
Effects of Nano-Graphene Oxide on the Growth and Reproductive Dynamics of Spodoptera frugiperda Based on an Age-Stage, Two-Sex Life Table


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Preparation of GO Diets
2.3. Characterization and Stability of GO
2.4. Developmental Times
2.5. Age-Stage, Two-Sex Life Table
2.6. Gene Expression Studies
2.7. Data Analysis
3. Results
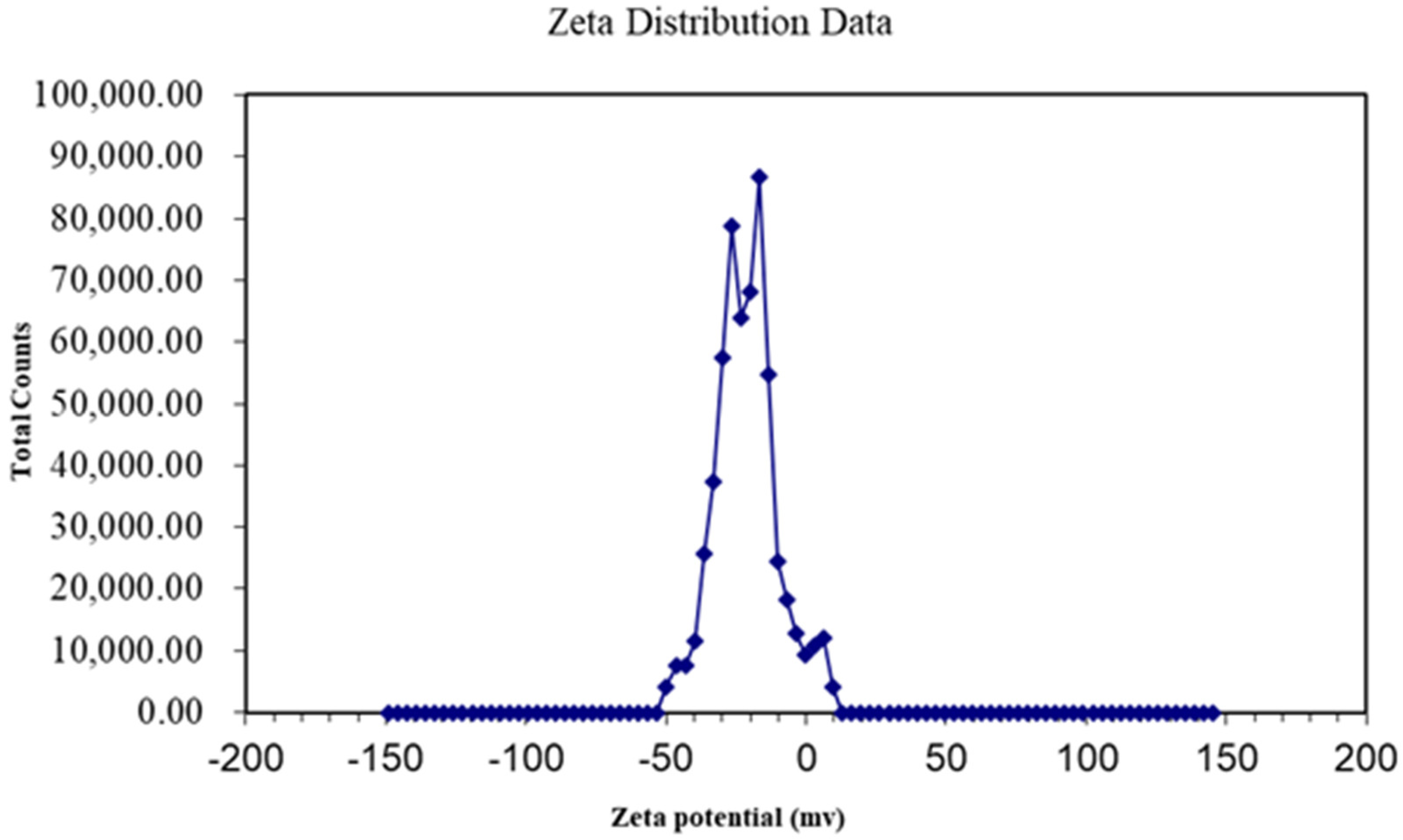
3.1. Characteristics of GO
3.2. Developmental Duration of FAW Life Stages
3.3. FAW Pre-Oviposition, Egg Production, Pupal Weight, Female-to-Male Ratio and Lifespan
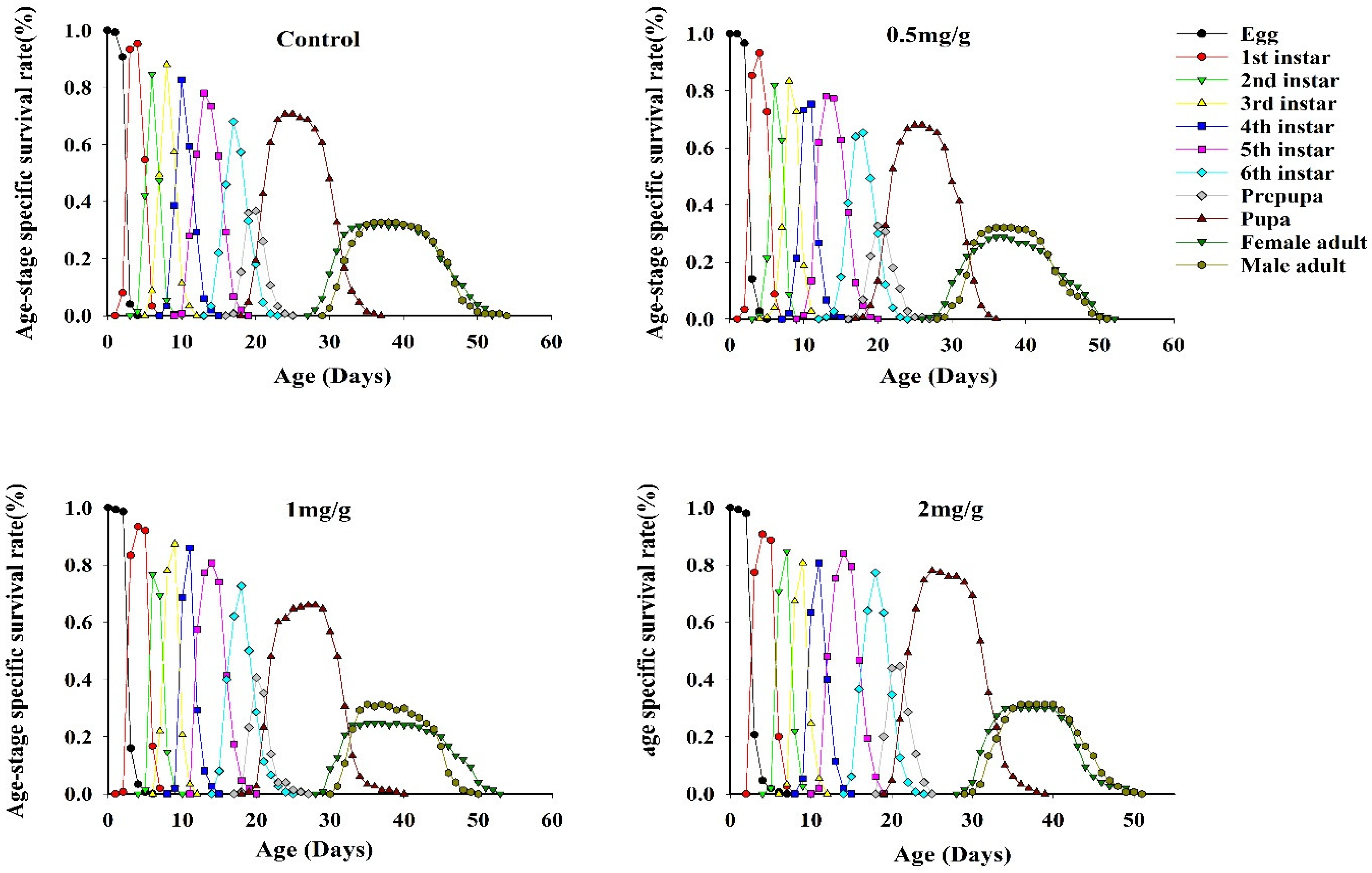
3.4. Survival Rates of FAWs
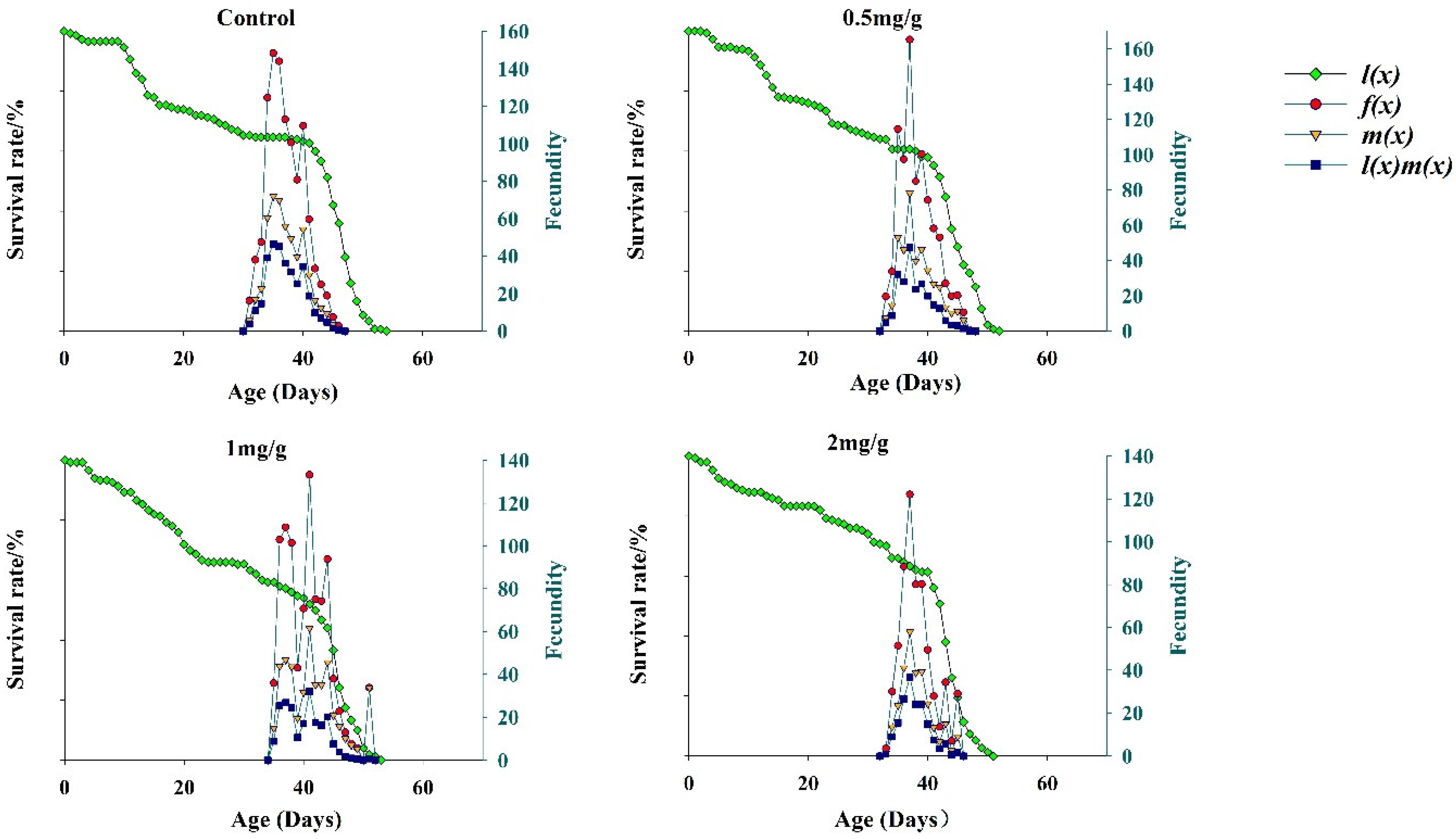
3.5. Fecundity of FAWs
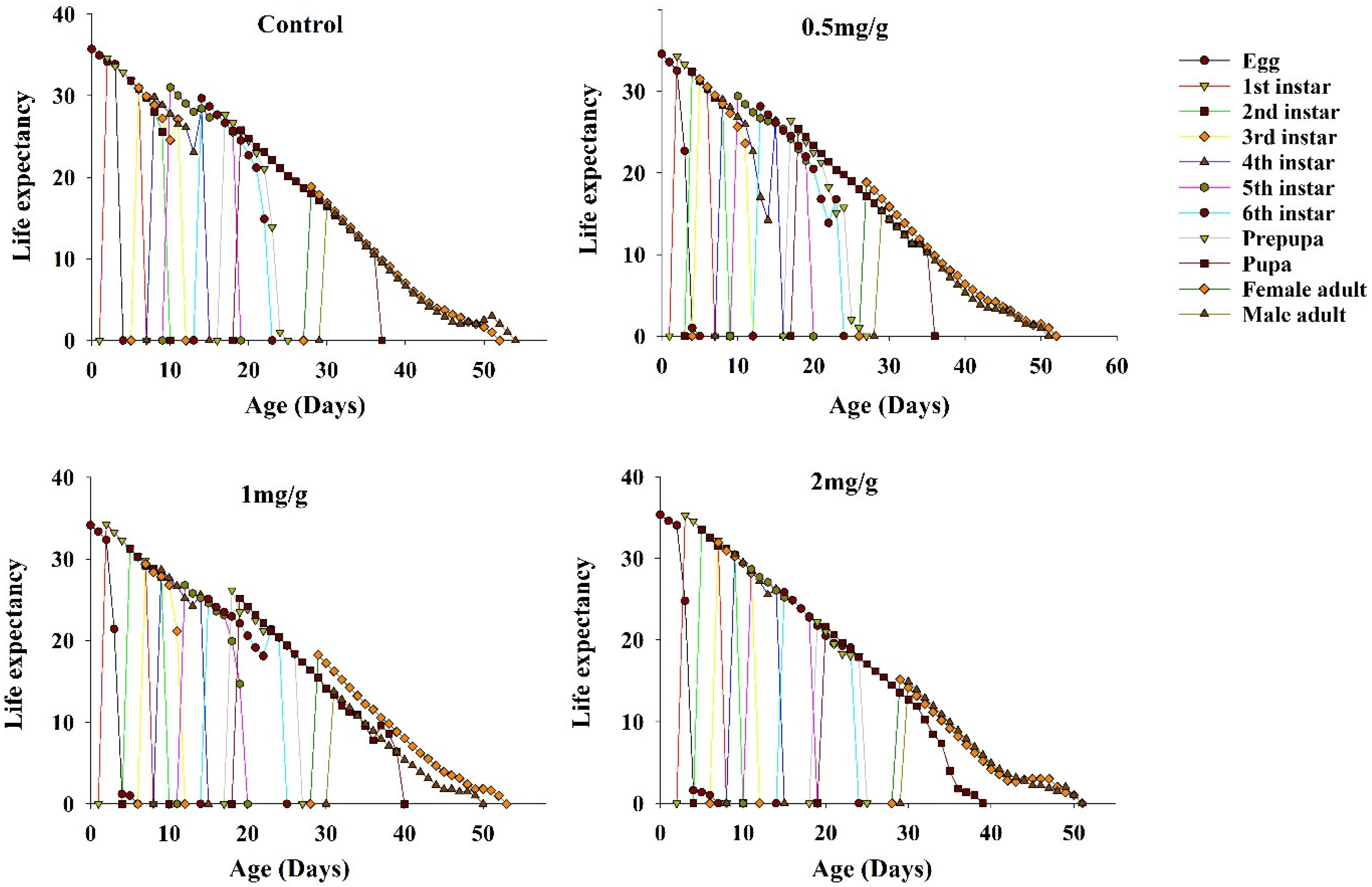
3.6. Life Expectancy of FAWs
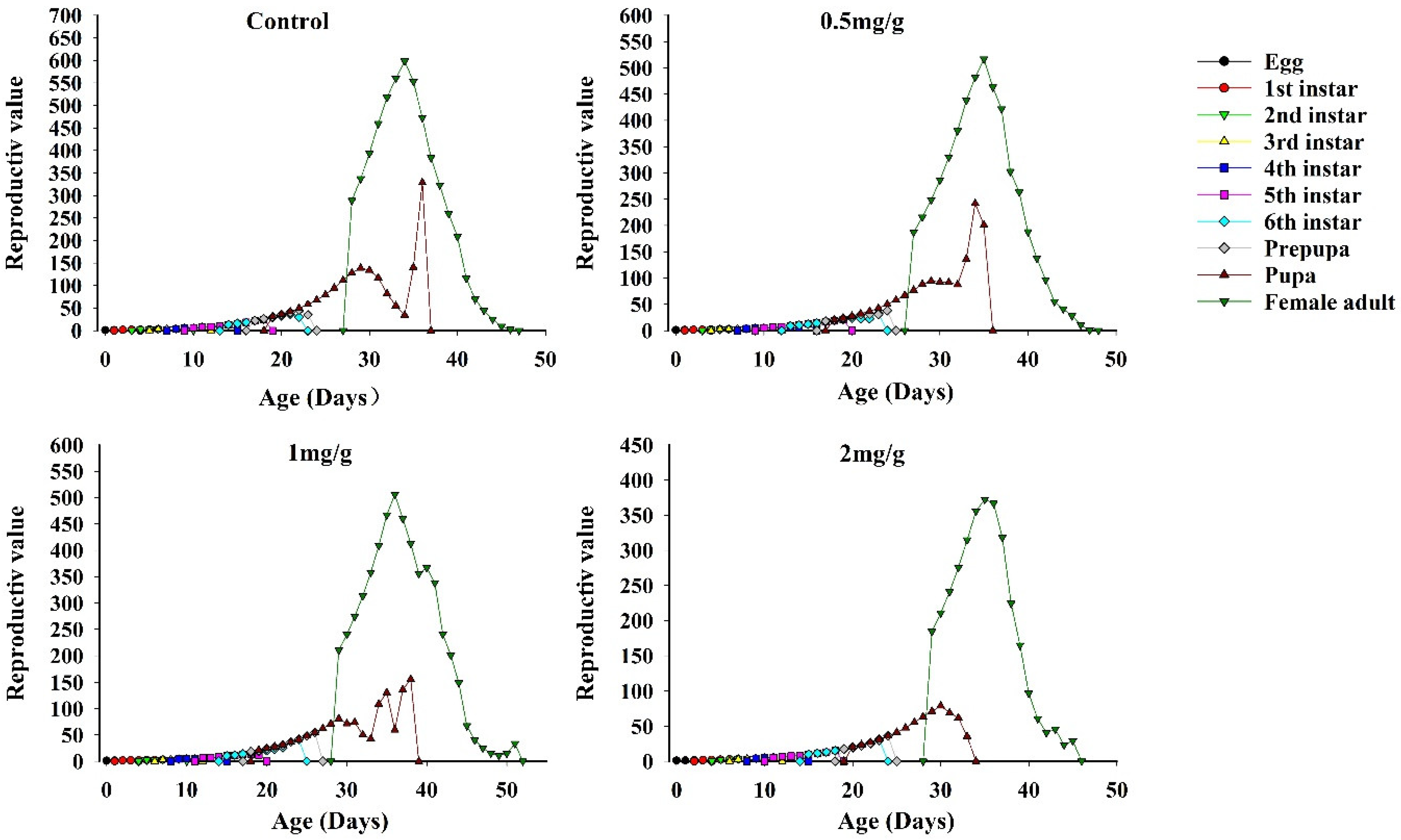
3.7. Reproductive Value of FAWs
3.8. Life Table Parameters
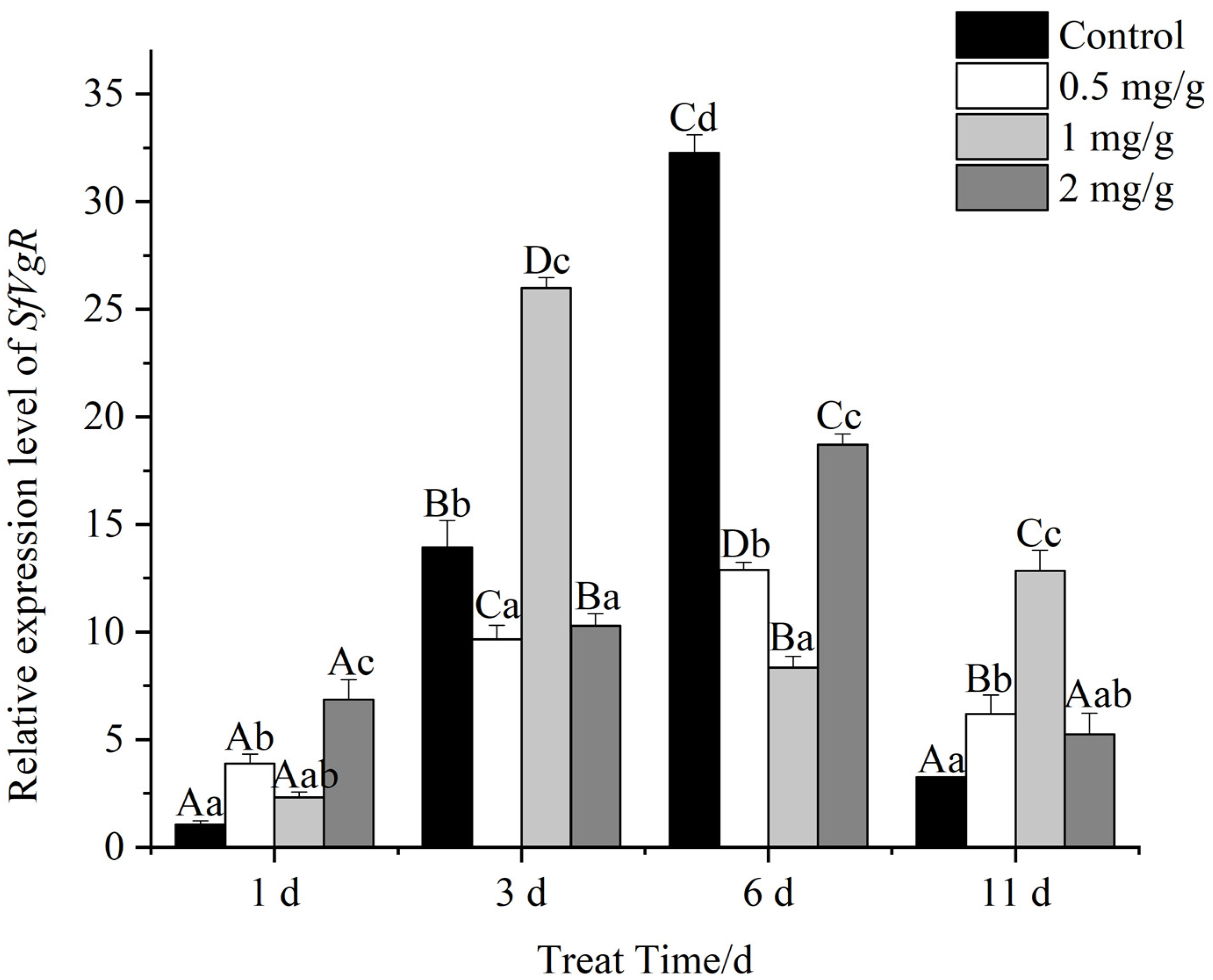
3.9. Expression of SfVg and SfVgR
3.10. Correlation Analysis of Gene Expression Levels and Fecundity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Marchi, L.; Coppola, F.; Soares, A.M.V.M.; Pretti, C.; Monserrat, J.M.; Della Torre, C.; Freitas, R. Engineered nanomaterials: From their properties and applications, to their toxicity towards marine bivalves in a changing environment. Environ. Res. 2019, 178, 108683. [Google Scholar] [CrossRef] [PubMed]
- Du, J.-J.; Zhang, Y.-Y.; Yin, Y.-T.; Zhang, J.; Ma, H.; Li, K.; Wan, N. Do environmental concentrations of zinc oxide nanoparticle pose ecotoxicological risk to aquatic fungi associated with leaf litter decomposition? Water Res. 2020, 178, 115840. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, N.P.; Scott-Fordsmand, J.J.; Amorim, M.J.B. Novel understanding of toxicity in a life cycle perspective–The mechanisms that lead to population effect–The case of Ag (nano) materials. Environ. Pollut. 2020, 262, 114277. [Google Scholar] [CrossRef] [PubMed]
- Malakar, A.; Kanel, S.R.; Ray, C.; Snow, D.D. Nanomaterials in the environment, human exposure pathway, and health effects: A review. Sci. Total Environ. 2021, 759, 143470. [Google Scholar] [CrossRef] [PubMed]
- Wigger, H.; Kagi, R.; Wiesner, M.; Nowack, B. Exposure and possible risks of engineered nanomaterials in the environment—Current knowledge and directions for the future. Rev. Geophys. 2020, 58, e2020RG000710. [Google Scholar] [CrossRef]
- Sreeprasad, T.S.; Pradeep, T. Graphene for environmental and biological applications. Int. J. Mod. Phys. B 2012, 26, 1242001. [Google Scholar] [CrossRef]
- Liu, J.-Q.; Tang, J.-G.; Gooding, J.J. Strategies for chemical modification of graphene and applications of chemically modified graphene. J. Mater. Chem. 2012, 22, 12435–12452. [Google Scholar] [CrossRef]
- Busy, C.; Ali-Boucetta, H.; Kostarelos, K. Safety considerations for graphene: Lessons learnt from carbon nanotubes. Acc. Chem. Res. 2013, 46, 692–701. [Google Scholar] [CrossRef]
- Ali-Boucetta, H.; Bitounis, D.; Raveendran-Nair, R.; Servant, A.; Van den Bossche, J.; Kostarelos, K. Purified graphene oxide dispersions lack in vitro cytotoxicity and in vivo pathogenicity. Adv. Healthc. Mater. 2013, 2, 433–441. [Google Scholar] [CrossRef]
- Hu, X.; Lu, K.; Mu, L.; Kang, J.; Zhou, Q. Interactions between graphene oxide and plant cells: Regulation of cell morphology, uptake, organelle damage, oxidative effects and metabolic disorders. Carbon 2014, 80, 665–676. [Google Scholar] [CrossRef]
- Liu, S.-J.; Wei, H.-M.; Li, Z.-Y.; Li, S.; Yan, H.; He, Y.; Tian, Z.-H. Effects of graphene on germination and seedling morphology in rice. J. Nanosci. Nanotechnol. 2015, 15, 2695–2701. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zeng, T.-H.; Hofmann, M.; Burcombe, E.; Wei, J.; Jiang, R.; Kong, J.; Chen, Y. Antibacterial activity of graphite, graphite oxide, graphene oxide, and reduced graphene oxide: Membrane and oxidative stress. ACS Nano 2011, 5, 6971–6980. [Google Scholar] [CrossRef] [PubMed]
- Akhavan, O.; Ghaderi, E. Toxicity of graphene and graphene oxide nanowalls against bacteria. ACS Nano 2010, 4, 5731–5736. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Singh, S.; Ganguli, A.K.; Shanmugam, V. Anti-drift nano-stickers made of graphene oxide for targeted pesticide delivery and crop pest control. Carbon 2017, 115, 781–790. [Google Scholar] [CrossRef]
- Tong, Y.-J.; Shao, X.-L.; Li, X.-L.; Lu, J.-Q.; Sun, H.-L.; Xiang, S.; Zhang, Z.-H.; Wu, Y.; Wu, X.-M. Adhesive and stimulus-responsive polydopamine-coated graphene oxide system for pesticide-loss control. J. Agric. Food Chem. 2018, 66, 2616–2622. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-P.; Xie, H.-C.; Wang, Z.-Y.; He, K.-L. Graphene oxide as a pesticide delivery vector for enhancing acaricidal activity against spider mites. Colloids Surf. B 2019, 173, 632–638. [Google Scholar] [CrossRef] [PubMed]
- Dziewięcka, M.; Karpeta-Kaczmarek, J.; Augustyniak, M.; Rost-Roszkowska, M. Short-term in vivo exposure to graphene oxide can cause damage to the gut and testis. J. Hazard Mater. 2017, 328, 80–89. [Google Scholar] [CrossRef]
- Jastrzębska, A.M.; Olszyna, A.R. The ecotoxicity of graphene family materials: Current status, knowledge gaps and future needs. J. Nanopart. Res. 2015, 17, 40. [Google Scholar] [CrossRef]
- Sparks, A.N. A review of the biology of the fall armyworm. Fla. Entomol. 1979, 62, 82–87. [Google Scholar] [CrossRef]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Witt, A. Fall armyworm: Impacts and implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Kasoma, C.; Shimelis, H.; Laing, M.D. Fall armyworm invasion in Africa: Implications for maize production and breeding. J. Crop Improv. 2021, 35, 111–146. [Google Scholar] [CrossRef]
- Wightman, J.A. Can lessons learned 30 years ago contribute to reducing the impact of the fall army worm Spodoptera frugiperda in Africa and India? Outlook Agric. 2018, 47, 259–269. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gomez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef]
- Paredes-Sánchez, F.A.; Rivera, G.; Bocanegra-García, V.; Martínez-Padrón, H.Y.; Berrones-Morales, M.; Niño-García, N.; Herrera-Mayorga, V. Advances in Control Strategies against Spodoptera frugiperda. A Review. Molecules 2021, 26, 5587. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-Y.; Dai, Q.-X.; Kuang, Z.-L.; Liang, M.-R.; Wang, L.; Lu, Y.-Y.; Chen, K.-W. Effects of three artificial diets on development and reproduction of the fall armyworm Spodoptera frugiperda (J. E. Smith). J. Environ. Entomol. 2019, 41, 1147–1154. [Google Scholar]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H.; Fu, J.-W.; You, M.-S. Age-stage, two-sex life table and its application in population ecology and integrated pest management. Acta Entomol. Sin. 2019, 62, 255–262. [Google Scholar]
- Chi, H.; Su, H.-Y. Age-stage, two-sex life tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and its host Myzus persicae (Sulzer) (Homoptera: Aphididae) with mathematical proof of the relationship between female fecundity and the net reproductive rate. Environ. Entomol. 2006, 35, 10–21. [Google Scholar] [CrossRef]
- Tuan, S.-J.; Lee, C.-C.; Chi, H. Population and damage projection of Spodoptera litura (F.) on peanuts (Arachis hypogaea L.) under different conditions using the age-stage, two-sex life table. Pest Manag. Sci. 2014, 70, 805–813. [Google Scholar] [CrossRef]
- Yang, Y.-T.; Li, W.-X.; Xie, W.; Wu, Q.-J.; Xu, B.-Y.; Wang, S.-L.; Li, C.-R.; Zhang, Y.-J. Development of Bradysia odoriphaga (Diptera: Sciaridae) as affected by humidity: An age–stage, two-sex, life-table study. Appl. Entomol. Zool. 2015, 50, 3–10. [Google Scholar] [CrossRef]
- Zhao, S.-Y.; Yang, X.-M.; He, W.; Zhang, H.-W.; Jiang, Y.-Y.; Wu, K.-M. Ovarian development gradation and reproduction potential prediction in Spodoptera frugiperda. Plant Prot. 2019, 45, 28–34. [Google Scholar]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age Stage, Two-Sex Life Table Analysis. 2020. Available online: https://140.120.197.173/Ecology/Download/TWOSEX.zip (accessed on 30 July 2020).
- Akca, I.; Ayvaz, T.; Yazici, E.; Smith, C.L.; Chi, H. Demography and population projection of Aphis fabae (Hemiptera: Aphididae): With additional comments on life table research criteria. J. Econ. Entomol. 2015, 108, 1466–1478. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, R.; Ullah, M.S.; Ho, C.C.; Gökçe, A.; Chi, H.; Gotoh, T. Temperature-dependent demography of two closely related predatory mites Neoseiulus womersleyi and N. longispinosus (Acari: Phytoseiidae). J. Econ. Entomol. 2017, 110, 1533–1546. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.H.Z.; De Sousa, M.; Fonseca, L.C.; Martinez, D.S.T.; Alves, O.L. Biological effects of oxidized carbon nanomaterials (1D versus 2D) on Spodoptera frugiperda: Material dimensionality influences on the insect development, performance and nutritional physiology. Chemosphere 2019, 215, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.K.; Qu, X.; Aleman-Meza, B.; Wang, T.; Riepe, C.; Liu, Z.; Li, Q.; Zhong, W. Multi-endpoint, high-throughput study of nanomaterial toxicity in Caenorhabditis elegans. Environ. Sci. Technol. 2015, 49, 2477–2485. [Google Scholar] [CrossRef] [PubMed]
- Afrasiabi, Z.; Popham, H.J.; Stanley, D.; Suresh, D.; Finley, K.; Campbell, J.; Kannan, R.; Upendran, A. Dietary silver nanoparticles reduce fitness in a beneficial, but not pest, insect species. Arch. Insect Biochem. Physiol. 2016, 93, 190–201. [Google Scholar] [CrossRef]
- Lin, C.; Wei, Z.; Yi, Z.; Tingting, T.; Huamao, D.; Lichun, F. Analysis of the effects of nanosilver on bacterial community in the intestinal fluid of silkworms using high-throughput sequencing. Bull. Entomol. Res. 2020, 110, 309–320. [Google Scholar] [CrossRef]
- Schoonhoven, L.M.; Van Loon, B.; van Loon, J.J.A.; Dicke, M. Insect-Plant Biology; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Panacek, A.; Prucek, R.; Safarova, D.; Dittrich, M.; Richtrova, J.; Benickova, K.; Zboril, R.; Kvitek, L. Acute and chronic toxicity effects of silver nanoparticles (NPs) on Drosophila melanogaster. Environ. Sci. Technol. 2011, 45, 4974–4979. [Google Scholar] [CrossRef]
- Ibrahim, A.M.A.; Ali, A.M. Silver and zinc oxide nanoparticles induce developmental and physiological changes in the larval and pupal stages of Spodoptera littoralis (Lepidoptera: Noctuidae). J. Asia-Pac. Entomol. 2018, 21, 1373–1378. [Google Scholar] [CrossRef]
- Liu, X.-Y.; Vinson, D.; Abt, D.; Hurt, R.H.; Rand, D.M. Differential toxicity of carbon nanomaterials in Drosophila: Larval dietary uptake is benign, but adult exposure causes locomotor impairment and mortality. Environ. Sci. Technol. 2009, 43, 6357–6363. [Google Scholar] [CrossRef]
- Marucci, R.C.; Freitas, L.M.; Santos-Rasera, J.R.; Alves, D.S.; Carvalho, G.A.; De-Carvalho, H.W.P. Are cerium oxide nanoparticles transferred from plants to the aphid Myzus persicae (Hemiptera: Aphididae)? Fla. Entomol. 2019, 102, 555–561. [Google Scholar] [CrossRef]
- Wang, X.-P.; Xie, H.-C.; Wang, Z.-Y.; He, K.-L.; Jing, D.-P. Graphene oxide as a multifunctional synergist of insecticides against lepidopteran insect. Environ. Sci. Nano 2019, 6, 75–84. [Google Scholar] [CrossRef]
- Chen, L.-G.; Guo, Y.-Y.; Hu, C.-Y.; Paul, K.S.L.; James, C.W.L.; Zhou, B.-S. Dysbiosis of gut microbiota by chronic coexposure to titanium dioxide nanoparticles and bisphenol A: Implications for host health in zebrafish. Environ. Pollut. 2018, 234, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Judy, J.D.; Unrine, J.M.; Rao, W.; Bertsch, P.M. Bioaccumulation of gold nanomaterials by Manduca sexta through dietary uptake of surface contaminated plant tissue. Environ. Sci. Technol. 2012, 46, 12672–12678. [Google Scholar] [CrossRef] [PubMed]
- Dziewięcka, M.; Witas, P.; Karpeta-Kaczmarek, J.; Kwaśniewska, J.; Flasz, B.; Balin, K.; Augustyniak, M. Reduced fecundity and cellular changes in Acheta domesticus after multigenerational exposure to graphene oxide nanoparticles in food. Sci. Total Environ. 2018, 635, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Shang, F.; Niu, J.-Z.; Ding, B.-Y.; Zhang, Q.; Ye, C.; Zhang, W.; Smagghe, G.; Wang, J.J. Vitellogenin and its receptor play essential roles in the development and reproduction of the brown citrus aphid, Aphis (Toxoptera) citricidus. Insect Mol. Biol. 2018, 27, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Shi, Q.; Guo, Y.; Guo, Y.-Y.; Hua, J.-H.; Wang, X.-F.; Zhou, B.-S. Enhanced bioconcentration of bisphenol A in the presence of nano-TiO2 can lead to adverse reproductive outcomes in zebrafish. Environ. Sci. Technol. 2016, 50, 1005–1013. [Google Scholar] [CrossRef]
- Kotil, T.; Akbulut, C.; Yön, N.D. The effects of titanium dioxide nanoparticles on ultrastructure of zebrafish testis (Danio rerio). Micron 2017, 100, 38–44. [Google Scholar] [CrossRef]
- Xie, J.; Ming, Z.; Li, H.; Yang, H.; Yu, B.; Wu, R.; Liu, X.; Bai, Y.; Yang, S.T. Toxicity of graphene oxide to white rot fungus Phanerochaete chrysosporium. Chemosphere 2016, 151, 324–331. [Google Scholar] [CrossRef]
- Li, P.; Xu, T.; Wu, S.; Lei, L.; He, D. Chronic exposure to graphene-based nanomaterials induces behavioral deficits and neural damage in Caenorhabditis elegans. J. Appl. Toxicol. 2017, 37, 1140–1150. [Google Scholar] [CrossRef]
- Chen, M.; Yin, J.; Liang, Y.; Yuan, S.; Wang, F.; Song, M.; Wang, H. Oxidative stress and immunotoxicity induced by graphene oxide in zebrafish. Aquat. Toxicol. 2016, 174, 54–60. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Ingredient Amount |
|---|---|
| Distilled water | 1200 mL |
| Agar | 24 g |
| Bean powder | 120 g |
| Wheat bran | 120 g |
| Yeast powder | 48 g |
| Casein | 24 g |
| Sorbic acid | 2.4 g |
| Cholesterol | 0.24 g |
| Inositol | 0.24 g |
| Ascorbic acid | 9.6 g |
| Choline chloride | 1.2 g |
| Gene | Primer (5′-3′) |
|---|---|
| RPL27-F | GAAGCCAGGTAAAGTGGTGCT |
| RPL27-R | GTGTCCGTAGGGCTTGTCTG |
| SfVg-F | TCCTCAGTGTTAACGTGCCC |
| SfVg-R | ACAGTCCCTGTTCACGTTCC |
| SfVgR-F | GTCGTGTGGTGGAAGCTGTA |
| SfVgR-R | GCCACCAACATTCTCCCGTA |
| Treatment | Larval Stadium/d | Pre-Adult Time/d | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Egg | 1st Instar | 2nd Instar | 3rd Instar | 4th Instar | 5th Instar | 6th Instar | Prepupal Period | Pupae | ||
| Control | 2.95 ± 0.02 c | 2.62 ± 0.04 c | 1.87 ± 0.03 c | 2.24 ± 0.03 a | 2.44 ± 0.05 a | 4.16 ± 0.06 a | 3.39 ± 0.06 b | 1.74 ± 0.04 ab | 10.39 ± 0.10 a | 31.59 ± 0.17 c |
| 0.5 mg/g | 3.06 ± 0.03 b | 2.78 ± 0.02 b | 1.85 ± 0.03 bc | 2.27 ± 0.04 a | 2.21 ± 0.04 b | 4.35 ± 0.07 a | 3.68 ± 0.06 a | 1.59 ± 0.05 b | 10.44 ± 0.10 a | 31.91 ± 0.19 bc |
| 1 mg/g | 3.10 ± 0.02 b | 3.08 ± 0.02 a | 1.76 ± 0.04 b | 2.31 ± 0.03 a | 2.23 ± 0.04 b | 4.28 ± 0.07 a | 3.66 ± 0.07 a | 1.79 ± 0.04 a | 10.51 ± 0.10 a | 32.47 ± 0.19 a |
| 2 mg/g | 3.15 ± 0.03 a | 3.07 ± 0.02 a | 2.01 ± 0.01 a | 2.03 ± 0.03 a | 2.29 ± 0.04 b | 4.22 ± 0.06 a | 3.59 ± 0.06 a | 1.84 ± 0.04 a | 10.36 ± 0.09 a | 32.38 ± 0.15 ab |
| Treatment | Male Adult Longevity (d) | Female Adult Longevity (d) | Adult Pre-Oviposition Period (d) | Fecundity (Eggs /Female) | Total Life (d) | Pupal Weight (mg) | Sex Ratio (Female/Male) |
|---|---|---|---|---|---|---|---|
| Control | 14.12 ± 0.38 a | 16.04 ± 0.48 a | 4.05 ± 0.15 c | 1029.29 ± 101.81 a | 46.67 ± 0.28 a | 228.29 ± 2.27 a | 0.95:1 b |
| 0.5 mg/g | 12.03 ± 0.49 b | 14.62 ± 0.64 a | 5.41 ± 0.38 b | 810.60 ± 107.08 ab | 45.14 ± 0.38 bc | 227.91 ± 3.31 a | 1.03:1 ab |
| 1 mg/g | 11.65 ± 0.42 bc | 15.64 ± 0.67 a | 6.89 ± 0.60 a | 823.28 ± 102.97 a | 45.89 ± 0.37 ab | 220.45 ± 3.56 a | 0.94:1 b |
| 2 mg/g | 11.87 ± 0.35 bc | 12.48 ± 0.34 b | 5.26 ± 0.30 b | 567.88 ± 73.14 b | 44.55 ± 0.24 c | 193.62 ± 2.97 b | 1.23:1 a |
| Survival Rate % | Life Stage | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Treatment | Egg | 1st Instar | 2nd Instar | 3rd Instar | 4th Instar | 5th Instar | 6th Instar | Prepupae | Pupal |
| Control | 97.33 ± 0.01 c | 99.32 ± 0.01 a | 100.00 ± 0.00 a | 96.56 ± 0.02 a | 88.72 ± 0.04 a | 91.04 ± 0.02 a | 97.46 ± 0.01 b | 97.35 ± 0.01 a | 90.46 ± 0.02 b |
| 0.5 mg/g | 94.67 ± 0.01 b | 100.00 ± 0.00 a | 99.29 ± 0.01 a | 98.61 ± 0.01 a | 89.93 ± 0.01 a | 93.61 ± 0.02 a | 95.70 ± 0.02 b | 93.9 ± 0.03 a | 90.47 ± 0.01 b |
| 1 mg/g | 93.33 ± 0.01 ab | 100.00 ± 0.00 a | 99.29 ± 0.01 a | 95.68 ± 0.01 a | 95.48 ± 0.01 ab | 93.70 ± 0.02 a | 88.21 ± 0.01 a | 96.12 ± 0.02 a | 87.92 ± 0.02 b |
| 2 mg/g | 91.33 ± 0.01 a | 99.26 ± 0.01 a | 98.51 ± 0.01 a | 98.48 ± 0.01 a | 98.48 ± 0.01 b | 96.19 ± 0.01 a | 100.00 ± 0.00 b | 95.97 ± 0.01 a | 78.29 ± 0.01 a |
| Treatment | λ Intrinsic Rate/(d−1) | r Finite Rate/(d−1) | R0 Net Reproduction Rate/Offspring | T(d) Mean Generation Time/d |
|---|---|---|---|---|
| Control | 1.16 ± 0.01 a | 0.15 ± 0.01 a | 329.37 ± 50.67 a | 37.41 ± 0.30 c |
| 0.5 mg/g | 1.15 ± 0.01 ab | 0.14 ± 0.01 ab | 232.37 ± 42.63 ab | 38.47 ± 0.35 b |
| 1 mg/g | 1.14 ± 0.01 b | 0.13 ± 0.01 b | 214.05 ± 39.62 b | 40.38 ± 0.43 a |
| 2 mg/g | 1.14 ± 0.01 b | 0.13 ± 0.01 b | 170.36 ± 30.39 b | 38.42 ± 0.31 b |
| Treat | Total Oviposition Period (d) | Stage Oviposition Quantity | Stage Hatchability | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Stage 1 | Stage 2 | Stage 3 | Stage 4 | Stage 1 | Stage 2 | Stage 3 | Stage 4 | ||
| Control | 18 | 1495.33 ± 937.79 a | 7488.33 ± 2319.71 a | 4163.33 ± 613.10 b | 2076.66 ± 1054.22 a | 0.97 ± 0.06 a | 0.98 ± 0.01 a | 0.98 ± 0.01 a | 0.97 ± 0.01 b |
| 0.5 mg/g | 16 | 3003.00 ± 665.98 a | 4262.66 ± 2092.05 a | 2628.66 ± 566.35 ab | 1837.66 ± 438.11 a | 0.95 ± 0.04 a | 0.97 ± 0.01 a | 0.98 ± 0.01 a | 0.96 ± 0.01 b |
| 1 mg/g | 15 | 3703.66 ± 1112.34 a | 2204.00 ± 805.66 a | 3136.66 ± 621.52 ab | 1475.33 ± 588.77 a | 0.96 ± 0.02 a | 0.98 ± 0.01 a | 0.97 ± 0.01 a | 0.95 ± 0.01 ab |
| 2 mg/g | 12 | 2193.00 ± 1159.23 a | 4386.00 ± 826.18 a | 1483.00 ± 581.13 a | 449.66 ± 135.47 a | 0.96 ± 0.05 a | 0.97 ± 0.01 a | 0.97 ± 0.01 a | 0.93 ± 0.01 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Hu, C.; Zhi, J.; Yue, W.; Li, H. Effects of Nano-Graphene Oxide on the Growth and Reproductive Dynamics of Spodoptera frugiperda Based on an Age-Stage, Two-Sex Life Table. Insects 2022, 13, 929. https://doi.org/10.3390/insects13100929
Li C, Hu C, Zhi J, Yue W, Li H. Effects of Nano-Graphene Oxide on the Growth and Reproductive Dynamics of Spodoptera frugiperda Based on an Age-Stage, Two-Sex Life Table. Insects. 2022; 13(10):929. https://doi.org/10.3390/insects13100929
Chicago/Turabian StyleLi, Cao, Chaoxing Hu, Junrui Zhi, Wenbo Yue, and Hongbo Li. 2022. "Effects of Nano-Graphene Oxide on the Growth and Reproductive Dynamics of Spodoptera frugiperda Based on an Age-Stage, Two-Sex Life Table" Insects 13, no. 10: 929. https://doi.org/10.3390/insects13100929
APA StyleLi, C., Hu, C., Zhi, J., Yue, W., & Li, H. (2022). Effects of Nano-Graphene Oxide on the Growth and Reproductive Dynamics of Spodoptera frugiperda Based on an Age-Stage, Two-Sex Life Table. Insects, 13(10), 929. https://doi.org/10.3390/insects13100929