
Transcriptomic Identification and Expression Profile Analysis of Odorant-Degrading Enzymes from the Asian Corn Borer Moth, Ostrinia furnacalis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing and Tissue Collection
2.2. RNA Extraction, cDNA Library Construction, and Transcriptome Sequencing
2.3. De Novo Assembly and Functional Annotation
2.4. Identification of Genes Encoding ODEs
2.5. Bioinformatic Analyses
2.6. Expression Profile Analysis
2.7. Data Statistics
3. Results
3.1. Unigene Assembly and Functional Annotation
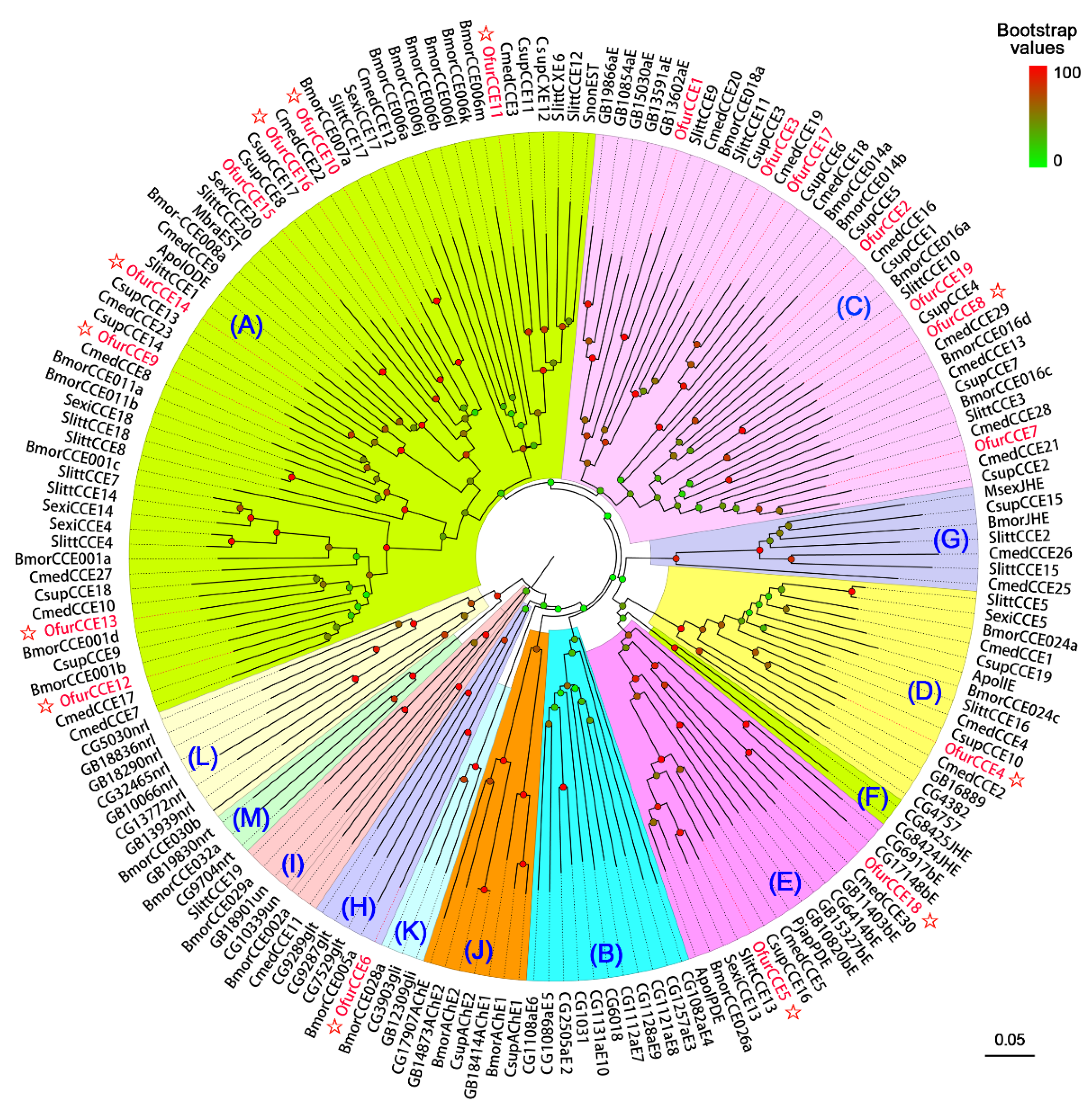
3.2. Identification of CCEs
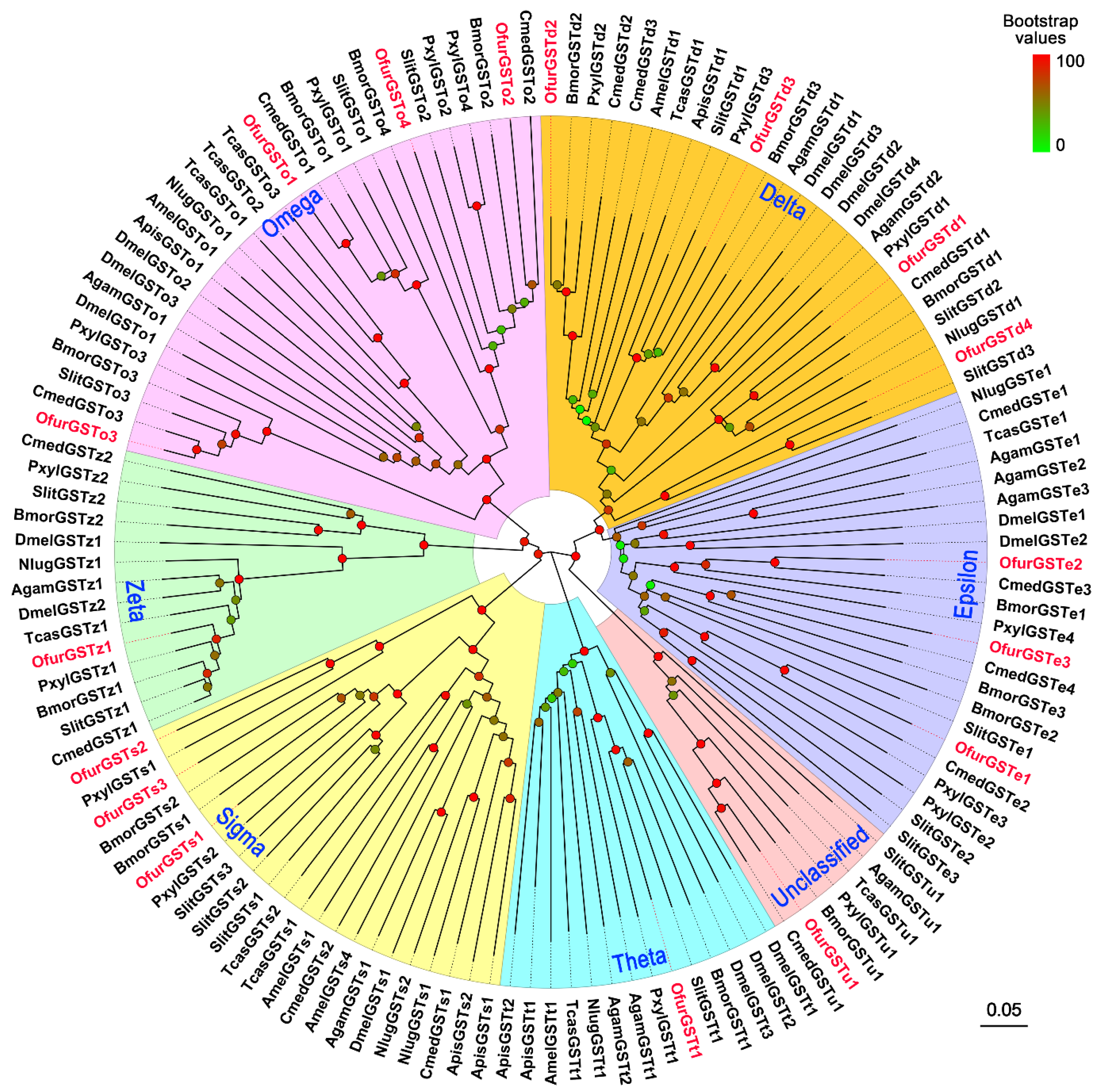
3.3. Identification of GSTs
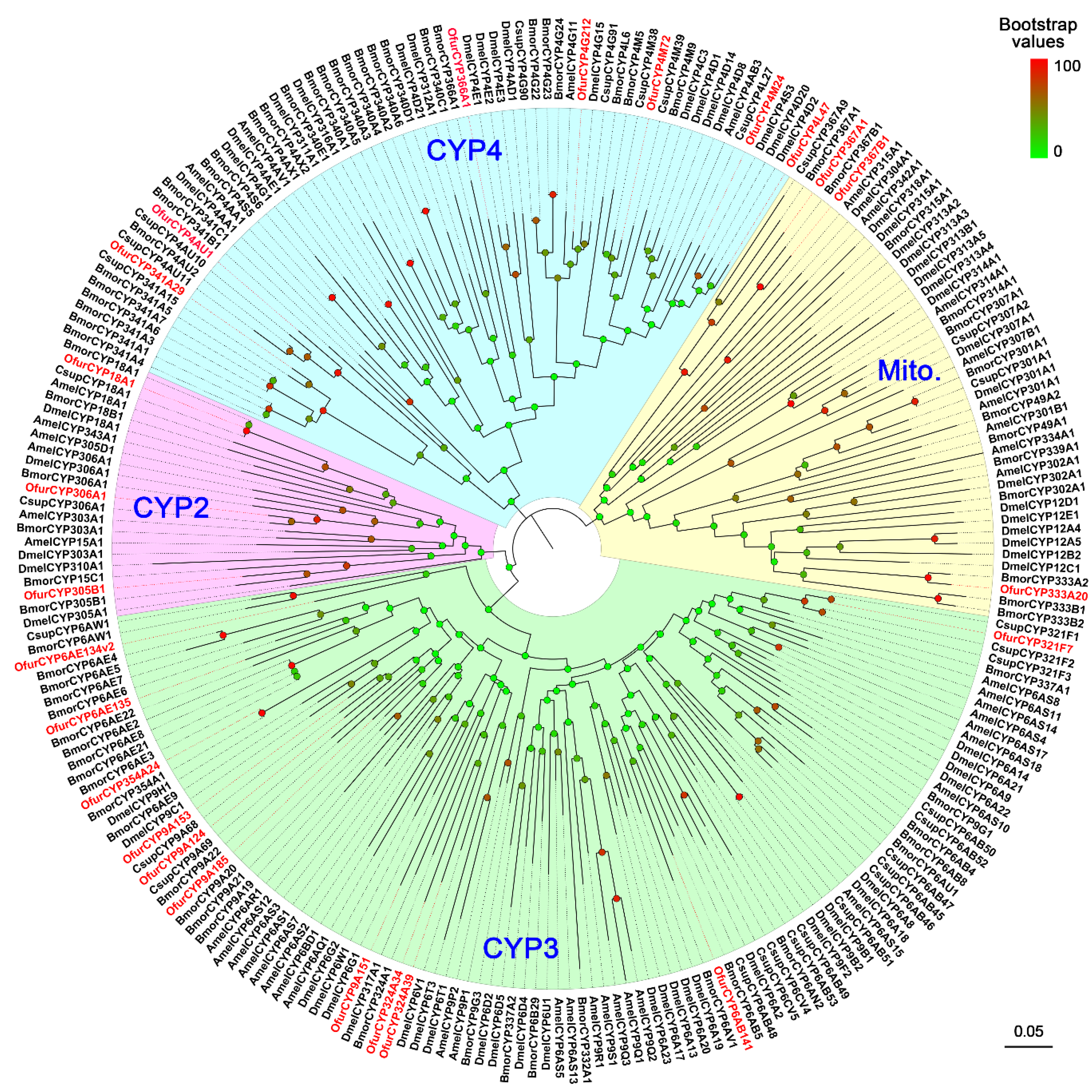
3.4. Identification of CYPs
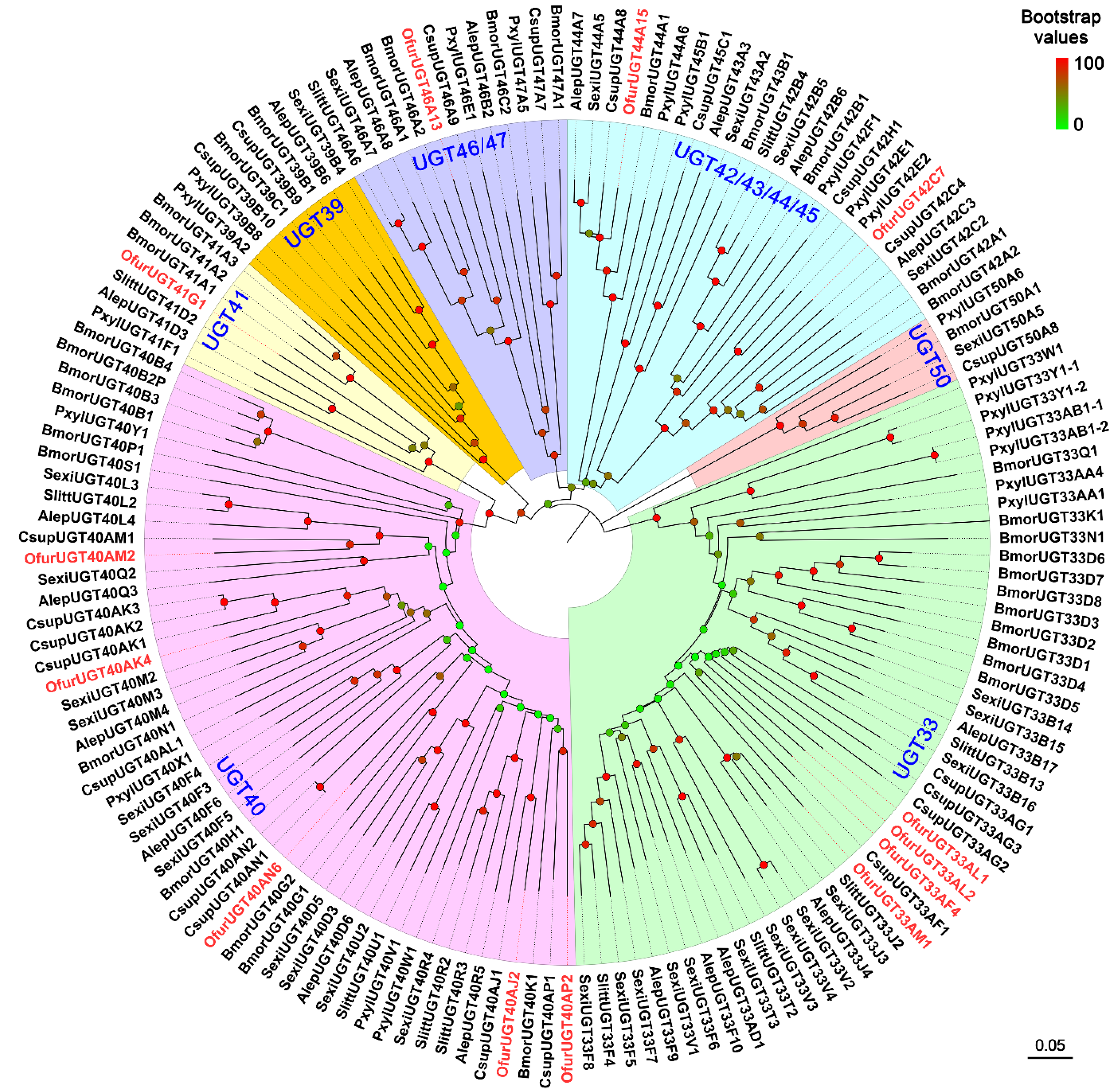
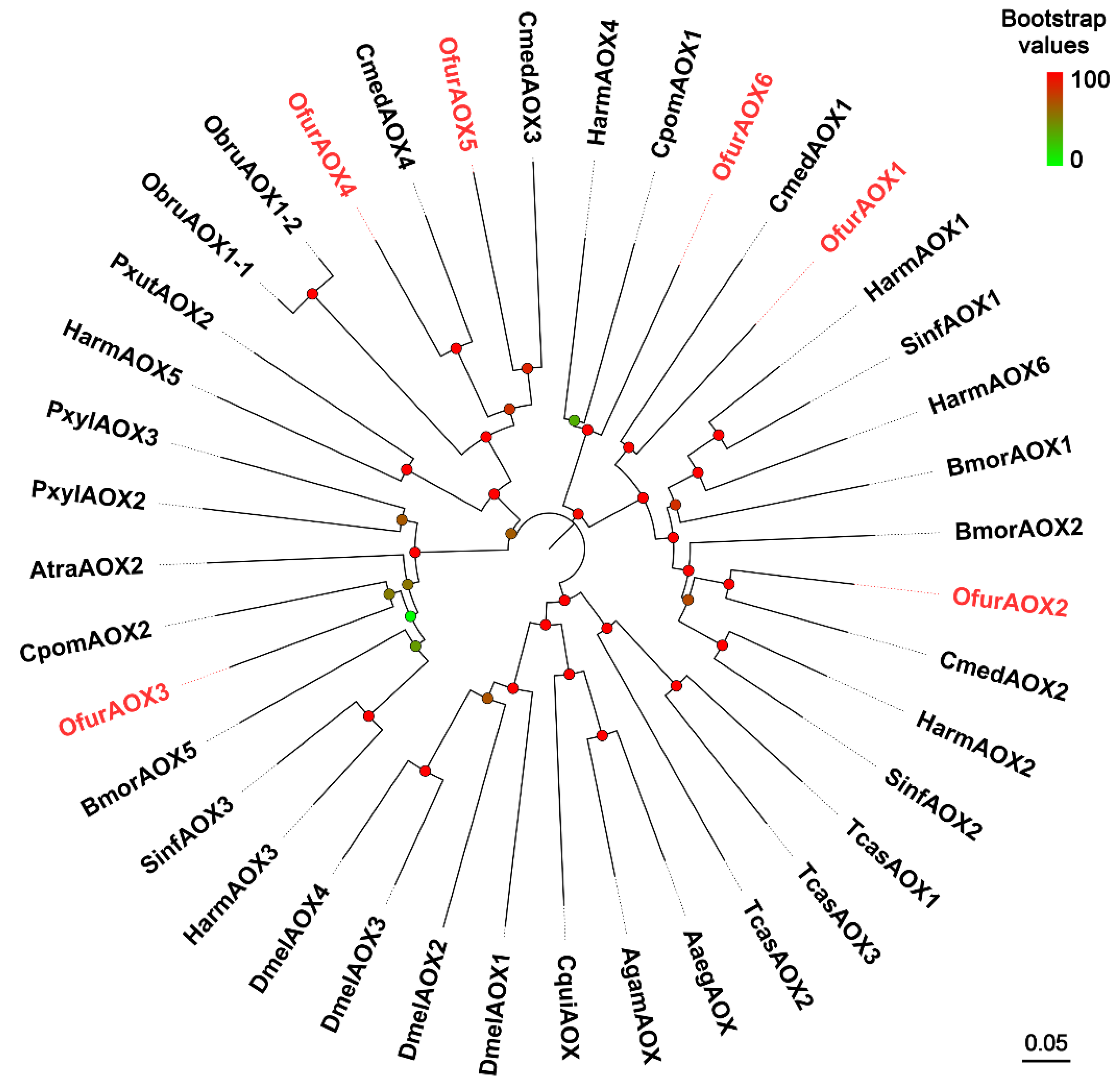
3.5. Identification of UGTs and AOXs
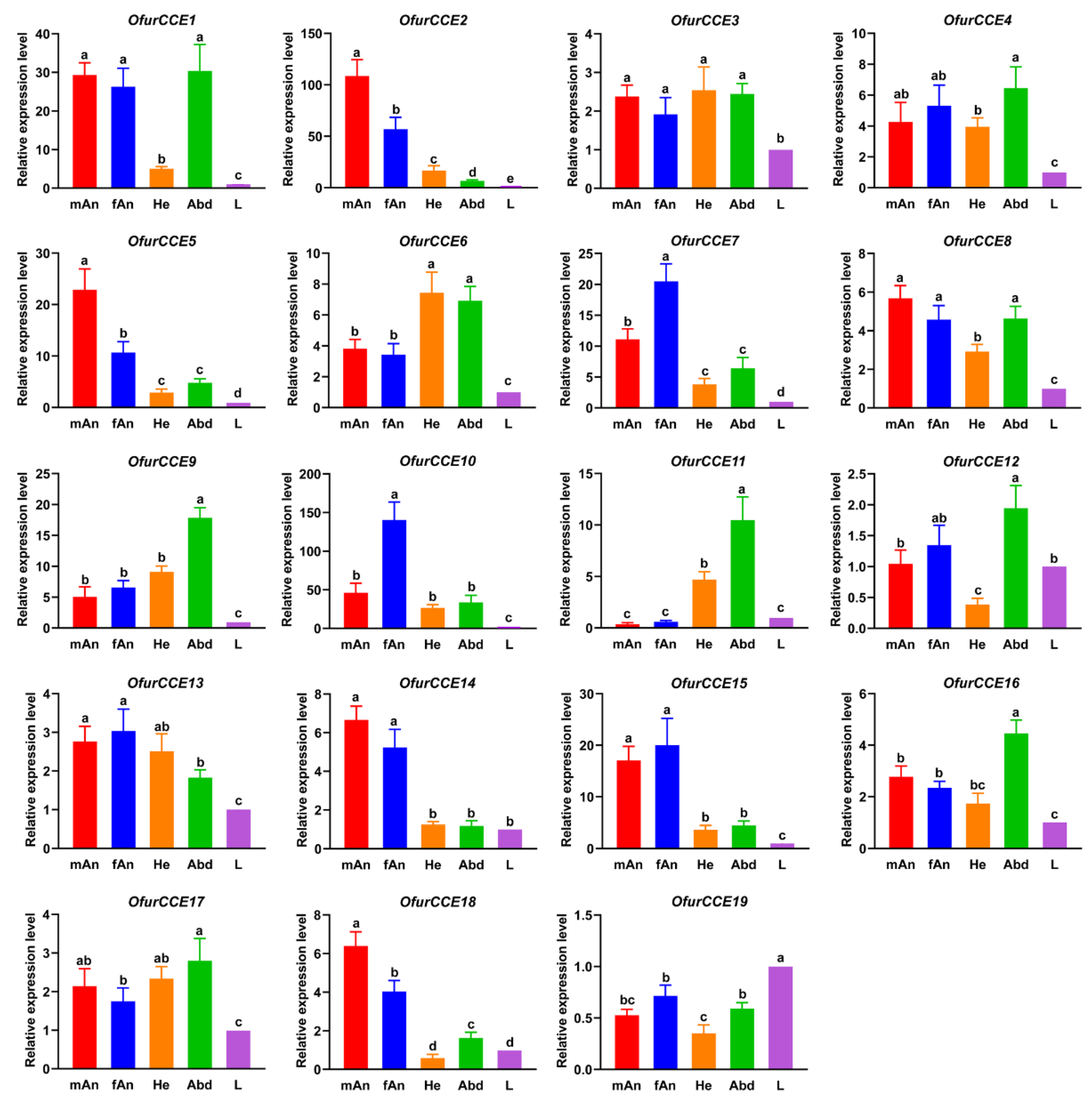
3.6. Expression Profiles of O. furnacalis CCEs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Renou, M.; Anton, S. Insect olfactory communication in a complex and changing world. Curr. Opin. Insect Sci. 2020, 42, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Leal, W.S. Odorant reception in insects: Roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef] [PubMed]
- Robertson, H.M. Molecular evolution of the major arthropod chemoreceptor gene families. Annu. Rev. Entomol. 2019, 64, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Steiner, C.; Chertemps, T.; Maïbèche, M. Diversity of biotransformation enzymes in insect antennae: Possible roles in odorant inactivation and xenobiotic processing. In Olfactory Concepts of Insect Control—Alternative to Insecticides; Picimbon, J.-F., Ed.; Springer International Publishing: Cham, Switzerland, 2019; Volume 2, pp. 115–145. [Google Scholar]
- Godoy, R.; Machuca, J.; Venthur, H.; Quiroz, A.; Mutis, A. An overview of antennal esterases in Lepidoptera. Front. Physiol. 2021, 12, 643281. [Google Scholar] [CrossRef]
- Vogt, R.G.; Riddiford, L.M. Pheromone binding and inactivation by moth antennae. Nature 1981, 293, 161–163. [Google Scholar] [CrossRef]
- Ishida, Y.; Leal, W.S. Rapid inactivation of a moth pheromone. Proc. Natl. Acad. Sci. USA 2005, 102, 14075–14079. [Google Scholar] [CrossRef]
- Chertemps, T.; Younus, F.; Steiner, C.; Durand, N.; Coppin, C.W.; Pandey, G.; Oakeshott, J.G.; Maïbèche, M. An antennal carboxylesterase from Drosophila melanogaster, esterase 6, is a candidate odorant-degrading enzyme toward food odorants. Front. Physiol. 2015, 6, 315. [Google Scholar] [CrossRef]
- Younus, F.; Chertemps, T.; Pearce, S.L.; Pandey, G.; Bozzolan, F.; Coppin, C.W.; Russell, R.J.; Maïbèche-Coisne, M.; Oakeshott, J.G. Identification of candidate odorant degrading gene/enzyme systems in the antennal transcriptome of Drosophila melanogaster. Insect Biochem. Mol. Biol. 2014, 53, 30–43. [Google Scholar] [CrossRef]
- Ishida, Y.; Leal, W.S. Chiral discrimination of the Japanese beetle sex pheromone and a behavioral antagonist by a pheromone-degrading enzyme. Proc. Natl. Acad. Sci. USA 2008, 105, 9076–9080. [Google Scholar] [CrossRef]
- Durand, N.; Carot-Sans, G.; Chertemps, T.; Bozzolan, F.; Party, V.; Renou, M.; Debernard, S.; Rosell, G.; Maïbèche-Coisne, M. Characterization of an antennal carboxylesterase from the pest moth Spodoptera littoralis degrading a host plant odorant. PLoS ONE 2010, 5, e15026. [Google Scholar] [CrossRef]
- Durand, N.; Carot-Sans, G.; Bozzolan, F.; Rosell, G.; Siaussat, D.; Debernard, S.; Chertemps, T.; Maïbèche-Coisne, M. Degradation of pheromone and plant volatile components by a same odorant-degrading enzyme in the cotton leafworm, Spodoptera littoralis. PLoS ONE 2011, 6, e29147. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Li, Z.-Q.; Liu, C.-C.; Liu, S.-J.; Dong, S.-L. Two esterases from the genus Spodoptera degrade sex pheromones and plant volatiles. Genome 2014, 57, 201–208. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Zhang, J.; Li, Z.-Q.; Zhang, Y.-N.; Yang, K.; Dong, S.-L. Functional characterization of an antennal esterase from the noctuid moth, Spodoptera exigua. Arch. Insect Biochem. 2014, 86, 85–99. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Zhang, Y.N.; Li, Z.Q.; Yang, K.; Zhu, J.Y.; Liu, S.J.; Dong, S.L. An antennae-enriched carboxylesterase from Spodoptera exigua displays degradation activity in both plant volatiles and female sex pheromones. Insect Mol. Biol. 2014, 23, 475–486. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Zhang, Y.-N.; Yang, K.; Li, Z.-Q.; Dong, S.-L. An antenna-biased carboxylesterase is specifically active to plant volatiles in Spodoptera exigua. Pestic. Biochem. Physiol. 2015, 123, 93–100. [Google Scholar] [CrossRef]
- He, P.; Mang, D.-Z.; Wang, H.; Wang, M.-M.; Ma, Y.-F.; Wang, J.; Chen, G.-L.; Zhang, F.; He, M. Molecular characterization and functional analysis of a novel candidate of cuticle carboxylesterase in Spodoptera exigua degradating sex pheromones and plant volatile esters. Pestic. Biochem. Physiol. 2020, 163, 227–234. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, Y.-X.; Wang, W.-L.; Zhang, B.-X.; Li, S.-G. Identification and characterisation of seventeen glutathione S-transferase genes from the cabbage white butterfly Pieris rapae. Pestic. Biochem. Physiol. 2017, 143, 102–110. [Google Scholar] [CrossRef]
- Rogers, M.E.; Jani, M.K.; Vogt, R.G. An olfactory-specific glutathione-S-transferase in the sphinx moth Manduca sexta. J. Exp. Biol. 1999, 202, 1625–1637. [Google Scholar] [CrossRef]
- Tan, X.; Hu, X.-M.; Zhong, X.-W.; Chen, Q.-M.; Xia, Q.-Y.; Zhao, P. Antenna-specific glutathione S-transferase in male silkmoth Bombyx mori. Int. J. Mol. Sci. 2014, 15, 7429–7443. [Google Scholar] [CrossRef]
- Li, G.-W.; Chen, X.-L.; Xu, X.-L.; Wu, J.-X. Degradation of sex pheromone and plant volatile components by an antennal glutathione S-transferase in the oriental fruit moth, Grapholita molesta Busck (Lepidoptera: Tortricidae). Arch. Insect Biochem. 2018, 99, e21512. [Google Scholar] [CrossRef]
- Feyereisen, R. Insect CYP genes and P450 enzymes. In Insect Molecular Biology and Biochemistry; Gilbert, L.I., Ed.; Elsevier: London, UK, 2012; pp. 236–316. [Google Scholar]
- Wojtasek, H.; Leal, W.S. Degradation of an alkaloid pheromone from the pale-brown chafer, Phyllopertha diversa (Coleoptera: Scarabaeidae), by an insect olfactory cytochrome P450. FEBS Lett. 1999, 458, 333–336. [Google Scholar] [CrossRef]
- Maïbèche-Coisne, M.; Nikonov, A.A.; Ishida, Y.; Jacquin-Joly, E.; Leal, W.S. Pheromone anosmia in a scarab beetle induced by in vivo inhibition of a pheromone-degrading enzyme. Proc. Natl. Acad. Sci. USA 2004, 101, 11459–11464. [Google Scholar] [CrossRef] [PubMed]
- Calla, B.; MacLean, M.; Liao, L.-H.; Dhanjal, I.; Tittiger, C.; Blomquist, G.J.; Berenbaum, M.R. Functional characterization of CYP4G11—A highly conserved enzyme in the western honey bee Apis mellifera. Insect Mol. Biol. 2018, 27, 661–674. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Lin, X.; Zheng, K.; Qian, K.; Chang, Y.; Du, Y. Transcriptome and expression profiling analysis link patterns of gene expression to antennal responses in Spodoptera litura. BMC Genom. 2015, 16, 269. [Google Scholar] [CrossRef]
- Keeling, C.I.; Henderson, H.; Li, M.; Dullat, H.K.; Ohnishi, T.; Bohlmann, J. CYP345E2, an antenna-specific cytochrome P450 from the mountain pine beetle, Dendroctonus ponderosae Hopkins, catalyses the oxidation of pine host monoterpene volatiles. Insect Biochem. Mol. Biol. 2013, 43, 1142–1151. [Google Scholar] [CrossRef]
- Ahn, S.-J.; Vogel, H.; Heckel, D.G. Comparative analysis of the UDP-glycosyltransferase multigene family in insects. Insect Biochem. Mol. Biol. 2012, 42, 133–147. [Google Scholar] [CrossRef]
- Fraichard, S.; Legendre, A.; Lucas, P.; Chauvel, I.; Faure, P.; Neiers, F.; Artur, Y.; Briand, L.; Ferveur, J.-F.; Heydel, J.-M. Modulation of sex pheromone discrimination by a UDP-glycosyltransferase in Drosophila melanogaster. Genes 2020, 11, 237. [Google Scholar] [CrossRef]
- He, P.; Zhang, Y.-F.; Hong, D.-Y.; Wang, J.; Wang, X.-L.; Zuo, L.-H.; Tang, X.-F.; Xu, W.-M.; He, M. A reference gene set for sex pheromone biosynthesis and degradation genes from the diamondback moth, Plutella xylostella, based on genome and transcriptome digital gene expression analyses. BMC Genom. 2017, 18, 219. [Google Scholar] [CrossRef]
- Zhang, Y.-N.; Ma, J.-F.; Xu, L.; Dong, Z.-P.; Xu, J.-W.; Li, M.-Y.; Zhu, X.-Y. Identification and expression patterns of UDP-glycosyltransferase (UGT) genes from insect pest Athetis lepigone (Lepidoptera: Noctuidae). J. Asia Pac. Entomol 2017, 20, 253–259. [Google Scholar] [CrossRef]
- Rybczynski, R.; Vogt, R.G.; Lerner, M.R. Antennal-specific pheromone-degrading aldehyde oxidases from the moths Antheraea polyphemus and Bombyx mori. J. Biol. Chem. 1990, 265, 19712–19715. [Google Scholar] [CrossRef]
- Choo, Y.-M.; Pelletier, J.; Atungulu, E.; Leal, W.S. Identification and characterization of an antennae-specific aldehyde oxidase from the navel orangeworm. PLoS ONE 2013, 8, e67794. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Liao, Y. Identification and characterization of aldehyde oxidases (AOXs) in the cotton bollworm. Sci. Nat. 2017, 104, 94. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-X.; Wang, W.-L.; Li, M.-Y.; Li, S.-G.; Liu, S. Identification of putative carboxylesterase and aldehyde oxidase genes from the antennae of the rice leaffolder, Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). J. Asia Pac. Entomol 2017, 20, 907–913. [Google Scholar] [CrossRef]
- Guo, L.; Li, G.Q. Olfactory perception of oviposition-deterring fatty acids and their methyl esters by the Asian corn borer, Ostrinia furnacalis. J. Insect Sci. 2009, 9, 67. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Jiang, X.-C.; Cao, S.; Yang, B.; Wang, G.-R. Functional studies of sex pheromone receptors in Asian corn borer Ostrinia furnacalis. Front. Physiol. 2018, 9, 591. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Coates, B.S.; Ge, X.; Bai, S.; He, K.; Wang, Z. Male- and female-biased gene expression of olfactory-related genes in the antennae of Asian corn borer, Ostrinia furnacalis (Guenée) (Lepidoptera: Crambidae). PLoS ONE 2015, 10, e0128550. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Ozaki, K.; Ishikawa, Y.; Matsuo, T. Identification of candidate odorant receptors in Asian corn borer Ostrinia furnacalis. PLoS ONE 2015, 10, e0121261. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Liu, L.; Su, X.; Feng, J. Identification of biotransformation enzymes in the antennae of codling moth Cydia pomonella. Gene 2016, 580, 73–79. [Google Scholar] [CrossRef]
- Liu, S.; Gong, Z.-J.; Rao, X.-J.; Li, M.-Y.; Li, S.-G. Identification of putative carboxylesterase and glutathione S-transferase genes from the antennae of the Chilo suppressalis (Lepidoptera: Pyralidae). J. Insect Sci. 2015, 15, 103. [Google Scholar] [CrossRef]
- Zhang, Y.-N.; Li, Z.-Q.; Zhu, X.-Y.; Qian, J.-L.; Dong, Z.-P.; Xu, L.; He, P. Identification and tissue distribution of carboxylesterase (CXE) genes in Athetis lepigone (Lepidoptera: Noctuidae) by RNA-seq. J. Asia Pac. Entomol. 2017, 20, 1150–1155. [Google Scholar] [CrossRef]
- Durand, N.; Carot-Sans, G.; Chertemps, T.; Montagné, N.; Jacquin-Joly, E.; Debernard, S.; Maïbèche-Coisne, M. A diversity of putative carboxylesterases are expressed in the antennae of the noctuid moth Spodoptera littoralis. Insect Mol. Biol. 2010, 19, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Pottier, M.A.; Bozzolan, F.; Chertemps, T.; Jacquin-Joly, E.; Lalouette, L.; Siaussat, D.; Maïbèche-Coisne, M. Cytochrome P450s and cytochrome P450 reductase in the olfactory organ of the cotton leafworm Spodoptera littoralis. Insect Mol. Biol. 2012, 21, 568–580. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, A.C.; Itoh, M.; Okuda, S.; Moriya, Y.; Kanehisa, M. KAAS: An automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 2007, 35, W182–W185. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Dong, B.; Liu, X.-Y.; Li, B.; Li, M.-Y.; Li, S.-G.; Liu, S. A heat shock protein protects against oxidative stress induced by lambda-cyhalothrin in the green peach aphid Myzus persicae. Pestic. Biochem. Physiol. 2022, 181, 104995. [Google Scholar] [CrossRef]
- Wu, W.; Hao, E.; Guo, B.; Yang, H.; Zhou, J.; Ma, M.; Lu, P.; Qiao, H. Comparison of chemosensory receptor genes in the antennae transcriptomes of Sirex noctilio and Sirex nitobei (Hymenoptera: Siricidae). Forests 2022, 13, 1495. [Google Scholar] [CrossRef]
- Rancurel, C.; van Tran, T.; Elie, C.; Hilliou, F. SATQPCR: Website for statistical analysis of real-time quantitative PCR data. Mol. Cell Probe. 2019, 46, 101418. [Google Scholar] [CrossRef]
- Tang, Q.-Y.; Zhang, C.-X. Data processing system (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research. Insect Sci. 2013, 20, 254–260. [Google Scholar] [CrossRef]
- Wei, H.; Tan, S.; Li, Z.; Li, J.; Moural, T.W.; Zhu, F.; Liu, X. Odorant degrading carboxylesterases modulate foraging and mating behaviors of Grapholita molesta. Chemosphere 2021, 270, 128647. [Google Scholar] [CrossRef]
- Sun, L.; Wang, Q.; Wang, Q.; Zhang, Y.; Tang, M.; Guo, H.; Fu, J.; Xiao, Q.; Zhang, Y.; Zhang, Y. Identification and expression patterns of putative diversified carboxylesterases in the tea geometrid Ectropis obliqua Prout. Front. Physiol. 2017, 8, 1085. [Google Scholar] [CrossRef]
- Ishida, Y.; Leal, W.S. Cloning of putative odorant-degrading enzyme and integumental esterase cDNAs from the wild silkmoth, Antheraea polyphemus. Insect Biochem. Mol. Biol. 2002, 32, 1775–1780. [Google Scholar] [CrossRef]
- Maïbèche-Coisne, M.; Merlin, C.; François, M.-C.; Queguiner, I.; Porcheron, P.; Jacquin-Joly, E. Putative odorant-degrading esterase cDNA from the moth Mamestra brassicae: Cloning and expression patterns in male and female antennae. Chem. Senses 2004, 29, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Durand, N.; Pottier, M.-A.; Siaussat, D.; Bozzolan, F.; Maïbèche, M.; Chertemps, T. Glutathione-S-transferases in the olfactory organ of the Noctuid moth Spodoptera littoralis, diversity and conservation of chemosensory clades. Front. Physiol. 2018, 9, 1283. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, W.; Tao, J.; Zong, S. Antennal transcriptome analyses and olfactory protein identification in an important wood-boring moth pest, Streltzoviella insularis (Lepidoptera: Cossidae). Sci. Rep. 2019, 9, 17951. [Google Scholar] [CrossRef]
- Huang, X.; Fan, D.; Liu, L.; Feng, J. Identification and characterization of glutathione S-transferase genes in the antennae of codling moth (Lepidoptera: Tortricidae). Ann. Entomol. Soc. Am. 2017, 110, 409–416. [Google Scholar] [CrossRef]
- Leal, W.S.; Ishida, Y.; Pelletier, J.; Xu, W.; Rayo, J.; Xu, X.Z.; Ames, J.B. Olfactory proteins mediating chemical communication in the navel orangeworm moth, Amyelois transitella. PLoS ONE 2009, 4, e7235. [Google Scholar] [CrossRef]
- Glaser, N.; Gallot, A.; Legeai, F.; Harry, M.; Kaiser, L.; Le Ru, B.; Calatayud, P.-A.; Jacquin-Joly, E. Differential expression of the chemosensory transcriptome in two populations of the stemborer Sesamia nonagrioides. Insect Biochem. Mol. Biol. 2015, 65, 28–34. [Google Scholar] [CrossRef]
- Corcoran, J.A.; Jordan, M.D.; Thrimawithana, A.H.; Crowhurst, R.N.; Newcomb, R.D. The peripheral olfactory repertoire of the lightbrown apple moth, Epiphyas postvittana. PLoS ONE 2015, 10, e0128596. [Google Scholar] [CrossRef]
- Wu, H.; Liu, Y.; Shi, X.; Zhang, X.; Ye, C.; Zhu, K.Y.; Zhu, F.; Zhang, J.; Ma, E. Transcriptome analysis of antennal cytochrome P450s and their transcriptional responses to plant and locust volatiles in Locusta migratoria. Int. J. Biol. Macromol. 2020, 149, 741–753. [Google Scholar] [CrossRef]
- Wang, L.; Dankert, H.; Perona, P.; Anderson, D.J. A common genetic target for environmental and heritable influences on aggressiveness in Drosophila. Proc. Natl. Acad. Sci. USA 2008, 105, 5657–5663. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Zheng, K.; Li, C.; Guo, Q.; Du, Y. A cytochrome P450 gene plays a role in the recognition of sex pheromones in the tobacco cutworm, Spodoptera litura. Insect Mol. Biol. 2017, 26, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Xuan, N.; Guo, X.; Xie, H.-Y.; Lou, Q.-N.; Lu, X.-B.; Liu, G.-X.; Picimbon, J.-F. Increased expression of CSP and CYP genes in adult silkworm females exposed to avermectins. Insect Sci. 2015, 22, 203–219. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Xuan, N.; Liu, G.; Xie, H.; Lou, Q.; Arnaud, P.; Offmann, B.; Picimbon, J.-F. An expanded survey of the moth PBP/GOBP clade in Bombyx mori: New insight into expression and functional roles. Front. Physiol. 2021, 12, 712593. [Google Scholar] [CrossRef]
- Bozzolan, F.; Siaussat, D.; Maria, A.; Durand, N.; Pottier, M.A.; Chertemps, T.; Maïbèche-Coisne, M. Antennal uridine diphosphate (UDP)-glycosyltransferases in a pest insect: Diversity and putative function in odorant and xenobiotics clearance. Insect Mol. Biol. 2014, 23, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, J.; Bozzolan, F.; Solvar, M.; François, M.-C.; Jacquin-Joly, E.; Maïbèche-Coisne, M. Identification of candidate aldehyde oxidases from the silkworm Bombyx mori potentially involved in antennal pheromone degradation. Gene 2007, 404, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.-M.; He, M.; Wang, H.; Ma, Y.-F.; Dewer, Y.; Zhang, F.; He, P. A candidate aldehyde oxidase in the antennae of the diamondback moth, Plutella xylostella (L.), is potentially involved in the degradation of pheromones, plant-derived volatiles and the detoxification of xenobiotics. Pestic. Biochem. Physiol. 2021, 171, 104726. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Shen, Y.; Jiang, X.; Liu, S. Transcriptomic Identification and Expression Profile Analysis of Odorant-Degrading Enzymes from the Asian Corn Borer Moth, Ostrinia furnacalis. Insects 2022, 13, 1027. https://doi.org/10.3390/insects13111027
Zhang L, Shen Y, Jiang X, Liu S. Transcriptomic Identification and Expression Profile Analysis of Odorant-Degrading Enzymes from the Asian Corn Borer Moth, Ostrinia furnacalis. Insects. 2022; 13(11):1027. https://doi.org/10.3390/insects13111027
Chicago/Turabian StyleZhang, Liya, Yidan Shen, Xingchuan Jiang, and Su Liu. 2022. "Transcriptomic Identification and Expression Profile Analysis of Odorant-Degrading Enzymes from the Asian Corn Borer Moth, Ostrinia furnacalis" Insects 13, no. 11: 1027. https://doi.org/10.3390/insects13111027
APA StyleZhang, L., Shen, Y., Jiang, X., & Liu, S. (2022). Transcriptomic Identification and Expression Profile Analysis of Odorant-Degrading Enzymes from the Asian Corn Borer Moth, Ostrinia furnacalis. Insects, 13(11), 1027. https://doi.org/10.3390/insects13111027