

Molecular Detection of Pentastiridius leporinus, the Main Vector of the Syndrome ‘Basses Richesses’ in Sugar Beet


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Planthopper Collection and Morphological Identification
2.2. Template DNA Preparation
2.3. Primer Design and PCR Conditions
2.4. Application to Adult and Immature Specimens
2.5. Evolutionary Relationships
3. Results
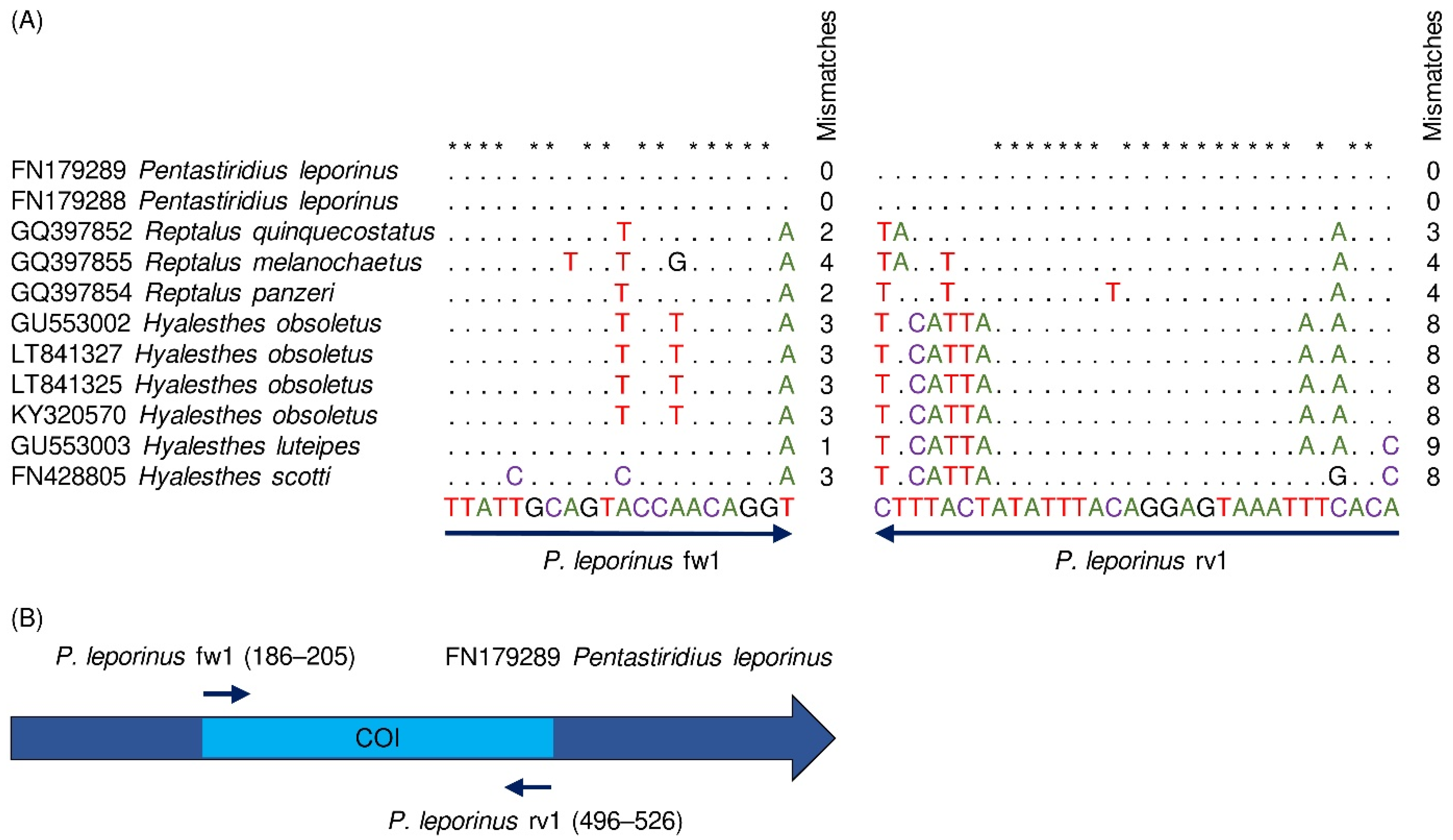
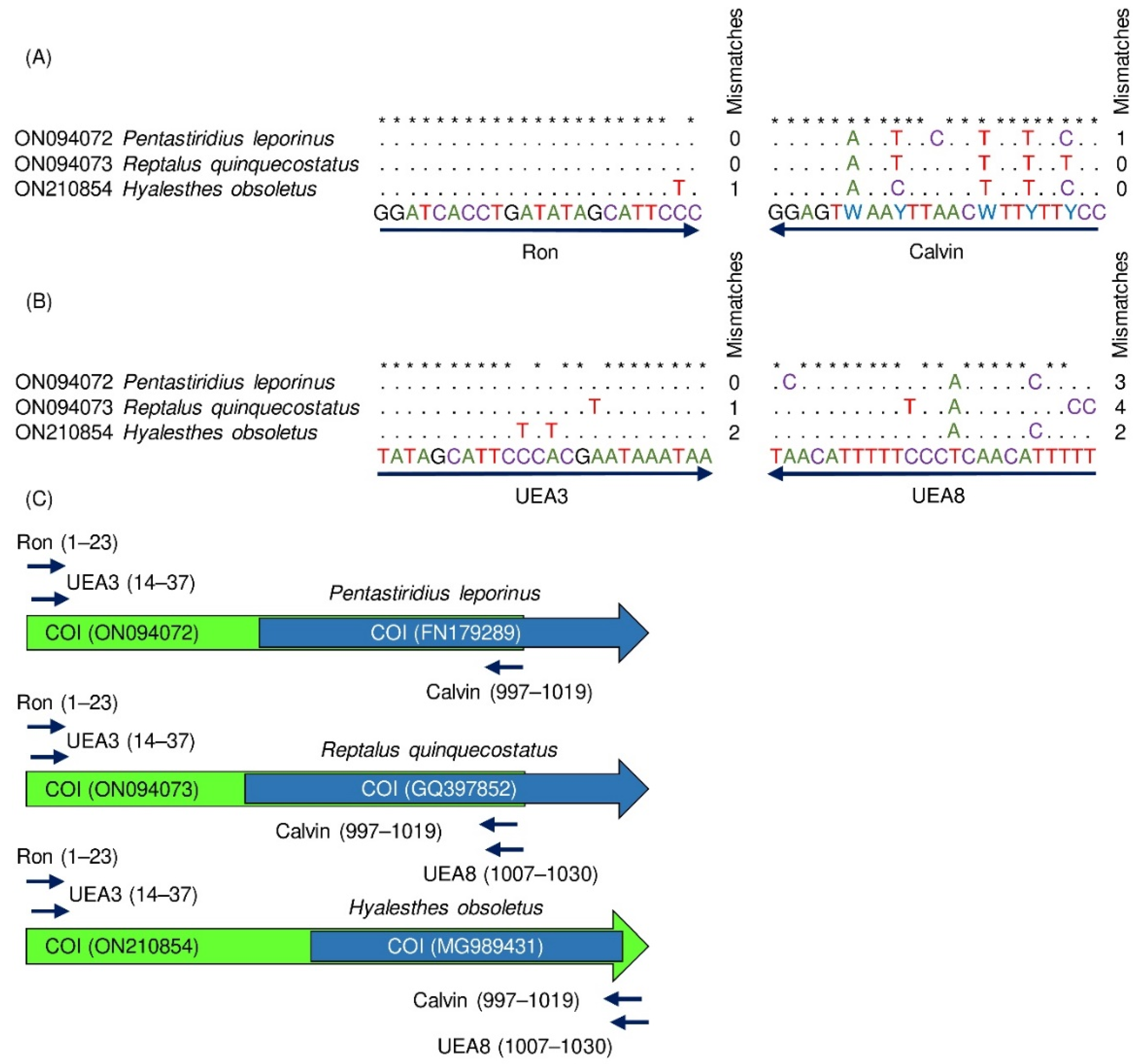
3.1. Species-Specific Primer Design
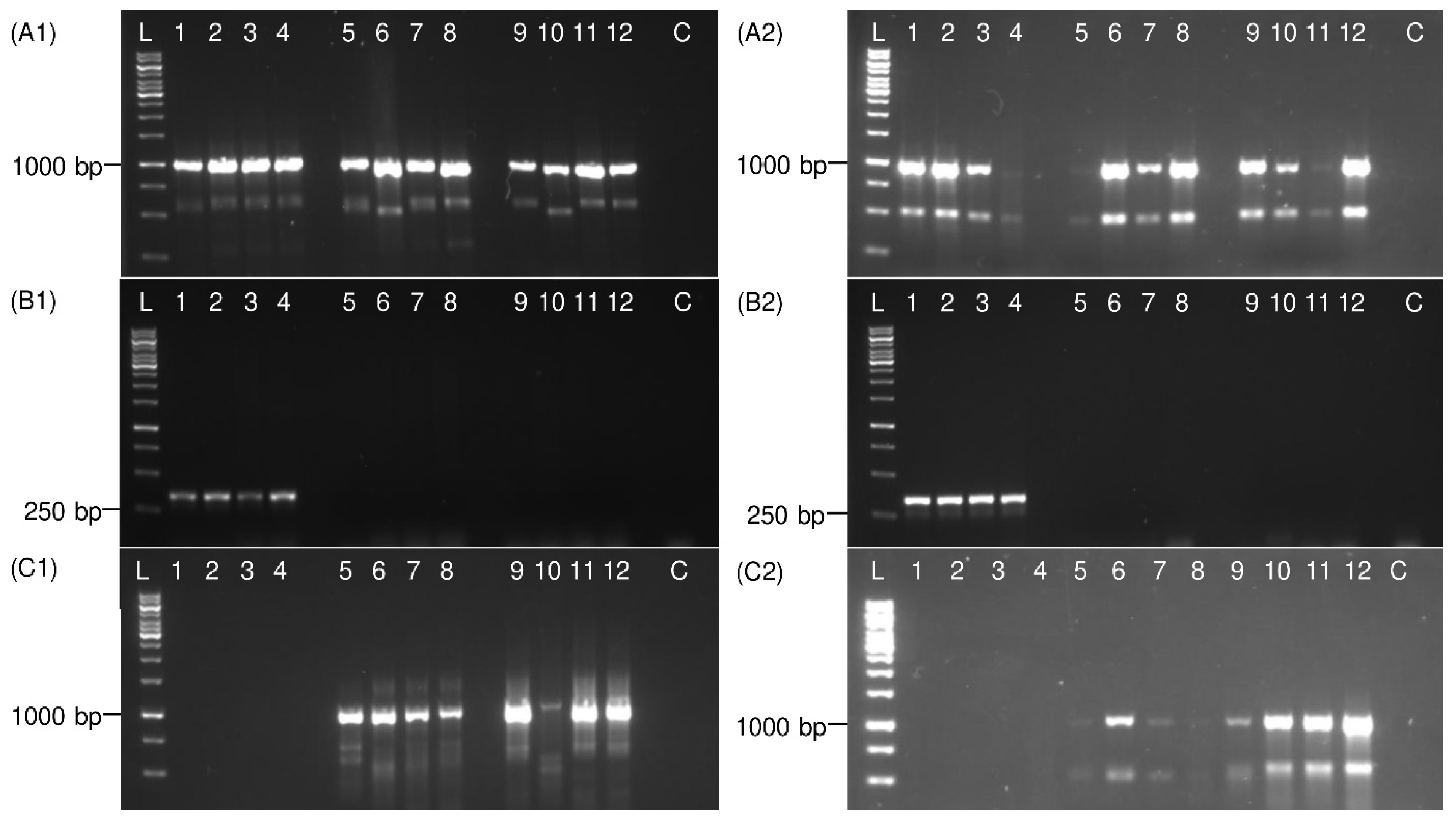
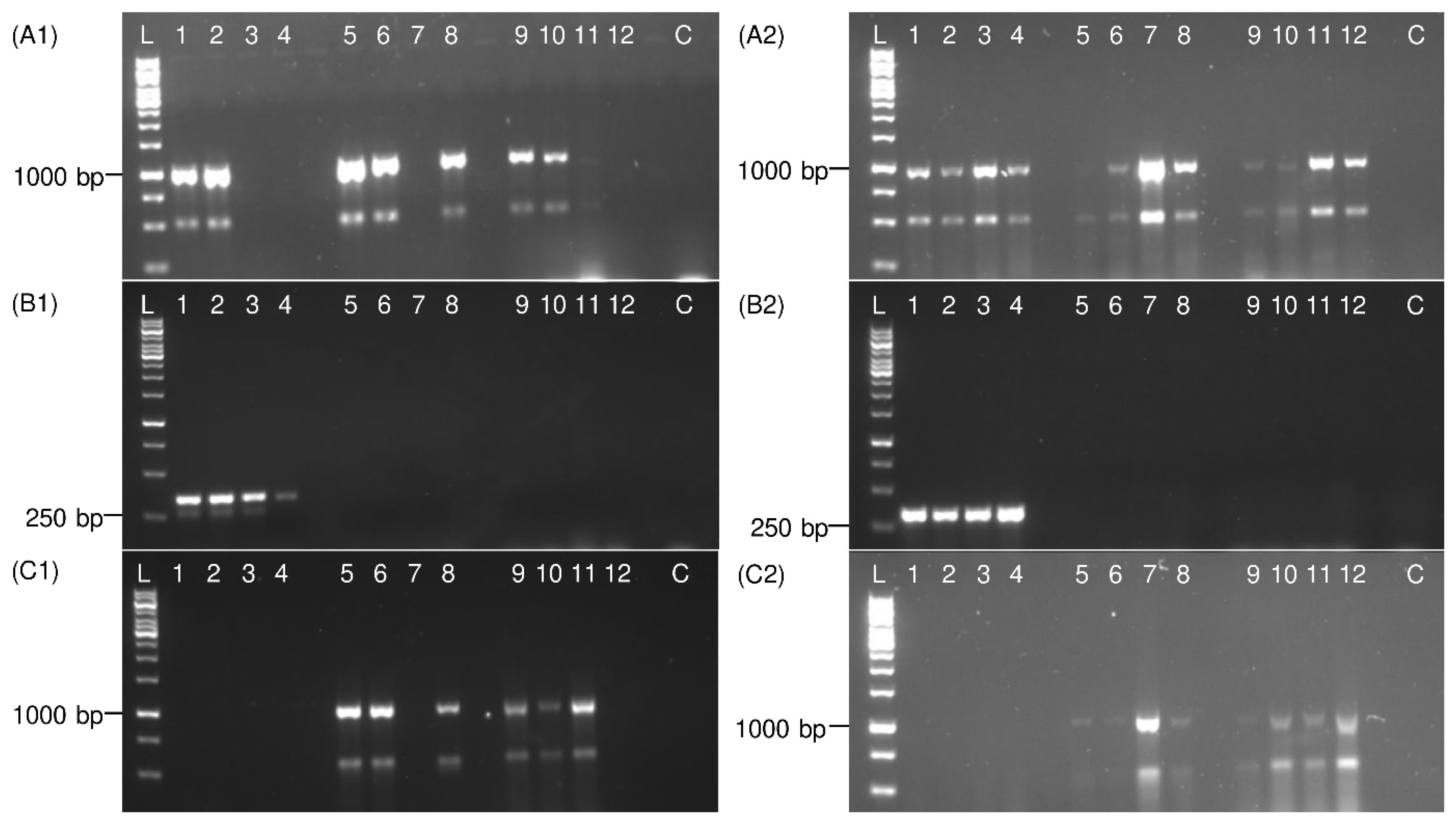
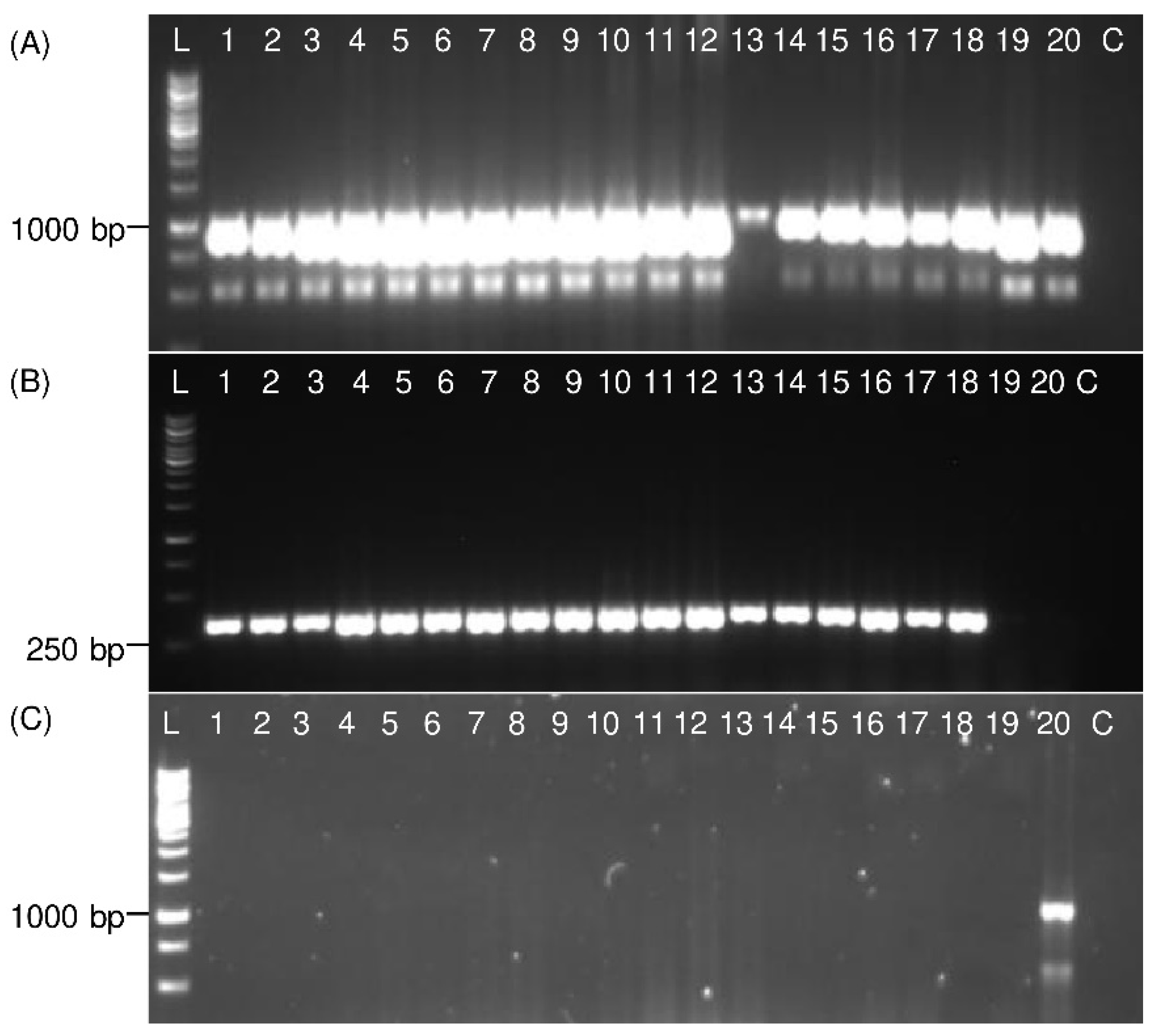
3.2. PCR Validation on Adult Planthoppers
3.3. Detection of Immature Life Stages of P. leporinus
3.4. Detection of Distantly Related Species from Sugar Beet Fields
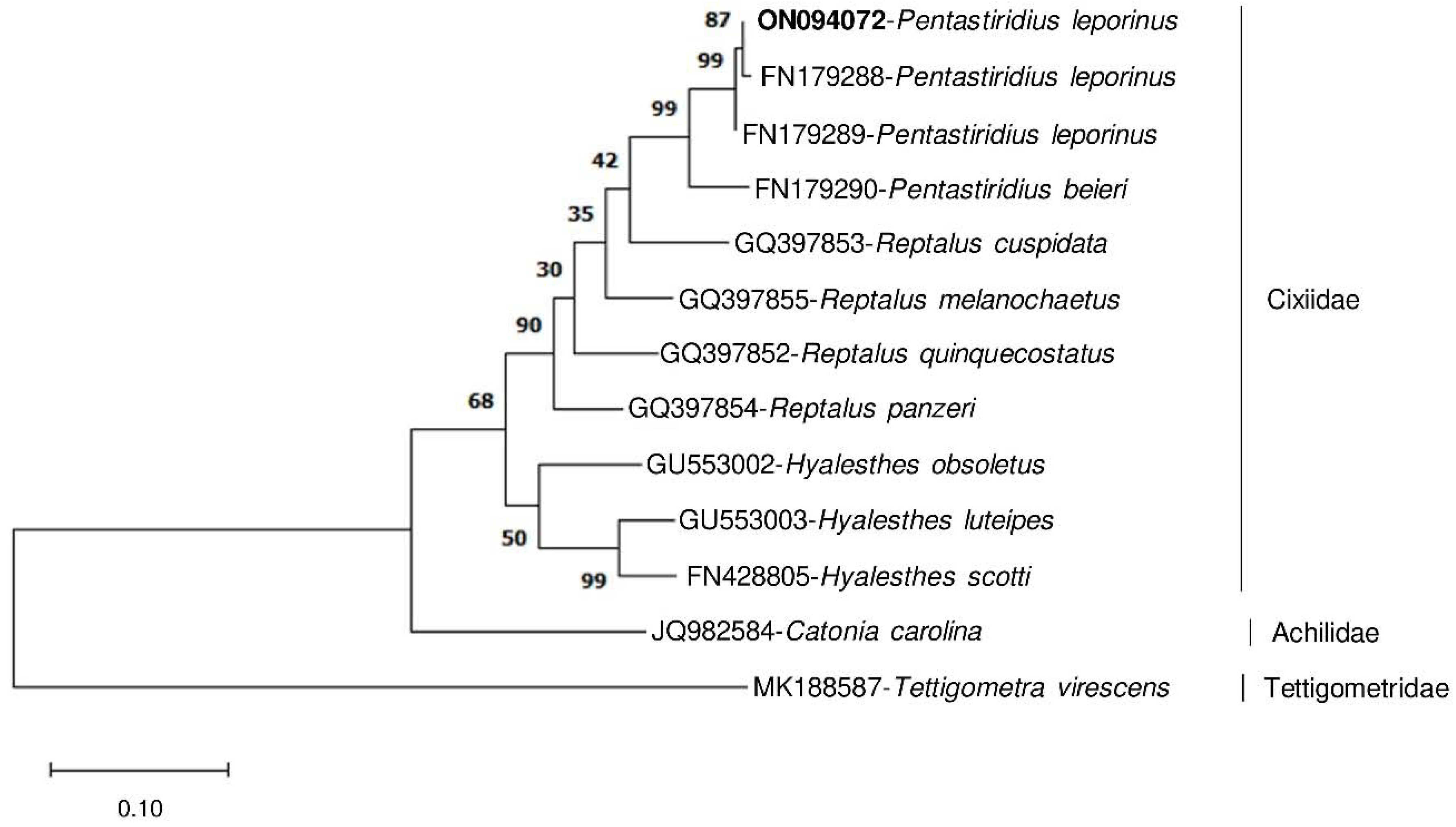
3.5. Evolutionary Relationships
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bressan, A.; Sémétey, O.; Nusillard, B.; Clair, D.; Boudon-Padieu, E. Insect Vectors (Hemiptera: Cixiidae) and Pathogens Associated with the Disease Syndrome “Basses Richesses” of Sugar Beet in France. Plant. Dis. 2008, 92, 113–119. [Google Scholar] [CrossRef]
- Gatineau, F.; Jacob, N.; Vautrin, S.; Larrue, J.; Lherminier, J.; Richard-Molard, M.; Boudon-Padieu, E. Association with the Syndrome “Basses Richesses” of Sugar Beet of a Phytoplasma and a Bacterium-Like Organism Transmitted by a Pentastiridius sp. Phytopathology 2002, 92, 384–392. [Google Scholar] [CrossRef]
- Sémétey, O.; Bressan, A.; Richard-Molard, M.; Boudon-Padieu, E. Monitoring of proteobacteria and phytoplasma in sugar beets naturally or experimentally affected by the disease syndrome ‘Basses richesses’. Eur. J. Plant. Pathol. 2007, 117, 187–196. [Google Scholar] [CrossRef]
- Sémétey, O.; Gatineau, F.; Bressan, A.; Boudon-Padieu, E. Characterization of a gamma-3 Proteobacteria Responsible for the Syndrome “Basses Richesses” of Sugar Beet Transmitted by Pentastiridius sp. (Hemiptera, Cixiidae). Phytopathology 2007, 97, 72–78. [Google Scholar] [CrossRef]
- Pfitzer, R.; Schrameyer, K.; Voegele, R.T.; Maier, J.; Lang, C.; Varrelmann, M. Ursachen und Auswirkungen des Auftretens von “Syndrome des basses richesses” in deutschen Zuckerrübenanbaugebieten. Sugar Ind. 2020, 145, 234–244. [Google Scholar]
- Behrmann, S.; Schwind, M.; Schieler, M.; Vilcinskas, A.; Martinez, O.; Lee, K.-Z.; Lang, C. Spread of bacterial and virus yellowing diseases of sugar beet in South and Central Germany from 2017–2020. Sugar Ind. 2021, 146, 476–485. [Google Scholar] [CrossRef]
- Peter, M. Lichtblicke bei den Zuckerrüben. UFA-Revue. Available online: https://www.ufarevue.ch/pflanzenbau/ackerkulturen/lichtblicke-bei-den-zuckerrueben (accessed on 29 June 2022).
- Bressan, A.; Moral García, F.J.; Boudon-Padieu, E. The prevalence of ‘Candidatus Arsenophonus phytopathogenicus’ infecting the planthopper Pentastiridius leporinus (Hemiptera: Cixiidae) increase nonlinearly with the population abundance in sugar beet fields. Environ. Entomol. 2011, 40, 1345–1352. [Google Scholar] [CrossRef]
- Firrao, G.; Gibb, K.S.; Streten, C. Short taxonomic guide to the genus ‘Candidatus Phytoplasma’. J. Plant. Pathol. 2005, 87, 249–263. [Google Scholar]
- Gatineau, F.; Larrue, J.; Clair, D.; Lorton, F.; Richard-Molard, M.; Boudon-Padieu, E. A New Natural Planthopper Vector of Stolbur Phytoplasma in the Genus Pentastiridius (Hemiptera: Cixiidae). Eur. J. Plant. Pathol. 2001, 107, 263–271. [Google Scholar] [CrossRef]
- Sémétey, O.; Bressan, A.; Gatineau, F.; Boudon-Padieu, E. Development of a specific assay using RISA for detection of the bacterial agent of ‘basses richesses’ syndrome of sugar beet and confirmation of a Pentastiridius sp. (Fulgoromopha, Cixiidae) as the economic vector. Plant. Pathol. 2007, 56, 797–804. [Google Scholar] [CrossRef]
- Bressan, A.; Sémétey, O.; Arneodo, J.; Lherminier, J.; Boudon-Padieu, E. Vector transmission of a plant-pathogenic bacterium in the Arsenophonus clade sharing ecological traits with facultative insect endosymbionts. Ecol. Epidemiol. 2009, 99, 1289–1296. [Google Scholar] [CrossRef] [PubMed]
- Pfitzer, R.; Varrelmann, M.; Schrameyer, K.; Rostás, M. Life history traits and a method for continuous mass rearing of the planthopper Pentastiridius leporinus, a vector of the causal agent of syndrome “basses richesses” in sugar beet. Pest. Manag. Sci. 2022, 78, 4700–4708. [Google Scholar] [CrossRef] [PubMed]
- Holzinger, W.E.; Kammerlander, I.; Nickel, H. The Auchenorrhyncha of Central Europe = Die Zikaden Mitteleuropas; Brill: Leiden, The Netherlands, 2003. [Google Scholar]
- Bressan, A. Agronomic practices as potential sustainable options for the management of Pentastiridius leporinus (Hemiptera: Cixiidae) in sugar beet crops. J. Appl. Entomol. 2009, 133, 760–766. [Google Scholar] [CrossRef]
- Bressan, A.; Holzinger, W.E.; Nusillard, B.; Sémétéy, O.; Gatineau, F.; Simonato, M.; Boudon-Padieu, E. Identification and biological traits of a planthopper from the genus Pentastiridius (Hemiptera: Cixiidae) adapted to an annual cropping rotation. Eur. J. Entomol. 2009, 106, 405–413. [Google Scholar] [CrossRef]
- Pfitzer, R. Untersuchung zur Verbreitung und Kontrolle von SBR in Zuckerrüben. M.Sc. thesis, University of Hohenheim, Stuttgart, Germany, February 2019. [Google Scholar]
- Pfitzer, R.; Schrameyer, K.; Varrelmann, M. Planthopper collection in sugar beet fields. Unpublished work. 2020. [Google Scholar]
- Maixner, M.; Ahrens, U.; Seemueller, E. Detection of the German grapevine yellows (Vergilbungskrankheit) MLO in grapevine, alternative hosts and a vector by a specific PCR procedure. Eur. J. Plant Pathol. 1995, 241–250. [Google Scholar] [CrossRef]
- Šafářová, D.; Lauterer, P.; Starý, M.; Válová, P.; Navrátil, M. Insight into epidemiological importance of phytoplasma vectors in vineyards in South Moravia, Czech Republic. Plant. Protect. Sci. 2018, 54, 234–239. [Google Scholar]
- Sforza, R.; Clair, D.; Daire, X.; Larrue, J.; Boudon-Padieu, E. The Role of Hyalesthes obsoletus (Hemiptera: Cixiidae) in the Occurrence of Bois noir of Grapevines in France. J. Phytopathol. 1998, 146, 549–556. [Google Scholar] [CrossRef]
- Mitrović, M.; Jakovljević, M.; Jović, J.; Krstić, O.; Kosovac, A.; Trivellone, V.; Jermini, M.; Toševski, I.; Cvrković, T. ‘Candidatus phytoplasma solani’ genotypes associated with potato stolbur in Serbia and the role of Hyalesthes obsoletus and Reptalus panzeri (hemiptera, cixiidae) as natural vectors. Eur. J. Plant. Pathol. 2015, 144, 619–630. [Google Scholar] [CrossRef]
- Sémétey, O.; Gaudin, J.; Danet, J.-L.; Salar, P.; Theil, S.; Fontaine, M.; Krausz, M.; Chaisse, E.; Eveillard, S.; Verdin, E.; et al. Lavender decline in France is associated with chronic infection by lavender-specific strain of “Candidatus Phytoplasma solani”. Appl. Environ. Microb. 2018, 84, e01507–e01518. [Google Scholar] [CrossRef]
- Mori, N.; Mitrović, J.; Smiljković, M.; Duduk, N.; Paltrinieri, S.; Bertaccini, A.; Duduk, B. Hyalesthes obsoletus in Serbia and its role in the epidemiology of corn reddening. B. Insectol. 2013, 66, 245–250. [Google Scholar]
- Ceotto, P.; Bourgoin, T. Insights into the phylogenetic relationships within Cixiidae (Hemiptera: Fulgoromorpha): Cladistic analysis of a morphological dataset. Syst. Entomol. 2008, 33, 484–500. [Google Scholar] [CrossRef]
- Stewart, A.J.A. Techniques for sampling Auchenorrhyncha in grasslands. Denisia 2002, 4, 491–512. [Google Scholar]
- Bertin, S.; Picciau, L.; Ács, Z.; Alma, A.; Bosco, D. Molecular differentiation of four Reptalus species (Hemiptera: Cixiidae). Bull. Entomol. Res. 2010, 100, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Luo, J.; Liu, R.; Zhang, C.; Duan, D.; Chen, H.; Bei, W.; Tang, J. Identification of Nilaparvata lugens and Its Two Sibling Species (N. bakeri and N. muiri) by Direct Multiplex PCR. J. Econ. Entomol. 2018, 111, 2869–2875. [Google Scholar] [CrossRef] [PubMed]
- Bertin, S.; Picciau, L.; Ács, Z.; Alma, A.; Bosco, D. Molecular identification of the Hyalesthes species (Hemiptera: Cixiidae) occurring in vineyard agroecosystems. Ann. Appl. Biol. 2010, 157, 435–445. [Google Scholar] [CrossRef]
- Biedermann, R.; Niedringhaus, R. Die Zikaden Deutschlands: Bestimmungstafeln für alle Arten; Wissenschaftlicher Akademischer Buchvertrieb: Kleve, Germany, 2004. [Google Scholar]
- Virgilio, M.; Backeljau, T.; Nevado, B.; de Meyer, M. Comparative performances of DNA barcoding across insect orders. BMC. Bioinform. 2010, 11, 206. [Google Scholar] [CrossRef]
- Wilson, J.J. DNA Barcodes for Insects. In DNA Barcodes: Methods and Protocols; Kress, W.J., Erickson, D.L., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2012; Volume 858, pp. 17–46. [Google Scholar]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Bressan, A.; Arneodo, J.; Simonato, M.; Haines, W.P.; Boudon-Padieu, E. Characterization and evolution of two bacteriome-inhabiting symbionts in cixiid planthoppers (Hemiptera: Fulgoromorpha: Pentastirini). Environ. Microbiol. 2009, 11, 3265–3279. [Google Scholar] [CrossRef]
- Jangra, S.; Baranwal, V.K.; Dietzgen, R.G.; Ghosh, A. A rapid field-based assay using recombinase polymerase amplification for identification of Thrips palmi, a vector of tospoviruses. J. Pest. Sci. 2021, 94, 219–229. [Google Scholar]
- BioEdit 7.2. Available online: https://bioedit.software.informer.com/7.2/ (accessed on 27 September 2022).
- Argüello Caro, E.B.; Dumón, A.D.; Mattio, M.F.; Alemandri, V.; Truol, G. A molecular framework for the identification of planthopper vectors (Hemiptera: Delphacidae) of central Argentina. Bull. Entomol. Res. 2015, 105, 754–762. [Google Scholar] [CrossRef]
- Lunt, D.H.; Zhang, D.X.; Szymura, J.M.; Hewitt, G.M. The insect cytochrome oxidase I gene: Evolutionary patterns and conserved primers for phylogenetic studies. Insect. Mol. Biol. 1996, 5, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. P. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Ceotto, P.; Kergoat, G.J.; Rasplus, J.-Y.; Bourgoin, T. Molecular phylogenetics of cixiid planthoppers (Hemiptera: Fulgoromorpha): New insights from combined analyses of mitochondrial and nuclear genes. Mol. Phylogenet. Evol. 2008, 48, 667–678. [Google Scholar] [CrossRef]
- Urban, J.M.; Bartlett, C.R.; Cryan, J.R. Evolution of Delphacidae (Hemiptera: Fulgoroidea): Combined-evidence phylogenetics reveals importance of grass host shifts. Syst. Entomol. 2010, 35, 678–691. [Google Scholar] [CrossRef]
- Yashiro, T.; Sanada-Morimura, S. A rapid multiplex PCR assay for species identification of Asian rice planthoppers (Hemiptera: Delphacidae) and its application to early-instar nymphs in paddy fields. PLoS ONE 2021, 16, e0250471. [Google Scholar] [CrossRef]
- Chen, M.N.; Santander, R.D.; Talamas, E.J.; Jentsch, P.J.; Bon, M.-C.; Aćimović, S.G. Molecular Identification of Trissolcus japonicus, Parasitoid of the Brown Marmorated Stink Bug, by Species-Specific PCR. Insects 2021, 12, 467. [Google Scholar] [CrossRef]
- Hao, S.; Wang, H.; Tao, W.; Wang, J.; Zhang, Z.; Zhang, Q.; Zhang, M.; Guo, L.; Shi, X. Multiplex-PCR for Identification of Two Species in Genus Hishimonus (Hemiptera: Cicadellidae) in Jujube Orchards. J. Econ. Entomol. 2015, 108, 2443–2449. [Google Scholar] [CrossRef]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, Weighting, and Phylogenetic Utility of Mitochondrial Gene Sequences and a Compilation of Conserved Polymerase Chain Reaction Primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Lin, P.; Wood, T.K. Molecular Phylogeny of the North American Enchenopa binotata (Homoptera: Membracidae) Species Complex. Ann. Entomol. Soc. Am. 2002, 95, 162–171. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′ to 3′) | PCR-Product Size [bp] | Reference |
|---|---|---|---|
| Ron (fw) | GGATCACCTGATATAGCATTCCC | ~1000 | Argüello Caro [37] |
| Calvin (rv) | GGRAARAAWGTTAARTTWACTCC | ||
| P. leporinus fw1 | TTATTGCAGTACCAACAGGT | 341 | This study |
| P. leporinus rv1 | TGTGAAATTTACTCCTGTAAATATAGTAAAG | ||
| UEA3 (fw) | TATAGCATTCCCACGAATAAATAA | ~1000 | Lunt et al. [38] |
| UEA8 (rv) | AAAAATGTTGAGGGAAAAATGTTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pfitzer, R.; Varrelmann, M.; Hesse, G.; Eini, O. Molecular Detection of Pentastiridius leporinus, the Main Vector of the Syndrome ‘Basses Richesses’ in Sugar Beet. Insects 2022, 13, 992. https://doi.org/10.3390/insects13110992
Pfitzer R, Varrelmann M, Hesse G, Eini O. Molecular Detection of Pentastiridius leporinus, the Main Vector of the Syndrome ‘Basses Richesses’ in Sugar Beet. Insects. 2022; 13(11):992. https://doi.org/10.3390/insects13110992
Chicago/Turabian StylePfitzer, René, Mark Varrelmann, Georgia Hesse, and Omid Eini. 2022. "Molecular Detection of Pentastiridius leporinus, the Main Vector of the Syndrome ‘Basses Richesses’ in Sugar Beet" Insects 13, no. 11: 992. https://doi.org/10.3390/insects13110992
APA StylePfitzer, R., Varrelmann, M., Hesse, G., & Eini, O. (2022). Molecular Detection of Pentastiridius leporinus, the Main Vector of the Syndrome ‘Basses Richesses’ in Sugar Beet. Insects, 13(11), 992. https://doi.org/10.3390/insects13110992