

Effects of Color and Light Intensity on the Foraging and Oviposition Behavior of Culex pipiens biotype molestus Mosquitoes

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mosquito Colony
2.2. Colors
2.3. Bioassays
2.3.1. Oviposition Bioassay
2.3.2. Foraging Bioassay
2.3.3. Statistical Analyses
3. Results
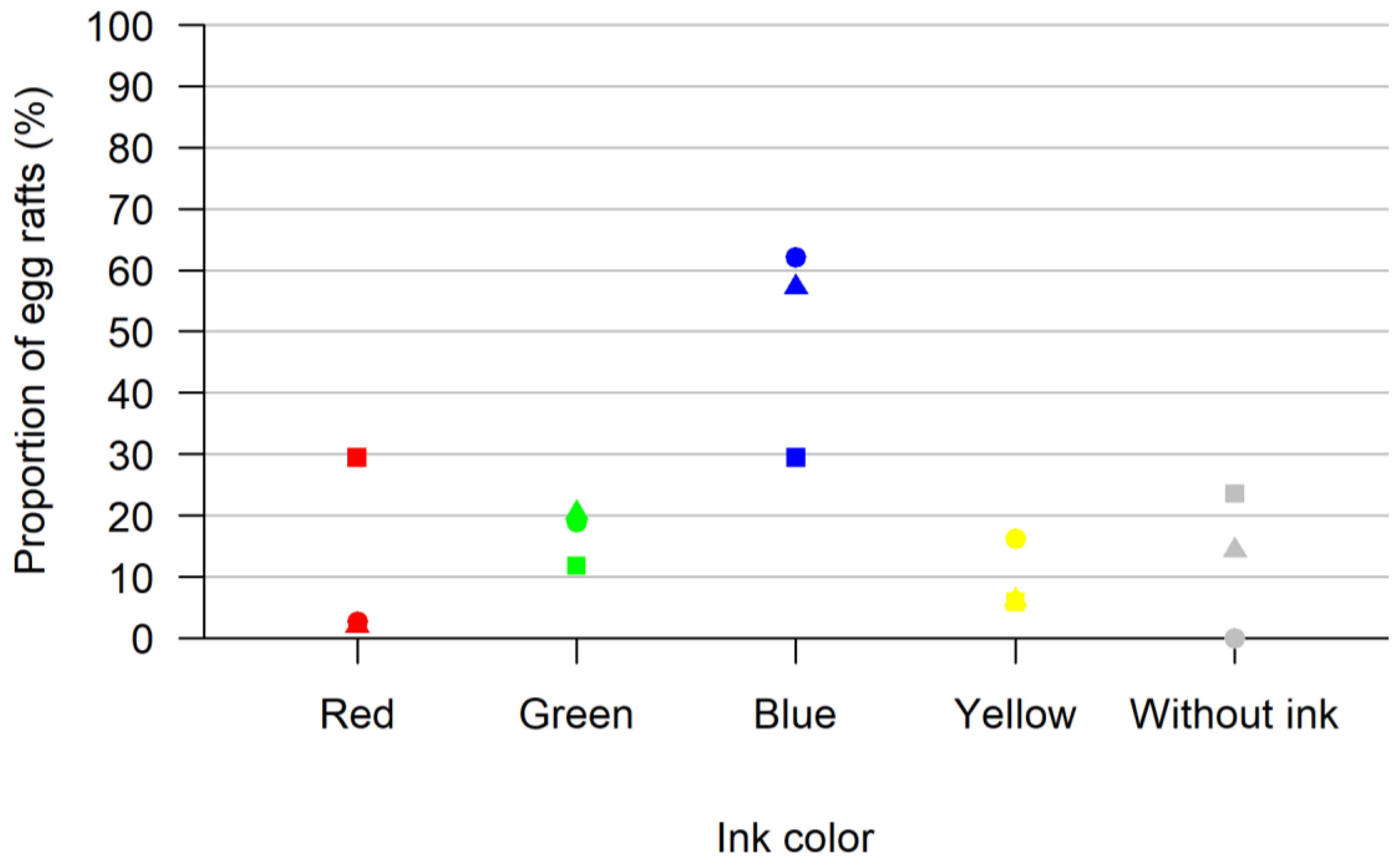
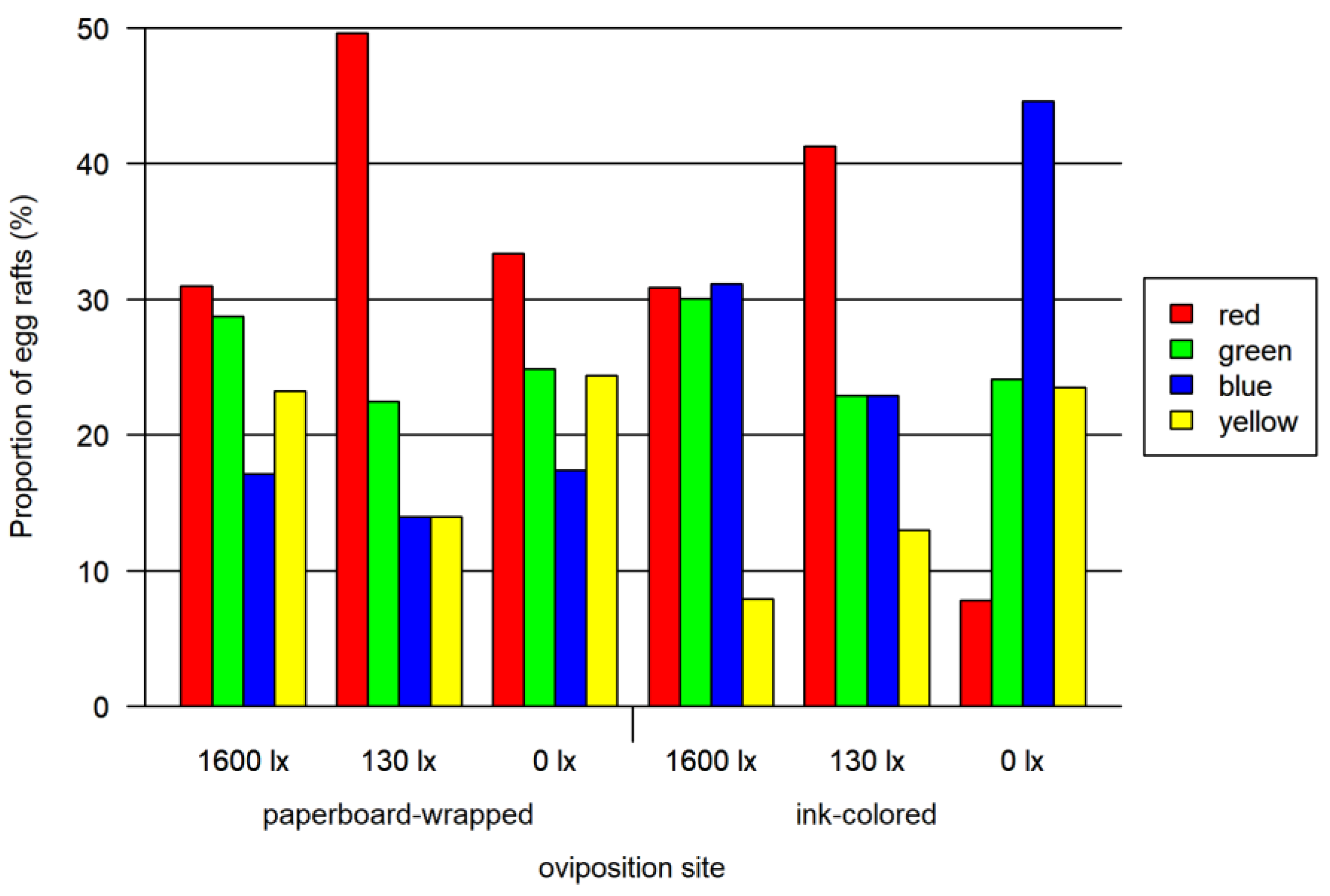
3.1. Oviposition Bioassay
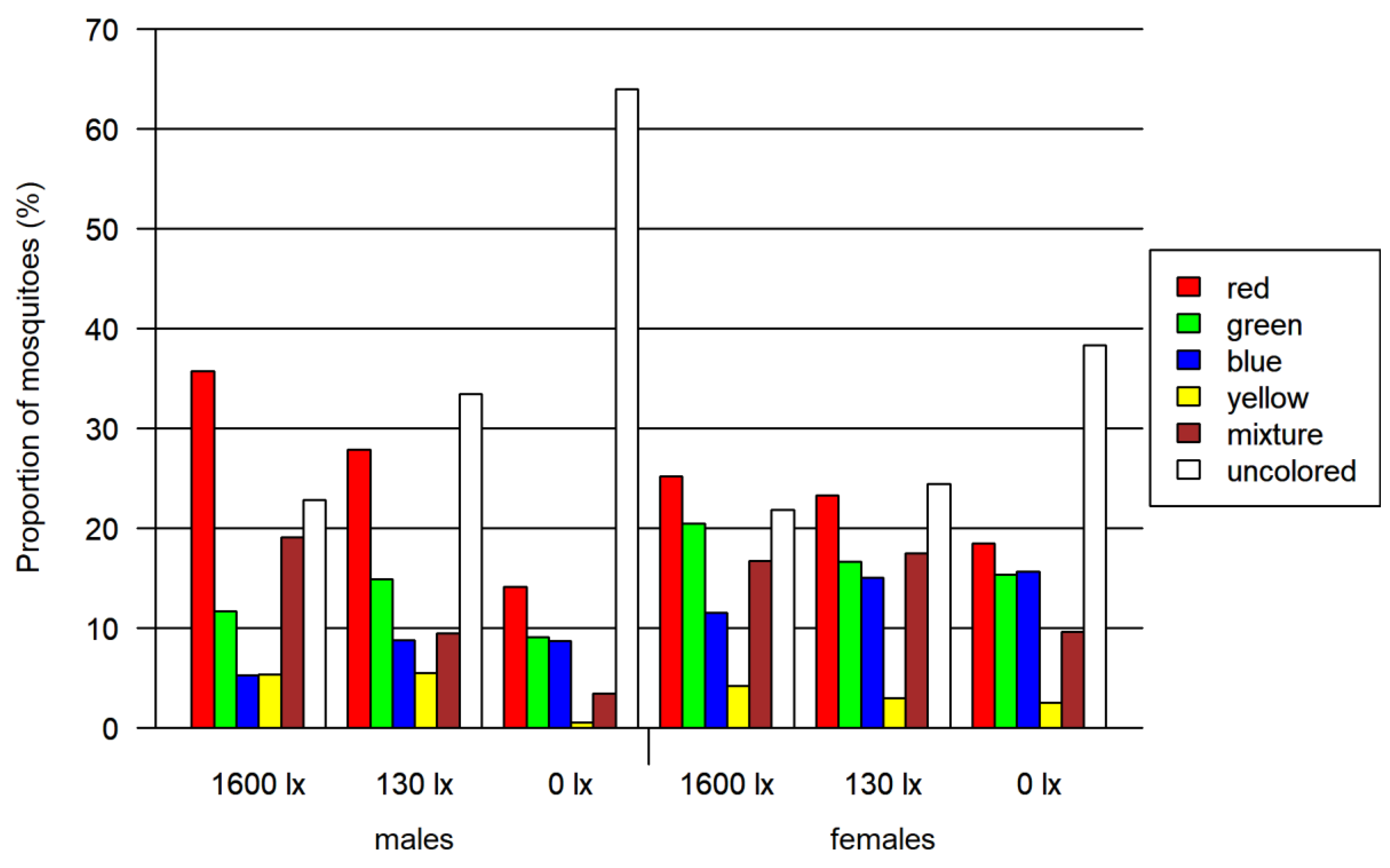
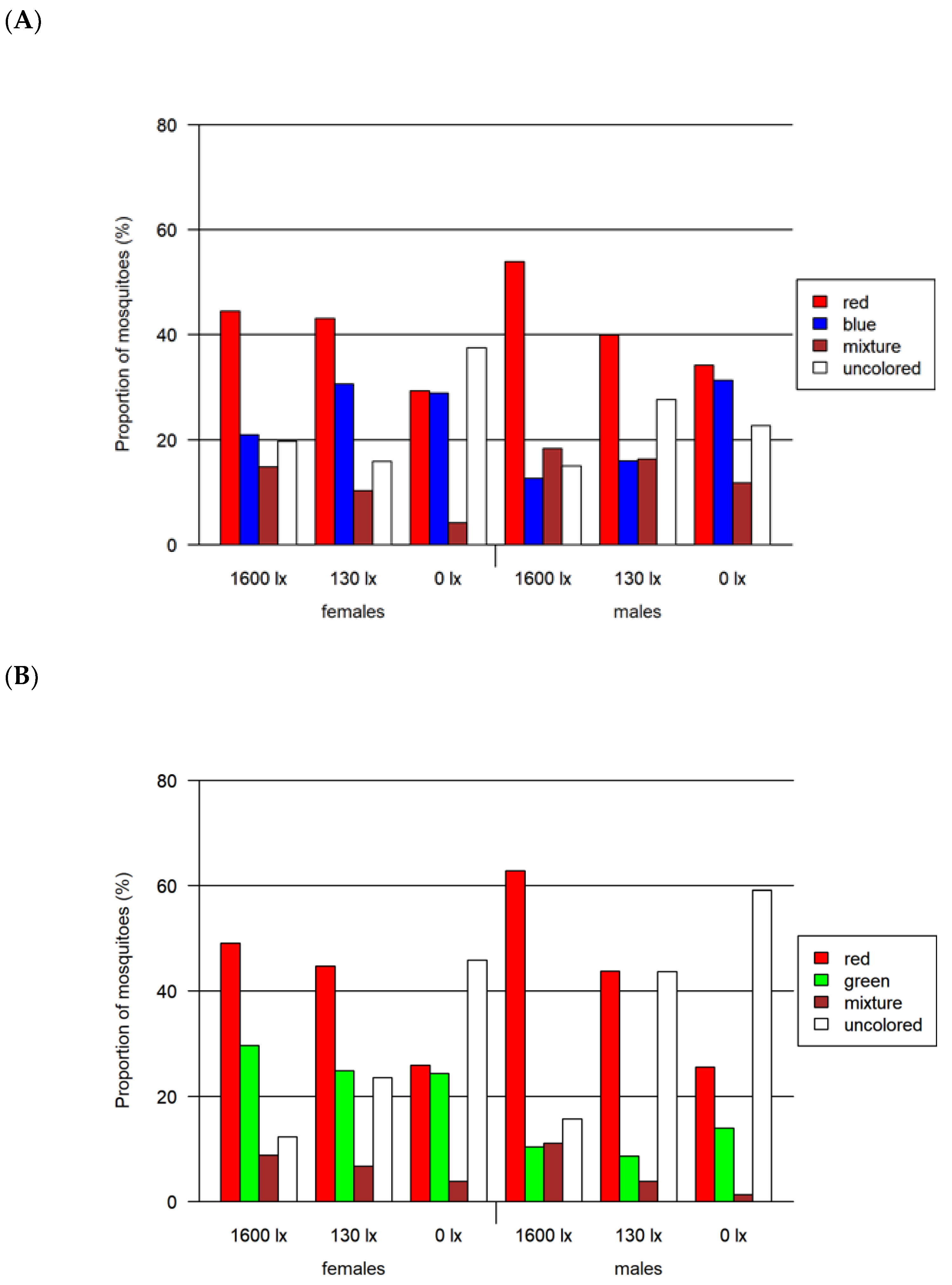
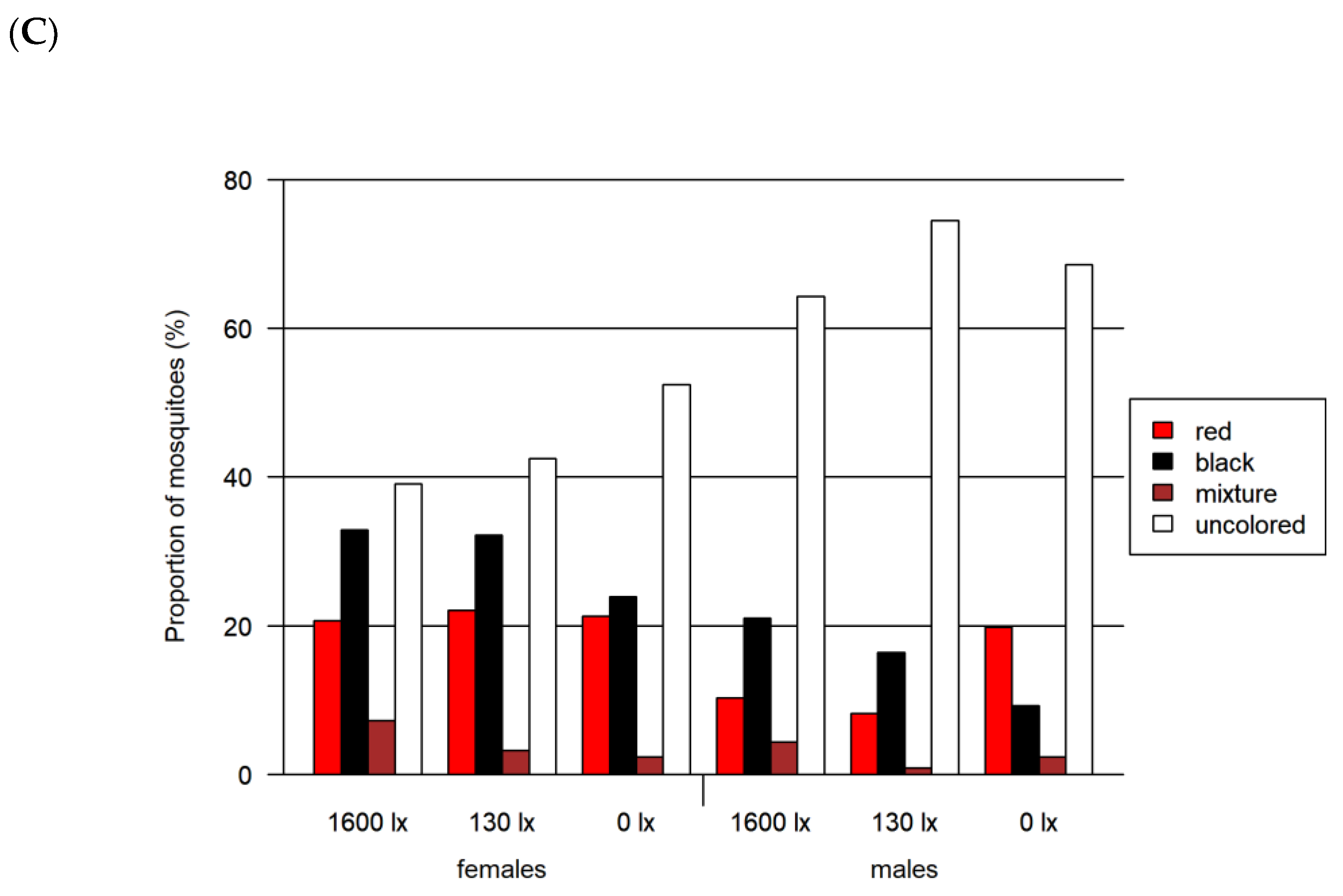
3.2. Foraging Bioassay
- (a)
- Two-choice assay red vs. blue (Figure 6A)
- (b)
- Two-choice assay red vs. green (Figure 6B)
- (c)
- Two-choice assay red vs. black (Figure 6C)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Day, J.F. Mosquito oviposition behavior and vector control. Insects 2016, 7, 65. [Google Scholar] [CrossRef] [PubMed]
- Borel, D.-T.; Elysée, N.; Abdoul, T.; Leslie, N.D.; Roland, B.; Edmond, K.; Parfait, A.-A.; Timoléon, T.; Antonio-Nkondjio, C. Oviposition Behavior of Culex quinquefasciatus and Anopheles coluzzii Females According to the Ovitrap Color and Presence of Fertilizer in Breeding Sites. Fortune J. Health Sci. 2021, 4, 207–220. [Google Scholar] [CrossRef]
- Allan, S.A.; Day, J.F.; Edman, J.D. Visual ecology of biting flies. Annu. Rev. Entomol. 1987, 32, 297–314. [Google Scholar] [CrossRef] [PubMed]
- Bentley, M.D.; Day, J.F. Chemical ecology and behavioral aspects of mosquito oviposition. Annu. Rev. Entomol. 1989, 34, 401–421. [Google Scholar] [CrossRef] [PubMed]
- Geetha, I.; Paily, K.; Padmanaban, V.; Balaraman, K. Oviposition response of the mosquito, Culex quinquefasciatus to the secondary metabolite (s) of the fungus, Trichoderma viride. Memórias Do Inst. Oswaldo Cruz 2003, 98, 223–226. [Google Scholar] [CrossRef]
- Millar, J.; Chaney, J.; Beehler, J.; Mulla, M. Interaction of the Culex quinquefasciatus egg raft pheromone with a natural chemical associated with oviposition sites. J. Am. Mosq. Control Assoc. 1994, 10, 374–379. [Google Scholar] [PubMed]
- Gubler, D.J. Studies on the comparative oviposition behavior of Aedes (Stegomyia) albopictus and Aedes (Stegomyia) polynesiensis Marks. J. Med. Entomol. 1971, 8, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Hoel, D.F.; Obenauer, P.J.; Clark, M.; Smith, R.; Hughes, T.H.; Larson, R.T.; Diclaro, J.W.; Allan, S.A. Efficacy of Ovitrap Colors and Patterns for Attracting Aedes albopictus at Suburban Field Sites in North-Central Florida1. J. Am. Mosq. Control Assoc. 2011, 27, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Muir, L.E.; Kay, B.H.; Thorne, M.J. Aedes aegypti (Diptera: Culicidae) vision: Response to stimuli from the optical environment. J. Med. Entomol. 1992, 29, 445–450. [Google Scholar] [CrossRef]
- Li, M.; Yang, T.; Kandul, N.P.; Bui, M.; Gamez, S.; Raban, R.; Bennett, J.; Lanzaro, G.C.; Schmidt, H.; Lee, Y. Development of a confinable gene drive system in the human disease vector Aedes aegypti. Elife 2020, 9, e51701. [Google Scholar] [CrossRef]
- Frank, J. Use of an artificial bromeliad to show the importance of color value in restricting colonization of bromeliads by Aedes aegypti and Culex quinquefasciatus. J. Am. Mosq. Control Assoc. 1985, 1, 28–32. [Google Scholar]
- Dhileepan, K. Physical factors and chemical cues in the oviposition behavior of arboviral vectors Culex annulirostris and Culex molestus (Diptera: Culicidae). Environ. Entomol. 1997, 26, 318–326. [Google Scholar] [CrossRef]
- Burkett, D.A.; Butler, J.F. Laboratory evaluation of colored light as an attractant for female Aedes aegypti, Aedes albopictus, Anopheles quadrimaculatus, and Culex nigripalpus. Fla. Entomol. 2005, 88, 383–390. [Google Scholar] [CrossRef]
- Kay, B.; Sutton, K.; Russell, B. A sticky entry-exit trap for sampling mosquitoes in subterranean habitats. J. Am. Mosq. Control Assoc.-Mosq. News 2000, 16, 262–265. [Google Scholar]
- Tainchum, K.; Polsomboon, S.; Grieco, J.P.; Suwonkerd, W.; Prabaripai, A.; Sungvornyothin, S.; Chareonviriyaphap, T.; Achee, N.L. Comparison of Aedes aegypti (Diptera: Culicidae) resting behavior on two fabric types under consideration for insecticide treatment in a push-pull strategy. J. Med. Entomol. 2013, 50, 59–68. [Google Scholar] [CrossRef]
- Brugman, V.A.; Hernández-Triana, L.M.; Medlock, J.M.; Fooks, A.R.; Carpenter, S.; Johnson, N. The role of Culex pipiens L.(Diptera: Culicidae) in virus transmission in Europe. Int. J. Environ. Res. Public Health 2018, 15, 389. [Google Scholar] [CrossRef]
- Darsie, R.F., Jr.; Ward, R.A. Identification and Geographical Distribution of the Mosquitoes of North America, North of Mexico; University Press of Florida: Washington DC, USA, 2005. [Google Scholar]
- Hamer, G.L.; Kitron, U.D.; Brawn, J.D.; Loss, S.R.; Ruiz, M.O.; Goldberg, T.L.; Walker, E.D. Culex pipiens (Diptera: Culicidae): A bridge vector of West Nile virus to humans. J. Med. Entomol. 2008, 45, 125–128. [Google Scholar] [CrossRef]
- Farajollahi, A.; Fonseca, D.M.; Kramer, L.D.; Kilpatrick, A.M. “Bird biting” mosquitoes and human disease: A review of the role of Culex pipiens complex mosquitoes in epidemiology. Infect. Genet. Evol. 2011, 11, 1577–1585. [Google Scholar] [CrossRef]
- Becker, N.; Jöst, A.; Weitzel, T. The Culex pipiens complex in Europe. J. Am. Mosq. Control Assoc. 2012, 28, 53–67. [Google Scholar] [CrossRef]
- Matthews, R.W.; Matthews, J.R. Insect Behavior; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Collins, L.E.; Blackwell, A. Colour cues for oviposition behaviour in Toxorhynchites moctezuma and Toxorhynchites amboinensis mosquitoes. J. Vector Ecol. J. Soc. Vector Ecol. 2000, 25, 127–135. [Google Scholar]
- Perea, N.O.; Callaghan, A. Pond dyes are Culex mosquito oviposition attractants. PeerJ 2017, 5, e3361. [Google Scholar] [CrossRef] [PubMed]
- Dieng, H.; Satho, T.; Arzemi, N.A.B.; Aliasan, N.E.; Abang, F.; Wydiamala, E.; Miake, F.; Zuharah, W.F.; Kassim, N.F.A.; Vargas, R.E.M. Exposure of a diurnal mosquito vector to floral mimics: Foraging responses, feeding patterns, and significance for sugar bait technology. Acta Trop. 2018, 185, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Peach, D.A.; Ko, E.; Blake, A.J.; Gries, G. Ultraviolet inflorescence cues enhance attractiveness of inflorescence odour to Culex pipiens mosquitoes. PLoS ONE 2019, 14, e0217484. [Google Scholar] [CrossRef]
- Jung, S.H.; Kim, D.; Jung, K.-S.; Lee, D.-K. Color Preference for Host-Seeking Activity of Aedes albopictus and Culex pipiens (Diptera: Culicidae). J. Med. Entomol. 2021, 58, 2446–2452. [Google Scholar] [CrossRef]
- Veronesi, R.; Gentile, G.; Carrieri, M.; Maccagnani, B.; Stermieri, L.; Bellini, R. Seasonal pattern of daily activity of Aedes caspius, Aedes detritus, Culex modestus, and Culex pipiens in the Po Delta of northern Italy and significance for vector-borne disease risk assessment. J. Vector Ecol. 2012, 37, 49–61. [Google Scholar] [CrossRef]
- Warrant, E.; Dacke, M. Vision and visual navigation in nocturnal insects. Annu. Rev. Entomol. 2011, 56, 239–254. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing Version 4.2.1; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Yee, T.W. The VGAM package for categorical data analysis. J. Stat. Softw. 2010, 32, 1–34. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Ben-Yakir, D.; Antignus, Y.; Offir, Y.; Shahak, Y. Colored shading nets impede insect invasion and decrease the incidences of insect-transmitted viral diseases in vegetable crops. Entomol. Exp. Et Appl. 2012, 144, 249–257. [Google Scholar] [CrossRef]
- Brett, G. On the relative attractiveness to Aedes aegypti of certain coloured cloths. Trans. R. Soc. Trop. Med. Hyg. 1938, 32, 113–124. [Google Scholar] [CrossRef]
- Haas, G. Entwicklung des Komplexauges bei Culex pipiens und Aedes aegypti. Z. Für Morphol. Und Ökologie Der Tiere 1956, 45, 198–216. [Google Scholar] [CrossRef]
- Land, M.; Gibson, G.; Horwood, J.; Zeil, J. Fundamental differences in the optical structure of the eyes of nocturnal and diurnal mosquitoes. J. Comp. Physiol. A 1999, 185, 91–103. [Google Scholar] [CrossRef]
- Muir, L.E.; Thorne, M.J.; Kay, B.H. Aedes aegypti (Diptera: Culicidae) vision: Spectral sensitivity and other perceptual parameters of the female eye. J. Med. Entomol. 1992, 29, 278–281. [Google Scholar] [CrossRef] [PubMed]
- Bowen, M.; Davis, E.; Haggart, D. A behavioural and sensory analysis of host-seeking behaviour in the diapausing mosquito Culex pipiens. J. Insect Physiol. 1988, 34, 805–813. [Google Scholar] [CrossRef]
- Anderson, J.F.; Main, A.J.; Ferrandino, F.J.; Andreadis, T.G. Nocturnal activity of mosquitoes (Diptera: Culicidae) in a West Nile virus focus in Connecticut. J. Med. Entomol. 2007, 44, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Honnen, A.-C.; Kypke, J.L.; Hölker, F.; Monaghan, M.T. Artificial light at night influences clock-gene expression, activity, and fecundity in the mosquito Culex pipiens f. molestus. Sustainability 2019, 11, 6220. [Google Scholar] [CrossRef]
- Pickel, L.; Sung, H.-K. Feeding rhythms and the circadian regulation of metabolism. Front. Nutr. 2020, 7, 39. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | Light Intensity in Lux | Egg Rafts Laid | Number of Tested Females | Average Number of Egg Rafts per Female | Mean |
|---|---|---|---|---|---|
| paperboard-wrapped beaker | 1600 | 181 | 654 | 0.28 | 0.31 |
| 130 | 258 | 669 | 0.39 | ||
| 0 | 201 | 744 | 0.27 | ||
| ink-colored water | 1600 | 642 | 1374 | 0.47 | 0.50 |
| 130 | 594 | 809 | 0.73 | ||
| 0 | 166 | 629 | 0.26 | ||
| Mean | 340.3 | 813.2 | 0.42 |
| Two-Choice Assay | Light Intensity (in Lux) | Number of Sucked (Colored) Mosquitoes (and Their Proportion) | Assay Mean | ||
|---|---|---|---|---|---|
| Total | Female | Male | |||
| Red vs. blue | 1600 (16:8 L:D) | 1325/1590 (83.4%) | 373/455 (82.0%) | 568/673 (84.4%) | 4686/6092 (76.9%) |
| 130 | 1804/2321 (77.2%) | 819/975 (84.0%) | 788/1106 (71.2%) | ||
| 0 | 1557/2181 (71.4%) | 545/872 (62.5%) | 1012/1309 (77.3%) | ||
| Red vs. green | 1600 (16:8 L:D) | 1309/1526 (85.8%) | 596/680 (87.6%) | 713/846 (84.3%) | 3216/4863 (66.1%) |
| 130 | 1171/1779 (65.8%) | 644/843 (76.4%) | 527/936 (56.3%) | ||
| 0 | 736/1558 (47.2%) | 405/748 (54.1%) | 331/810 (40.9%) | ||
| Red vs. black | 1600 (16:8 L:D) | 593/1274 (46.5%) | 333/547 (60.9%) | 260/727 (35.8%) | 1908/4606 (41.4%) |
| 130 | 699/1782 (39.2%) | 438/761 (57.6%) | 261/1021 (25.6%) | ||
| 0 | 616/1550 (39.7%) | 378/794 (47.6%) | 238/756 (31.5%) | ||
| Overall mean | 9229/14,859 (62.1%) | 4531/6675 (67.9%) | 4698/8184 (57.4%) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hellhammer, F.; Heidtmann, H.; Freise, F.; Becker, S.C. Effects of Color and Light Intensity on the Foraging and Oviposition Behavior of Culex pipiens biotype molestus Mosquitoes. Insects 2022, 13, 993. https://doi.org/10.3390/insects13110993
Hellhammer F, Heidtmann H, Freise F, Becker SC. Effects of Color and Light Intensity on the Foraging and Oviposition Behavior of Culex pipiens biotype molestus Mosquitoes. Insects. 2022; 13(11):993. https://doi.org/10.3390/insects13110993
Chicago/Turabian StyleHellhammer, Fanny, Hella Heidtmann, Fritjof Freise, and Stefanie C. Becker. 2022. "Effects of Color and Light Intensity on the Foraging and Oviposition Behavior of Culex pipiens biotype molestus Mosquitoes" Insects 13, no. 11: 993. https://doi.org/10.3390/insects13110993
APA StyleHellhammer, F., Heidtmann, H., Freise, F., & Becker, S. C. (2022). Effects of Color and Light Intensity on the Foraging and Oviposition Behavior of Culex pipiens biotype molestus Mosquitoes. Insects, 13(11), 993. https://doi.org/10.3390/insects13110993