperda (J. E. Smith) in India. Bull. Entomol. Res. 2021, 112, 1–15. [Google Scholar] [CrossRef]
- Du Plessis, H.; Schlemmer, M.L.; Van den Berg, J. The effect of temperature on the development of Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2020, 11. [Google Scholar] [CrossRef]
- Kadzamira, M.A.T.J.; Chaudhary, M.; Williams, F.; Dutta, N.K. A non-linear approach to the establishment of local biological control agent production units: A case study of fall armyworm in Bangladesh. CABI Agric. Biosci. 2022, 3, 48. [Google Scholar] [CrossRef]
- Maino, J.L.; Schouten, R.; Overton, K.; Day, R.; Ekesi, S.; Bett, B.; Barton, M.; Gregg, P.C.; Umina, P.A.; Reynolds, O.L. Regional and seasonal activity predictions for fall armyworm in Australia. Curr. Res. Insect Sci. 2021, 1, 100010. [Google Scholar] [CrossRef]
- Gilioli, G.; Colli, P.; Colturato, M.; Gervasio, P.; Sperandio, G. A nonlinear model for stage-structured population dynamics with nonlocal density-dependent regulation: An application to the fall armyworm moth. Math. Biosci. 2021, 335, 108573. [Google Scholar] [CrossRef]
- Acharya, R.; Hwang, H.S.; Mostafiz, M.M.; Yu, Y.S.; Lee, K.Y. Susceptibility of various developmental stages of the fall armyworm, Spodoptera frugiperda, to entomopathogenic nematodes. Insects 2020, 11, 868. [Google Scholar] [CrossRef]
- Malekera, M.J.; Acharya, R.; Hwang, H.S.; Lee, K.Y. Effect of cold acclimation and rapid cold-hardening on the survival of Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) under cold stress. J. Asia Pac. Entomol. 2022, 25, 101862. [Google Scholar] [CrossRef]
- Angulo, A.O.; Carmen, J.S. Pupae of the genus Spodoptera guenee, 1852, from the north of Chile (Lepidoptera: Noctuidae). Agric. Tech. 1982, 42, 347–349. [Google Scholar]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; De Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Developmental parameters of Spodoptera frugiperda (Lepidoptera: Noctuidae ) immature stages under controlled and standardized conditions. J. Agric. Sci. 2019, 11, 76–89. [Google Scholar] [CrossRef]
- Gotthard, K.; Nylin, S.; Wiklund, C. Adaptive variation in growth rate: Life history costs and consequences in the speckled wood butterfly, Pararge aegeria. Oecologia 1994, 99, 281–289. [Google Scholar] [CrossRef]
- Su, Y.; Li, X.; Zhang, W.; Zhang, B.; Ma, C. Sen Dynamics of heat shock protein responses to thermal stress changes after metamorphosis in a lepidopteran insect. Arch. Insect Biochem. Physiol. 2021, 107, e21791. [Google Scholar] [CrossRef]
- Agbodzavu, M.K.; Osiemo-Lagat, Z.; Gikungu, M.; Ekesi, S.; Fiaboe, K.K.M. Temperature-dependent development, survival and reproduction of Apanteles hemara (Nixon) (Hymenoptera: Braconidae) on Spoladea recurvalis (F.) (Lepidoptera: Crambidae). Bull. Entomol. Res. 2020, 110, 577–587. [Google Scholar] [CrossRef]
- Kaplan, E.L.; Meier, P. Nonparametric estimation from incomplete observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]
- Ali, A.; Luttrell, R.G.; Schneider, J.C. Effects of temperature and larval diet on development of the fall armyworm (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 1990, 83, 725–733. [Google Scholar] [CrossRef]
- Simmons, A.M. Effects of constant and fluctuating temperatures and humidities on the survival of Spodoptera frugiperda pupae (Lepidoptera: Noctuidae). Fla. Entomol. 1993, 76, 333–340. [Google Scholar] [CrossRef]
- Valdez-Torres, J.B.; Soto-Landeros, F.; Osuna-Enciso, T.; Báez-Sañudo, M.A. Phenological prediction models for white corn (Zea mays L.) and fall armyworm (Spodoptera frugiperda J.E. Smith). Agrociencia 2012, 46, 399–410. [Google Scholar]
- Dahi, H.F.; Salem, S.A.R.; Gamil, W.E.; Mohamed, H.O. Heat requirements for the fall armyworm Spodoptera frugiperda (J.E. Smith) as a new invasive pest in Egypt. Egypt. Acad. J. Biol. Sci. 2020, 13, 73–85. [Google Scholar] [CrossRef]
- Campbell, A.; Frazer, B.D.; Gilbert, N.; Gutierrez, A.P.; Mackauer, M. Temperature requirements of some Aphids and their parasites. J. Appl. Ecol. 1974, 11, 431–438. [Google Scholar] [CrossRef]
- Damos, P.T.; Savopoulou-Soultani, M. Temperature-dependent bionomics and modeling of Anarsia lineatella (Lepidoptera: Gelechiidae) in the laboratory. J. Econ. Entomol. 2008, 101, 1557–1567. [Google Scholar] [CrossRef]
- Briere, J.F.; Pracros, P.; Roux, A.Y.; Pierre, J.S. A novel rate model of temperature-dependent development for arthropods. Environ. Entomol. 1999, 28, 22–29. [Google Scholar] [CrossRef]
- Shi, P.; Ge, F. A comparison of different thermal performance functions describing temperature-dependent development rates. J. Therm. Biol. 2010, 35, 225–231. [Google Scholar] [CrossRef]
- Taylor, F. Ecology and Evolution of Physiological Time in Insects. Am. Nat. 1981, 117, 1–23. [Google Scholar] [CrossRef]
- Kontodima, D.; Eliopoulos, P..; Stathas, G..; Economou, L. Comparative temperature-dependent development of Nephus includens (Kirsch) and Nephus bisignatus (Boheman) (Coleoptera: Coccinellidae) Preying on Planococcus citri (Risso) (Homoptera: Pseudococcidae): Evaluation of a linear and various nonlinear models us. Environ. Entomol. 2004, 33, 1–11. [Google Scholar] [CrossRef]
- Marchioro, C.A.; Krechemer, S.; Foerster, L.A. Estimating the development rate of the tomato leaf miner, Tuta absoluta (Lepidoptera: Gelechiidae), using linear and non-linear models. Pest Manag. Sci. 2017, 73, 1486–1493. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, H.S.; Nuessly, G.S.; Webb, S.E.; Cherry, R.H.; Gilbert, R.A. Temperature-dependent development of Elasmopalpus lignosellus (Lepidoptera: Pyralidae) on sugarcane under laboratory conditions. Physiol. Ecol. 2010, 39, 1012–1020. [Google Scholar] [CrossRef]
- Symonds, M.R.E.; Moussalli, A. A brief guide to model selection, multimodel inference and model averaging in behavioural ecology using Akaike’s information criterion. Behav. Ecol. Sociobiol. 2011, 65, 13–21. [Google Scholar] [CrossRef]
- Karban, R.; Valpine, P.D. Population dynamics of an Arctiid caterpillar – tachinid parasitoid system using state-space models. J. Anim. Ecol. 2010, 650–661. [Google Scholar] [CrossRef] [PubMed]
- Marco, V.; Taberner, A.N.A.; Castanera, P. Development and survival of immature Aubeonymus mariaefranciscae (Coleoptera: Curculionidae) at constant temperatures. Ann. Entomol. Soc. Am 1997, 90, 169–176. [Google Scholar] [CrossRef]
- Barfield, C.S.; Mitchell, E.R.; Poe, S.L.B. A temperature-dependent model for Fall Armyworm development. Ann. Entomol. Soc. Am. 1978, 71, 70–74. [Google Scholar] [CrossRef]
- Hance, T.; Van Baaren, J.; Vernon, P.; Boivin, G. Impact of extreme temperatures on parasitoids in a climate change perspective. Annu. Rev. Entomol. 2007, 52, 107–260. [Google Scholar] [CrossRef]
- Jacques, J.; Sampaio, F.; Talissa, H.; Marchioro, C.A. Climate change and voltinism of Mythimna sequax: The location and choice of phenological models matter. Agric. For. Entomol. 2019, 21, 431–444. [Google Scholar] [CrossRef]
- Borchert, D.M.; Stinner, R.E.; Walgenbach, J.F.; Kennedy, G.G. Oriental fruit moth (Lepidoptera: Tortricidae) phenology and management with methoxyfenozide in North Carolina apples. J. Econ. Entomol. 2004, 97, 1353–1364. [Google Scholar] [CrossRef]
- dos Santos, H.T.; Marchioro, C.A. Selection of models to describe the temperature-dependent development of Neoleucinodes elegantalis (Lepidoptera: Crambidae) and its application to predict the species voltinism under future climate conditions. Bull. Entomol. Res. 2021, 111, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, F.; Dos Santos, H.T.; Marchioro, C.A. Differences in thermal tolerances between two soybean pests may differently affect their voltinism under climate change. Agric. For. Entomol. 2022, 24, 380–389. [Google Scholar] [CrossRef]
- Marchioro, C.A.; Krechemer, F.S.; Moraes, C.P.; Foerster, L.A. A stochastic model for predicting the stage emergence of Plutella xylostella under field conditions. Ann. Appl. Biol. 2016, 169, 190–199. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Model | Function | Reference |
|---|---|---|
| linear | D(T) = a + bT | [38] |
| β type | D(T) = ρ.(aT/10).(T/10)β | [39] |
| Briere-1 | D(T) = aT(T − TL)(TH − T)1/2 | [40] |
| Briere-2 | D(T) = aT(T − TL)(TH − T)1/m | [40] |
| Shi | D(T) = m(T − TL(1− ) | [41] |
| Logan-6 | D(T) = Ψ( | [10] |
| Taylor | D(T) = Rm.EXP − 0.5(T − Topt)2 | [42] |
| Temperature (°C) | N * | Egg † | Larvae | Pupae | Egg-to-Adult | ||
|---|---|---|---|---|---|---|---|
| Male | Female | Mean | |||||
| 15 | 80 | 14.25 ± 1.31 a | 87.42 ± 2.68 a | - | - | - | - |
| 20 | 87 | 5.33 ± 0.88 b | 25.41 ± 0.93 ab | 21.85 ± 0.45 a | 19.6 ± 1.43 a | 20.91 ± 0.70 a | 50.46 ± 1.71 a |
| 25 | 70 | 2.29 ± 0.17 bc | 15.37 ± 0.84 ab | 10.25 ± 0.25 ab | 9.57 ± 0.12 ab | 9.72 ± 0.125 ab | 31.88 ± 0.25 ab |
| 28 | 103 | 2.00 ± 0.00 c | 13.61 ± 0.22 bc | 8.75 ± 0.18 b | 7.55 ± 0.16 b | 8.15 ± 0.15 b | 25.22 ± 0.58 abc |
| 30 | 90 | 1.50 ± 0.28 c | 12.64 ± 0.31 c | 7.87 ± 0.39 bc | 7.40 ± 0.63 bc | 7.58 ± 0.57 bc | 23.44 ±1.27 bc |
| 32 | 80 | 1.33 ± 0.33 c | 9.11 ± 0.30 c | 7.12 ± 0.21 bc | 6.62 ± 0.17 bc | 6.87 ± 0.14 bc | 23.43 ± 0.38 c |
| 34 | 80 | 2.36 ± 0.06 c | 13.11 ± 0.34 c | 6.83 ± 0.40 c | 5.81 ± 0.23 c | 6.06 ± 0.21 c | 23.87 ± 0.42 c |
| Temperature (°C) | Pupal Weight (mg) | Growth Rate (mg/Day) | ||
|---|---|---|---|---|
| Male | Female | Male | Female | |
| 20 | 194.42 ± 3.74 aB (20) † | 202.12 ± 6.15 aA (23) † | 8.31 ± 0.31 c | 9.26 ± 0.87 d |
| 25 | 142.98 ± 9.27 bA (13) | 139.53 ± 4.10 bB (18) | 8.32 ± 0.31 c | 9.51 ± 0.71 d |
| 28 | 193.96 ± 10.88 aA (52) | 192.06 ± 6.30 aB (48) | 14.59 ± 0.77 a | 13.87 ± 0.38 a |
| 30 | 167.26 ± 7.90 bA (25) | 150.24 ± 5.73 bB (33) | 13.13 ± 1.25 a | 14.68 ± 1.4 a |
| 32 | 126.43 ± 4.65 cA (16) | 120.54 ± 4.72 cB (13) | 12.11 ± 0.98 b | 10.94 ± 0.67 b |
| 34 | 127.35 ± 14.95 cA (16) | 117.80 ± 6.05 cB (15) | 10.79 ± 0.97 b | 9.75 ± 0.72 c |
| Model | Egg | Larvae | Pupae | Egg–Adult | ||||
|---|---|---|---|---|---|---|---|---|
| R2 | ∆AICc | R2 | ∆AICc | R2 | ∆AICc | R2 | ∆AICc | |
| Linear | 0.99 | 0.00 | 0.91 | 8.99 | 0.97 | 7.13 | 0.96 | 0.00 |
| β type | 0.81 | 8.29 | 0.71 | 8.91 | 0.86 | 20.93 | 0.74 | 11.29 |
| Briere-1 | 0.91 | 4.46 | 0.95 | 0.00 | 1.00 | 2.89 | 0.94 | 4.17 |
| Briere-2 | 0.86 | 6.68 | 0.95 | 0.00 | 0.98 | 9.91 | 0.94 | 4.17 |
| Shi | 0.88 | 5.96 | 0.86 | 5.21 | 0.99 | 0.00 | 0.95 | 0.99 |
| Logan | 0.89 | 5.54 | 0.88 | 4.33 | 0.93 | 17.59 | 0.91 | 5.97 |
| Taylor | 0.96 | 0.25 | 0.95 | 0.10 | 0.99 | 8.93 | 0.97 | 0.13 |
| Model | Egg | Larvae | Pupae | Egg–Adult |
|---|---|---|---|---|
| Linear | ||||
| a (10−2) | 0.00 | −6.60 | −10.64 | −1.99 |
| b (10−2) | 4.70 | 0.52 | 0.80 | 0.20 |
| TL | 15.92 | 12.78 | 13.26 | 9.73 |
| β-type | ||||
| ρ (10−3) | 15.88 | 3.70 | 2.72 | 1.67 |
| a | 3.89 | 4.04 | 3.90 | 4.04 |
| b | 3.31 | 2.64 | 3.58 | 2.71 |
| TH | 38.86 | 40.36 | 39.01 | 40.33 |
| Topt | 30.28 | 30.10 | 31.09 | 30.13 |
| Briere-1 | ||||
| a (10−5) | 16.12 | 2.15 | 2.29 | 1.14 |
| TL | 12.23 | 12.88 | 13.47 | 13.08 |
| TH | 43.50 | 45.31 | 54.14 | 44.42 |
| Topt | 33.82 | 33.19 | 33.58 | 32.77 |
| Briere-2 | ||||
| a (10−6) | 89.54 | 45.39 | 32.38 | 24.80 |
| m | 7.27 | 3.19 | 3.04 | 4.35 |
| TL | 14.96 | 9.94 | 9.77 | 13.09 |
| TH | 40.90 | 39.53 | 51.04 | 44.41 |
| Topt | 31.21 | 30.35 | 36.74 | 32.99 |
| Shi | ||||
| K | 14.16 | 3.51 | 2.60 | 12.79 |
| m (10−3) | 41.00 | 5.16 | 9.03 | 2.56 |
| TL | 14.27 | 12.78 | 14.68 | 13.51 |
| TH | 34.08 | 34.33 | 37.58 | 34.13 |
| Topt | 33.91 | 33.62 | 36.35 | 33.95 |
| Logan-6 | ||||
| Ρ | 0.10 | 0.10 | 0.11 | 0.10 |
| Δ | 9.12 | 9.56 | 8.96 | 9.07 |
| Ψ | 0.22 | 0.18 | 0.13 | 0.01 |
| TH | 40.61 | 40.63 | 40.71 | 41.09 |
| Topt | 32.12 | 32.00 | 32.43 | 32.13 |
| Taylor | ||||
| Rm (10−2) | 72.49 | 8.87 | 15.75 | 4.18 |
| TL | 6.50 | 8.83 | 9.03 | 10.41 |
| Topt | 32.74 | 32.04 | 34.38 | 33.38 |
| Life Stage | Regression Parameters | |||||
|---|---|---|---|---|---|---|
| Equation | Slope ± SEM | Intercept | R2 | K (Degree-Days) | TL (°C) | |
| Egg | Y = 0.04696 * X − 0.7474 | 0.04696 ± 0.0009017 | −0.7474 ± 0.02464 | 0.999 | 21.29 | 15.92 |
| Total larval | Y = 0.00516 * X − 0.06596 | 0.00516 ± 0.0009558 | −0.06596 ± 0.02612 | 0.907 | 193.80 | 12.78 |
| Pupa | Y = 0.00802 * X − 0.1064 | 0.00802 ± 0.0007856 | −0.1064 ± 0.02146 | 0.972 | 124.70 | 13.27 |
| Egg–adult | Y = 0.002041 * X − 0.01986 | 0.002041 ± 0.0002483 | −0.01986 ± 0.006786 | 0.958 | 490.00 | 9.73 |
| Model | Number of Estimated Thermal Threshold | Accuracy ª | ||
|---|---|---|---|---|
| TL | Topt | TH | ||
| Linear | 1 | + | * | * |
| β type | 2 | * | + | − |
| Briere-1 | 3 | + | + | − |
| Briere-2 | 3 | + | + | − |
| Lactine-2 | 3 | − | + | − |
| Shi | 3 | + | + | + |
| Logan-6 | 2 | * | + | − |
| Taylor | 2 | + | + | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malekera, M.J.; Acharya, R.; Mostafiz, M.M.; Hwang, H.-S.; Bhusal, N.; Lee, K.-Y. Temperature-Dependent Development Models Describing the Effects of Temperature on the Development of the Fall Armyworm Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae). Insects 2022, 13, 1084. https://doi.org/10.3390/insects13121084
Malekera MJ, Acharya R, Mostafiz MM, Hwang H-S, Bhusal N, Lee K-Y. Temperature-Dependent Development Models Describing the Effects of Temperature on the Development of the Fall Armyworm Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae). Insects. 2022; 13(12):1084. https://doi.org/10.3390/insects13121084
Chicago/Turabian StyleMalekera, Matabaro Joseph, Rajendra Acharya, Md Munir Mostafiz, Hwal-Su Hwang, Narayan Bhusal, and Kyeong-Yeoll Lee. 2022. "Temperature-Dependent Development Models Describing the Effects of Temperature on the Development of the Fall Armyworm Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae)" Insects 13, no. 12: 1084. https://doi.org/10.3390/insects13121084
APA StyleMalekera, M. J., Acharya, R., Mostafiz, M. M., Hwang, H.-S., Bhusal, N., & Lee, K.-Y. (2022). Temperature-Dependent Development Models Describing the Effects of Temperature on the Development of the Fall Armyworm Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae). Insects, 13(12), 1084. https://doi.org/10.3390/insects13121084