



Small Islands, Small Ponds, Small Communities—Water Beetles and Water Boatmen in the Faroe Islands


Abstract
Simple Summary
Abstract
1. Introduction
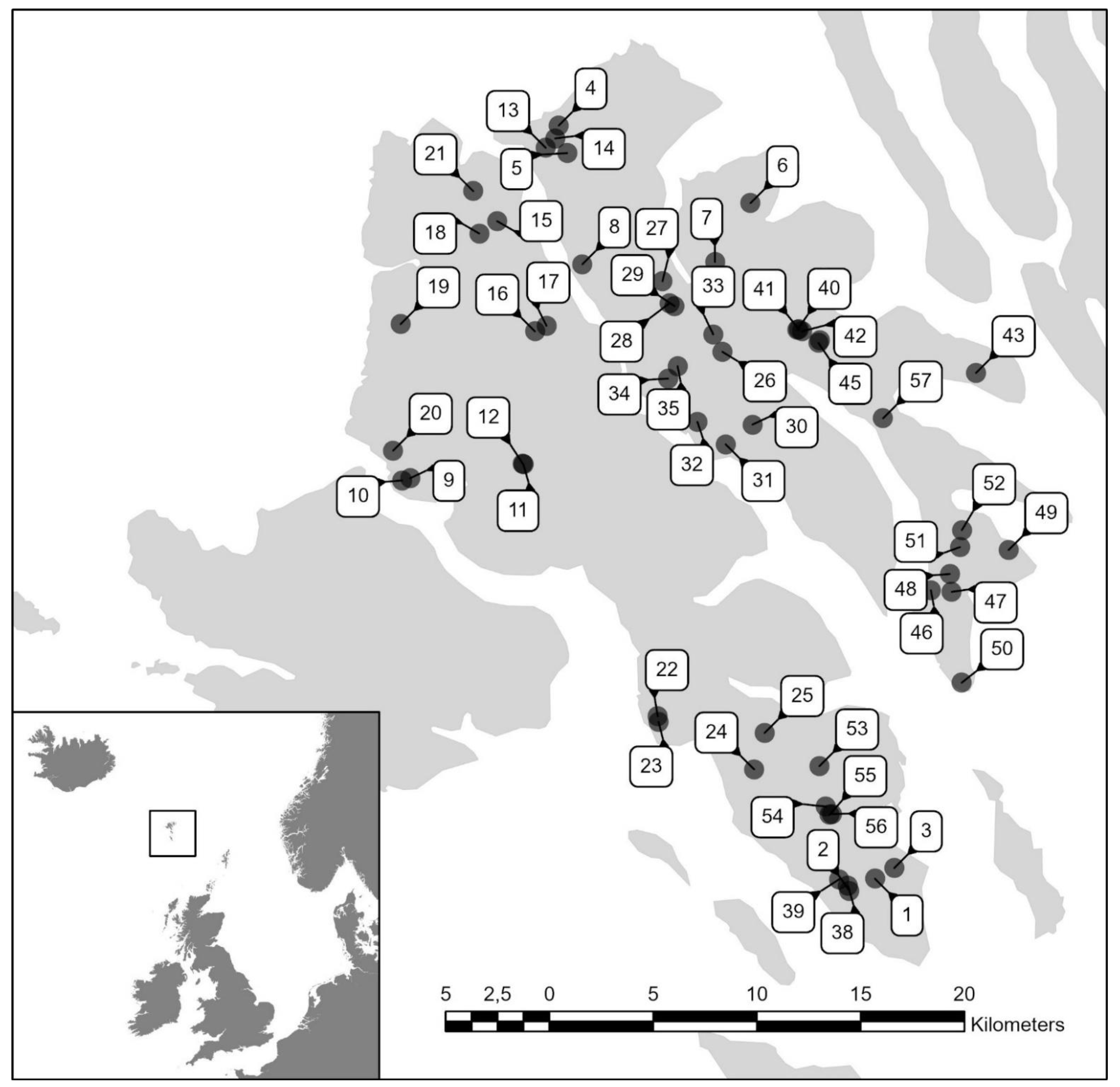
2. Materials and Methods
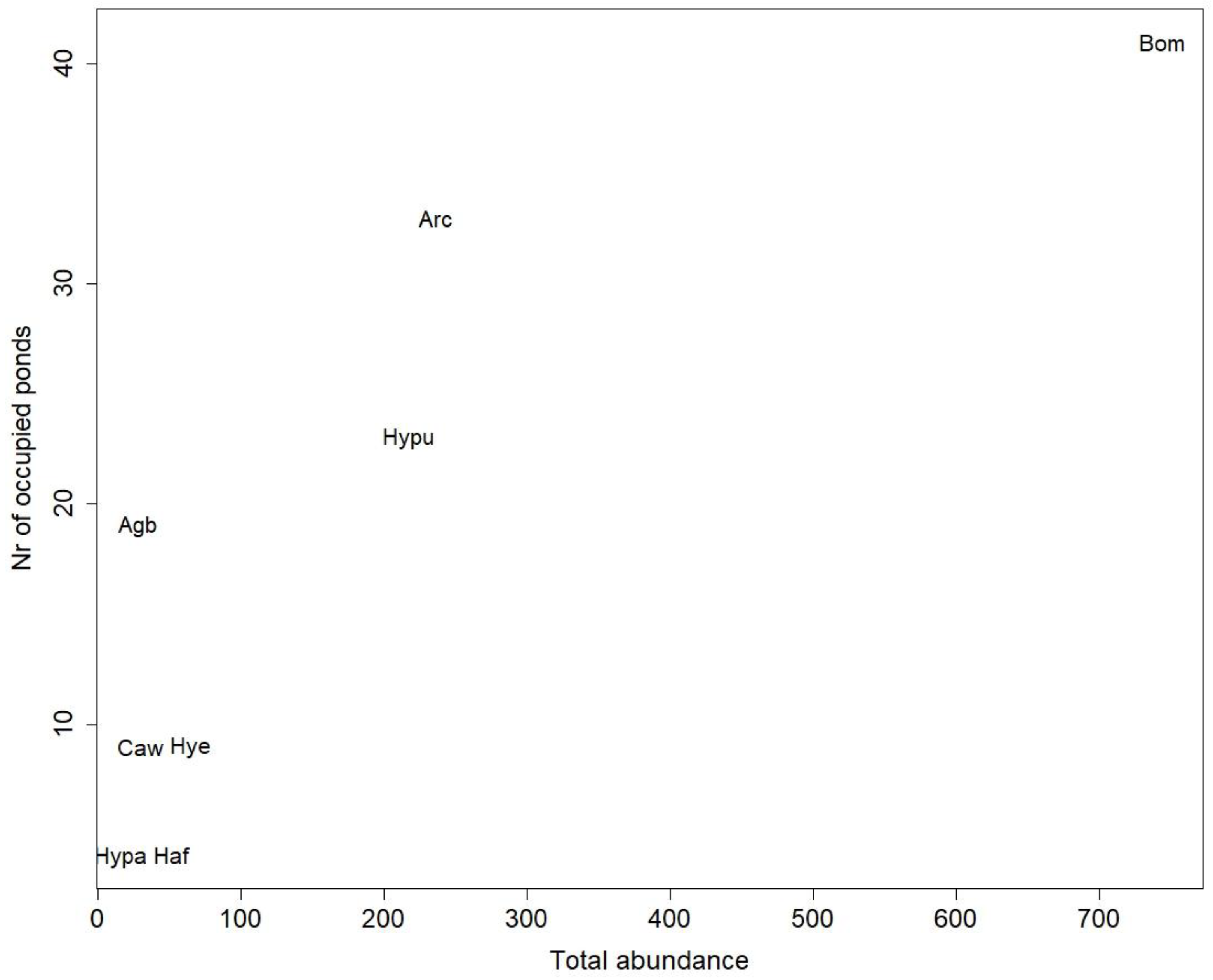
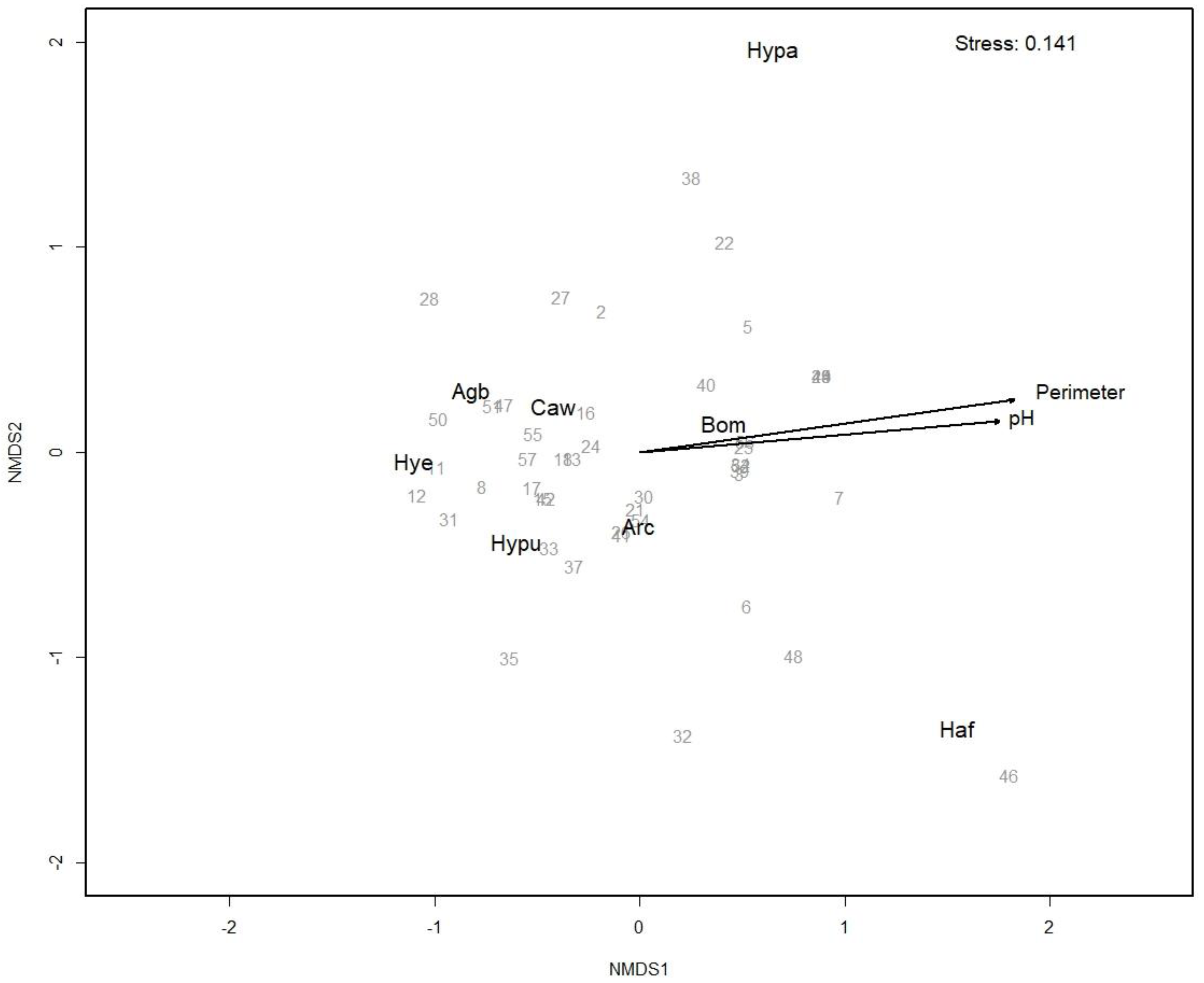
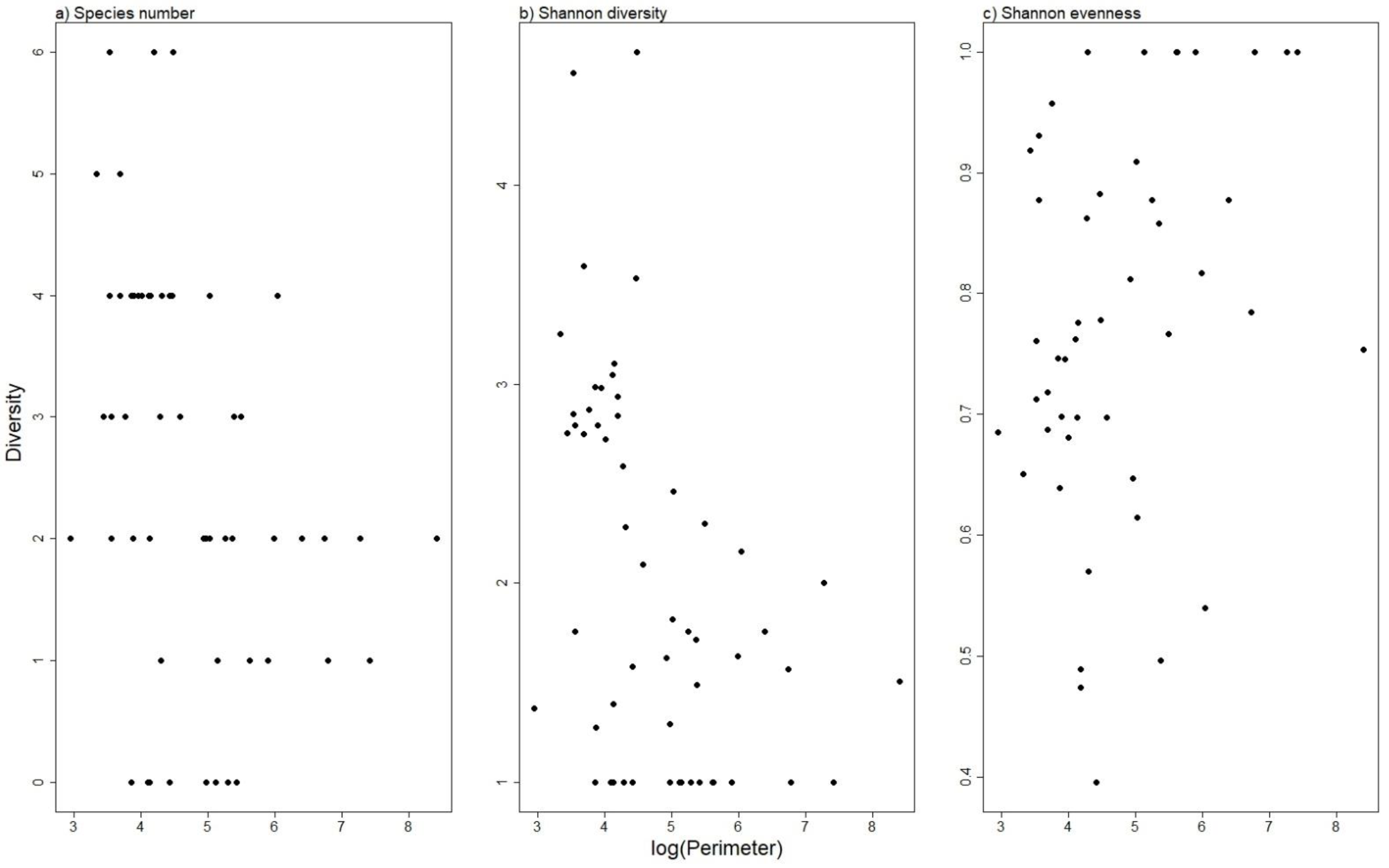
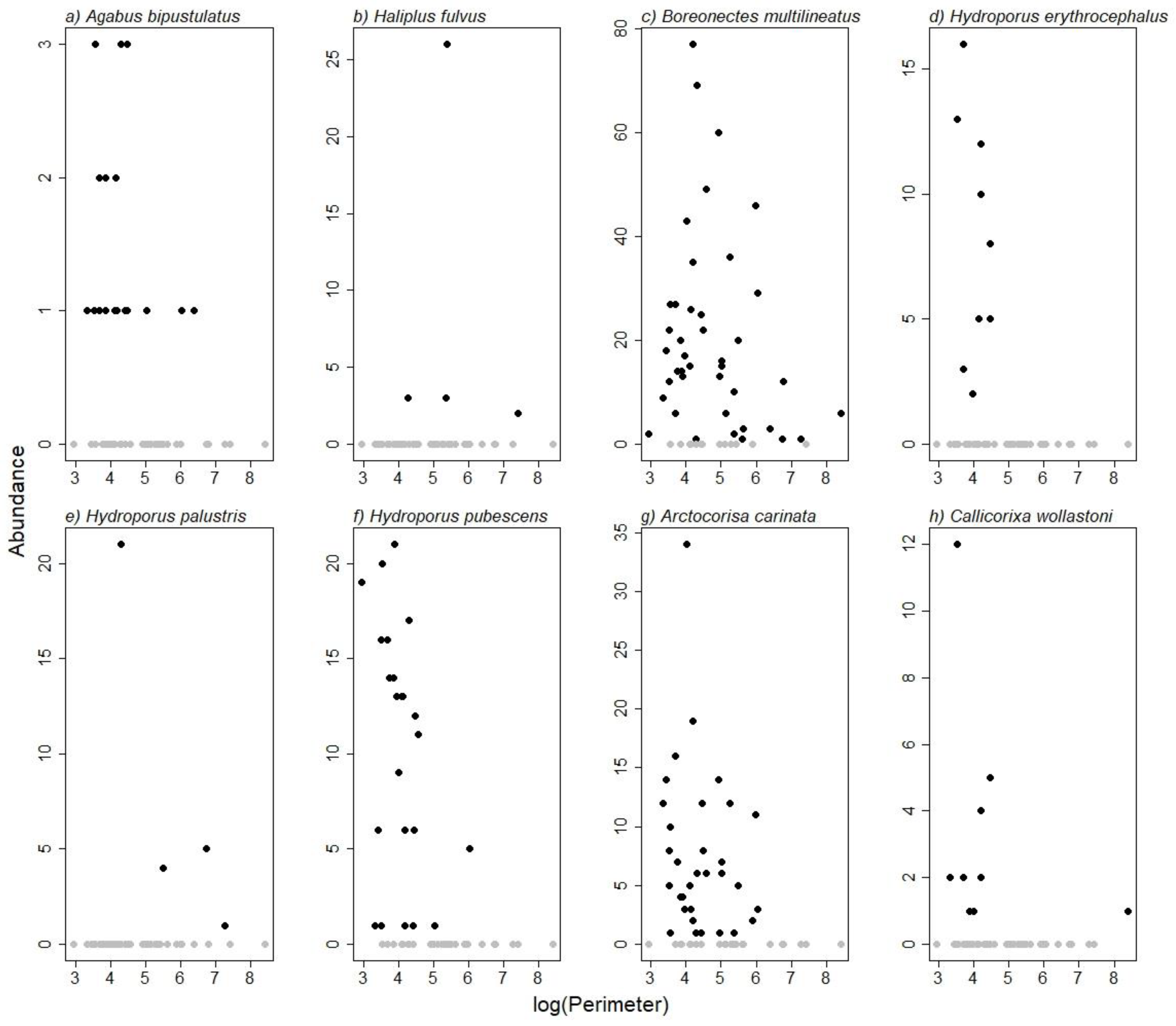
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MacArthur, R.H. Geographical Ecology; Princeton University Press: Princeton, NJ, USA, 1972. [Google Scholar]
- Gíslason, G.M. The Aquatic Fauna of the North Atlantic Islands with Emphasis on Iceland. In Biogeography in the Sub-Arctic; Panagiotakopulu, E., Sadler, J.P., Eds.; Wiley: Hoboken, NJ, USA, 2021; pp. 103–112. [Google Scholar]
- Hansen, L.J.; Gíslason, G.M. Physical Nature of Streams in the Faroe Islands with Notes on Their Biota. Ann. Soc. Sci. Færoensis Suppl. 2010, 52, 259–287. [Google Scholar]
- Bengtson, S.-A. Terrestrial Invertebrates of the Faroe Islands: III. Beetles (Coleoptera): Check-List, Distribution, and Habitats. Fauna Nor. Ser. B 1981, 28, 52–82. [Google Scholar]
- Gíslason, G.M. Origin of Freshwater Fauna of the North-Atlantic Islands: Present Distribution in Relation to Climate and Possible Migration Routes. Verh. Int. Ver. Limnol. 2005, 29, 198–203. [Google Scholar] [CrossRef]
- Panagiotakopulu, E. Hitchhiking across the North Atlantic—Insect Immigrants, Origins, Introductions and Extinctions. Quat. Int. 2014, 341, 59–68. [Google Scholar] [CrossRef]
- Christoffersen, K. Previous Studies of Freshwater Biota in Faroese Lakes. Ann. Soc. Sci. Færoensis Suppl. 2002, 36, 7–13. [Google Scholar]
- Brown, E.S. The Corixidæ of the Faeroe Islands, with Observations on the Geographical Distribution of Corixidæ in Neighbouring Parts of the World. Proc. Zool. Soc. Lond. 1945, 114, 490–506. [Google Scholar] [CrossRef]
- Foster, G.N.; Hansen, J. Wetland Coleoptera of the Faroe Islands. Fróðskaparrit 2004, 52, 42–53. [Google Scholar]
- Hansen, J.; Foster, G.N. Elodes pseudominuta (Coleoptera: Scirtidae) in the Faroe Islands. Fróðskaparrit 2009, 57, 113–114. [Google Scholar]
- Nilsson, A.N. Community Structure in the Dytiscidae (Coleoptera) of a Northern Swedish Seasonal Pond. Ann. Zool. Fenn. 1986, 23, 39–47. [Google Scholar]
- Holmen, M. The Aquatic Adephaga (Coleoptera) of Fennoscandia and Denmark. I: Gyrinidae, Haliplidae, Hygrobiidae and Noteridae. In Fauna Entomologica Scandinavica: 20; E. J. Brill: Leiden, The Netherlands, 1987. [Google Scholar]
- Jansson, A. Heteroptera Nepomorpha, Aquatic Bugs. In The Aquatic Insects of North Europe 1; Nilsson, A., Ed.; Apollo Books Aps.: Stenstrup, Denmark, 1996; pp. 91–104. [Google Scholar]
- Nilsson, A.N.; Holmen, M. The Aquatic Adephaga (Coleoptera) of Fennoscandia and Denmark. II: Dytiscidea. In Fauna Entomologica Scandinavica: 32; E. J. Brill: Leiden, The Netherlands, 1995. [Google Scholar]
- Balfour-Browne, F. British Water Beetles; Ray Society: London, UK, 1940; Volume 1. [Google Scholar]
- Balfour-Browne, F. British Water Beetles; Ray Society: London, UK, 1950; Volume 2. [Google Scholar]
- Savage, A.A. Adults of British Aquatic Hemiptera Heteroptera; Freshwater Biological Association: Ambleside, UK, 1989. [Google Scholar]
- Hanski, I.; Gilpin, M. Metapopulation Dynamics: Brief History and Conceptual Domain. Biol. J. Linn. Soc. 1991, 42, 3–16. [Google Scholar] [CrossRef]
- Flechtner, G. Association Analysis of Water Beetle Communities (Coleoptera: Dytiscidae et Haliplidae). Entomogica Basiliensia 1986, 11, 297–308. [Google Scholar]
- Nilsson, A.N.; Elmberg, J.; Sjöberg, K. Abundance and Species Richness Patterns of Predaceous Diving Beetles (Coleoptera, Dytiscidae) in Swedish Lakes. J. Biogeogr. 1994, 21, 197. [Google Scholar] [CrossRef]
- Ranta, E. Communities of Water-Beetles in Different Kinds of Waters in Finland. Proc. Acad. Nat. Sci. Phila. 1985, 137, 33–45. [Google Scholar]
- Eyre, M.D.; Foster, G.N.; Foster, A.P. Factors Affecting the Distribution of Water Beetle Species Assemblages in Drains of Eastern England. J. Appl. Entomol. 1990, 109, 217–225. [Google Scholar] [CrossRef]
- Juliano, S.A. Changes in Structure and Composition of an Assemblage of Hydroporus Species (Coleoptera: Dytiscidae) along a PH Gradient. Freshw. Biol. 1991, 25, 367–378. [Google Scholar] [CrossRef]
- Lancaster, J.; Scudder, G.G.E. Aquatic Coleoptera and Hemiptera in Some Canadian Saline Lakes: Patterns in Community Structure. Can. J. Zool. 1987, 65, 1383–1390. [Google Scholar] [CrossRef]
- Larson, D.J. Structure in Temperate Predaceous Diving Beetle Communities (Coleoptera: Dytiscidae). Ecography 1985, 8, 18–32. [Google Scholar] [CrossRef]
- Nilsson, A.N.; Soderberg, H. Abundance and Species Richness Patterns of Diving Beetles (Coleoptera, Dytiscidae) from Exposed and Protected Sites in 98 Northern Swedish Lakes. Hydrobiologia 1996, 321, 83–88. [Google Scholar] [CrossRef]
- Lillie, R.A. The Adult and Semiaquatic Coleoptera of Nine Northwestern Wisconsin Wetlands. Coleopt. Bull. 1991, 45, 101–111. [Google Scholar]
- Timms, B.V.; Hammer, U.T. Water Beetles of Some Saline Lakes in Saskatchewan. Can. Field-Nat. 1988, 102, 246–250. [Google Scholar]
- Eyre, M.D.; Carr, R.; Mcblane, R.P.; Foster, G.N. The Effects of Varying Site-Water Duration on the Distribution of Water Beetle Assemblages, Adults and Larvae (Coleoptera, Haliplidae Dytiscidae, Hydrophilidae). Arch. Hydrobiol. 1992, 124, 281–291. [Google Scholar] [CrossRef]
- Jeffries, M. Invertebrate Communities and Turnover in Wetland Ponds Affected by Drought. Freshw. Biol. 1994, 32, 603–612. [Google Scholar] [CrossRef]
- Landin, J. Seasonal Patterns in Abundance of Water-Beetles Belonging to the Hydrophiloidea (Coleoptera). Freshw. Biol. 1976, 6, 89–108. [Google Scholar] [CrossRef]
- Robert, L.L.; Matta, J.F. Aquatic Macroinvertebrates in an Irregularly Flooded Salt Marsh: Diversity and Seasonal Variation. Environ. Entomol. 1984, 13, 1097–1104. [Google Scholar] [CrossRef]
- Ribera, I.; Foster, G.N.; Vogler, A.P. Does Habitat Use Explain Large Scale Species Richness Patterns of Aquatic Beetles in Europe? Ecography 2003, 26, 145–152. [Google Scholar] [CrossRef]
- Eyre, M.D.; Foster, G.N.; Young, A.G. Relationships between Water-Beetle Distributions and Climatic Variables—A Possible Index for Monitoring Global Climatic-Change. Arch. Hydrobiol. 1993, 127, 437–450. [Google Scholar] [CrossRef]
- Islands, Mountains, Islets Og Lakes | Statistics Faroe Islands. Available online: https://hagstova.fo/en/environment/geography/islands-mountains-islets-og-lakes (accessed on 13 September 2022).
- Føroyar Topografisk Kort, 3rd ed.; (Maps 1:20,000); Kort og Matrikelstyrelsen: Denmark, 1989.
- Fosaa, A.M.; Gaard, E.; Dalsgarð, J. Føroya Náttura. In Føroya Náttura—Lívfrøðiligt Margfeldi; Fosaa, A.M., Gaard, E., Dalsgarð, J., Eds.; Føroya Skúlabókagrunnur: Tórshavn, Faroe Islands, 2006; pp. 1–9. [Google Scholar]
- Richardson, D.C.; Holgerson, M.A.; Farragher, M.J.; Hoffman, K.K.; King, K.B.S.; Alfonso, M.B.; Andersen, M.R.; Cheruveil, K.S.; Coleman, K.A.; Farruggia, M.J.; et al. A Functional Definition to Distinguish Ponds from Lakes and Wetlands. Sci. Rep. 2022, 12, 1–13. [Google Scholar] [CrossRef]
- Nilsson, A.N. Coleoptera Haliplidae, Crawling Water Beetles. In The Aquatic Insects of North Europe 1; Nilsson, A., Ed.; Apollo Books Aps.: Stenstrup, Denmark, 1996; pp. 131–138. [Google Scholar]
- Nilsson, A. (Ed.) Aquatic Insects of North Europe 1; Apollo Books: Stenstrup, Denmark, 2005; ISBN 87-88757-55-2. [Google Scholar]
- Mapinfo Corporation. Mapinfo Professional 6.5.0.3100; Mapinfo Corporation: Troy, New York, USA, 2002. [Google Scholar]
- ESRI. ArcGIS Pro 3.0; ESRI: Redlands, CA, USA, 2022. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing 2022; R Foundation for Statistical Computing: Vienna, Austria.
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. R Package Version 2.6-2 2022. Available online: https://github.com/vegandevs/vegan (accessed on 15 August 2022).
- Peterson, B.G.; Carl, P. PerformanceAnalytics: Econometric Tools for Performance and Risk Analysis. R Package Version 2.0.4 2020. Available online: https://github.com/vegandevs/vegan (accessed on 15 August 2022).
- West, A. Coleoptera. In The Zoology of the Faroes, Volume II, Part I; Jensen, A.S., Lundbeck, W., Mortensen, T., Spärck, R., Eds.; Andr. Fred. Høst & Søn: Copenhagen, Denmark, 1937; pp. 1–92. [Google Scholar]
- Enckell, P.H. Island Life: Agency of Man upon Dispersal, Distribution, and Genetic Variation in Faroese Populations of Terrestrial Invertebrates; University of Lund: Lund, Sweden, 1985. [Google Scholar]
- Spärck, R.; Tuxen, S.L.; Jensen, A.S.; Lundbeck, W.; Mortensen, T. (Eds.) The Zoology of the Faroes; Andr. Fred. Høst & Søn: Copenhagen, Denmark, 1928. [Google Scholar]
- Beetle Families|UK Beetle Recording. Available online: https://www.coleoptera.org.uk/beetle-families (accessed on 16 September 2022).
- Berry, R.J. The Natural History of Orkney; Collins: London, UK, 1985. [Google Scholar]
- Pennington, M.G. A Naturalist’s Shetland; Johnston, J.L., Ed.; T & A D Poyser: London, UK, 1999. [Google Scholar]
- Ólafsson, E. Islenskt Skordýratal. Fjölrit Náttúrufræðistofnunar 1991, 17, 1–69. [Google Scholar]
- Larson, D.J.; House, N.L. Insect Communities of Newfoundland Bog Pools with Emphasis on the Odonata. Can. Entomol. 1990, 122, 469–501. [Google Scholar] [CrossRef]
- Kholin, S.K.; Nilsson, A.N. Regional Enrichment of Predacious Water Beetles in Temporary Ponds at Opposite East–West Ends of the Palearctic. J. Biogeogr. 1998, 25, 47–55. [Google Scholar] [CrossRef][Green Version]
- Jeppesen, E.; Christoffersen, K.; Malmquist, H.J.; Faafeng, B.; Hansson, L.-A. Ecology of Five Faroese Lakes: Summary and Synthesis. Ann. Soc. Sci. Færoensis Suppl. 2002, 36, 126–139. [Google Scholar]
- Fairchild, G.W.; Cruz, J.; Faulds, A.M.; Short, A.E.Z.; Matta, J.F. Microhabitat and Landscape Influences on Aquatic Beetle Assemblages in a Cluster of Temporary and Permanent Ponds. J. North Am. Benthol. Soc. 2003, 22, 224–240. [Google Scholar] [CrossRef]
- Gutiérrez-Estrada, J.C.; Bilton, D.T. A Heuristic Approach to Predicting Water Beetle Diversity in Temporary and Fluctuating Waters. Ecol. Model. 2010, 221, 1451–1462. [Google Scholar] [CrossRef]
- Urban, M.C. Disturbance Heterogeneity Determines Freshwater Metacommunity Structure. Ecology 2004, 85, 2971–2978. [Google Scholar] [CrossRef]
- Ribera, I.; Vogler, A.P. Habitat Type as a Determinant of Species Range Sizes: The Example of Lotic–Lentic Differences in Aquatic Coleoptera. Biol. J. Linn. Soc. 2000, 71, 33–52. [Google Scholar] [CrossRef]
- Fairchild, G.W.; Faulds, A.M.; Matta, J.F. Beetle Assemblages in Ponds: Effects of Habitat and Site Age. Freshw. Biol. 2000, 44, 523–534. [Google Scholar] [CrossRef]
- Winkelmann, C.; Hellmann, C.; Worischka, S.; Petzoldt, T.; Benndorf, J. Fish Predation Affects the Structure of a Benthic Community. Freshw. Biol. 2011, 56, 1030–1046. [Google Scholar] [CrossRef]
- Liao, W.; Venn, S.; Niemelä, J. Environmental Determinants of Diving Beetle Assemblages (Coleoptera: Dytiscidae) in an Urban Landscape. Biodivers. Conserv. 2020, 29, 2343–2359. [Google Scholar] [CrossRef]
- Nilsson, A.N.; Svensson, B.W. Assemblages of Dytiscid Predators and Culicid Prey in Relation to Environmental Factors in Natural and Clear-Cut Boreal Swamp Forest Pools. Hydrobiologia 1995, 308, 183–196. [Google Scholar] [CrossRef]
- Hanski, I. Distributional Ecology of Anthropochorous Plants in Villages Surrounded by Forest. Ann. Bot. Fenn. 1982, 19, 1–15. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pond ID | Latitude | Longitude | pH | Temperature | Sed. Depth | Wat. Depth | Perimeter | Dist. Pond | Altitude | Spec.rich. | Shan.div. | Evenness | Agb | Arc | Caw | Haf | Bom | Hye | Hypa | Hypu |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 61.9766 | −6.7842 | 6.42 | 12.2 | 0.5 | 1.5 | 60 | 8 | 248 | 0 | – | – | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 2 | 61.9716 | −6.8081 | 5.87 | 19.9 | 0 | 0.3 | 48 | 94 | 320 | 2 | 1.28 | 0.64 | 1 | 0 | 0 | 0 | 14 | 0 | 0 | 0 |
| 3 | 61.9809 | −6.7663 | 5.66 | 13.7 | 0.5 | 0.4 | 151 | 98 | 178 | 2 | 1.82 | 0.91 | 0 | 6 | 0 | 0 | 15 | 0 | 0 | 0 |
| 4 | 62.3064 | −7.0540 | 6.01 | 18.2 | 0 | 0 | 83 | 43 | 162 | 0 | – | – | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 5 | 62.2943 | −7.0469 | – | 15 | 0 | 0.4 | 4492 | 537 | 193 | 2 | 1.51 | 0.75 | 0 | 0 | 1 | 0 | 6 | 0 | 0 | 0 |
| 6 | 62.2703 | −6.8783 | 6.8 | 10.3 | 0 | 0.3 | 72 | 1305 | 270 | 3 | 2.59 | 0.86 | 0 | 1 | 0 | 3 | 1 | 0 | 0 | 0 |
| 7 | 62.2453 | −6.9130 | 6.6 | 12.3 | 0 | 0.5 | 212 | 1905 | 353 | 2 | 1.72 | 0.86 | 0 | 0 | 0 | 3 | 10 | 0 | 0 | 0 |
| 8 | 62.2462 | −7.0360 | 5.02 | 20.8 | 0.5 | 0.6 | 88 | 8 | 215 | 6 | 4.67 | 0.78 | 1 | 8 | 5 | 0 | 22 | 8 | 0 | 12 |
| 9 | 62.1558 | −7.2010 | 7.01 | 13.9 | 0 | 1.5 | 226 | 341 | 249 | 0 | – | – | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 10 | 62.1549 | −7.2087 | 6.68 | 9 | 0.4 | 1 | 60 | 341 | 236 | 0 | – | – | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 11 | 62.1604 | −7.0961 | 5.07 | 10.3 | 0 | 0.5 | 87 | 18 | 364 | 4 | 3.53 | 0.88 | 3 | 12 | 0 | 0 | 0 | 5 | 0 | 6 |
| 12 | 62.1606 | −7.0966 | 5.07 | 9.2 | 0 | 0.5 | 63 | 18 | 365 | 4 | 3.10 | 0.78 | 2 | 3 | 0 | 0 | 0 | 5 | 0 | 13 |
| 13 | 62.2969 | −7.0668 | 5.94 | 16.9 | 0.05 | 0.2 | 420 | 15 | 190 | 4 | 2.16 | 0.54 | 1 | 3 | 0 | 0 | 29 | 0 | 0 | 5 |
| 14 | 62.3009 | −7.0577 | – | 15 | 0 | 0.5 | 73 | 240 | 169 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 21 | 0 |
| 15 | 62.2659 | −7.1139 | 6.13 | 18.1 | 0.2 | 0.2 | 61 | 33 | 334 | 4 | 3.05 | 0.76 | 1 | 5 | 0 | 0 | 15 | 0 | 0 | 13 |
| 16 | 62.2177 | −7.0818 | 6.68 | 18.5 | 0.1 | 0.5 | 83 | 409 | 406 | 4 | 1.58 | 0.40 | 1 | 1 | 0 | 0 | 25 | 0 | 0 | 1 |
| 17 | 62.2200 | −7.0709 | – | 0 | 0 | 0 | 47 | 117 | 340 | 4 | 2.99 | 0.75 | 2 | 4 | 0 | 0 | 20 | 0 | 0 | 14 |
| 18 | 62.2607 | −7.1308 | 6.33 | 13.8 | 0.1 | 0.3 | 74 | 22 | 421 | 4 | 2.28 | 0.57 | 3 | 6 | 0 | 0 | 69 | 0 | 0 | 17 |
| 19 | 62.2225 | −7.2061 | – | 7 | 0 | 0.4 | 47 | 22 | 487 | 0 | – | – | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 20 | 62.168 | −7.2162 | 6.73 | 10 | 0 | 1.5 | 62 | 461 | 168 | 0 | – | – | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 21 | 62.2791 | −7.1352 | 5.65 | 11.6 | 0.1 | 0.4 | 55 | 8 | 338 | 4 | 2.72 | 0.68 | 0 | 34 | 1 | 0 | 43 | 0 | 0 | 9 |
| 22 | 62.0498 | −6.9791 | – | 11 | 0 | 1.5 | 1438 | 36 | 245 | 2 | 2 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 |
| 23 | 62.0474 | −6.9785 | 6.48 | 13 | 0.2 | 0.5 | 274 | 36 | 247 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| 24 | 62.0256 | −6.8919 | 5.87 | 14.1 | 0 | 0.4 | 152 | 493 | 470 | 4 | 2.46 | 0.61 | 1 | 7 | 0 | 0 | 16 | 0 | 0 | 1 |
| 25 | 62.0412 | −6.8813 | 6.29 | 11.9 | 0.05 | 0.2 | 62 | 95 | 340 | 2 | 1.39 | 0.70 | 0 | 3 | 0 | 0 | 26 | 0 | 0 | 0 |
| 26 | 62.2064 | −6.9088 | 5.12 | 13.4 | 0.1 | 0.2 | 35 | 49 | 393 | 3 | 2.79 | 0.93 | 0 | 10 | 0 | 0 | 27 | 0 | 0 | 20 |
| 27 | 62.2377 | −6.9622 | 6.71 | 9.6 | 0 | 1.5 | 599 | 128 | 324 | 2 | 1.75 | 0.88 | 1 | 0 | 0 | 0 | 3 | 0 | 0 | 0 |
| 28 | 62.2281 | −6.9563 | 5.03 | 11.9 | 0.1 | 0.4 | 35 | 45 | 380 | 2 | 1.75 | 0.88 | 3 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 29 | 62.2268 | −6.9518 | 6.61 | 12.4 | 0.3 | 0.4 | 198 | 112 | 338 | 0 | – | – | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 30 | 62.1745 | −6.8832 | 5.05 | 12.9 | 0.1 | 0.3 | 97 | 11 | 403 | 3 | 2.09 | 0.70 | 0 | 6 | 0 | 0 | 49 | 0 | 0 | 11 |
| 31 | 62.1664 | −6.9085 | 6.11 | 18.3 | 0 | 0.1 | 40 | 13 | 232 | 4 | 2.75 | 0.69 | 1 | 0 | 0 | 0 | 6 | 3 | 0 | 16 |
| 32 | 62.1764 | −6.9340 | 6.76 | 17.5 | 0.1 | 1 | 364 | 93 | 208 | 1 | 1 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 |
| 33 | 62.2140 | −6.9168 | - | 10 | 0 | 0.1 | 52 | 65 | 218 | 4 | 2.98 | 0.75 | 0 | 3 | 0 | 0 | 17 | 2 | 0 | 13 |
| 34 | 62.1956 | −6.9599 | 6.53 | 11.8 | 0 | 0.4 | 138 | 17 | 347 | 2 | 1.62 | 0.81 | 0 | 14 | 0 | 0 | 60 | 0 | 0 | 0 |
| 35 | 62.2008 | −6.9506 | 5.56 | 12 | 0.4 | 0.4 | 19 | 59 | 406 | 2 | 1.37 | 0.68 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 19 |
| 36 | 61.986 | −6.8317 | 6.53 | 16.2 | 0.2 | 0.5 | 226 | 46 | 205 | 0 | – | – | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 37 | 61.9875 | −6.8280 | 5.67 | 0 | 0 | 0.4 | 49 | 11 | 210 | 4 | 2.79 | 0.70 | 0 | 4 | 1 | 0 | 13 | 0 | 0 | 21 |
| 38 | 61.9740 | −6.8093 | 6.25 | 14.4 | 0 | 1 | 842 | 123 | 326 | 2 | 1.57 | 0.78 | 0 | 0 | 0 | 0 | 1 | 0 | 5 | 0 |
| 39 | 61.9769 | −6.8171 | 6.63 | 11.9 | 0.5 | 0.3 | 191 | 30 | 274 | 2 | 1.75 | 0.88 | 0 | 12 | 0 | 0 | 36 | 0 | 0 | 0 |
| 40 | 62.2149 | −6.8386 | - | 5 | 0 | 0.4 | 243 | 16 | 497 | 3 | 2.30 | 0.77 | 0 | 5 | 0 | 0 | 20 | 0 | 4 | 0 |
| 41 | 62.2150 | −6.8373 | 5.86 | 19.4 | 0.05 | 0.3 | 43 | 5 | 500 | 3 | 2.87 | 0.96 | 0 | 7 | 0 | 0 | 14 | 0 | 0 | 14 |
| 42 | 62.2141 | −6.8343 | - | 10 | 0.2 | 0.3 | 34 | 28 | 490 | 4 | 2.85 | 0.71 | 1 | 5 | 0 | 0 | 22 | 0 | 0 | 16 |
| 43 | 62.1935 | −6.6749 | 6.83 | 13.8 | 0.1 | 0.4 | 166 | 559 | 290 | 0 | – | – | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 44 | 62.2100 | −6.8181 | 6.7 | 13.2 | 0 | 1.5 | 883 | 29 | 366 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 12 | 0 | 0 | 0 |
| 45 | 62.2089 | −6.8196 | – | 8 | 0 | 0 | 170 | 10 | 366 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 6 | 0 | 0 | 0 |
| 46 | 62.1004 | −6.7236 | 6.7 | 13.9 | 0 | 1.5 | 1666 | 5 | 16 | 1 | 1 | 1 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 |
| 47 | 62.0994 | −6.7045 | 6.23 | 20.1 | 0.05 | 0.3 | 40 | 112 | 144 | 5 | 3.59 | 0.72 | 2 | 16 | 2 | 0 | 27 | 16 | 0 | 0 |
| 48 | 62.1072 | −6.7055 | 6.85 | 20.4 | 0 | 0.4 | 217 | 54 | 140 | 3 | 1.49 | 0.50 | 0 | 1 | 0 | 26 | 2 | 0 | 0 | 0 |
| 49 | 62.1166 | −6.6505 | – | 10 | 0 | 0 | 276 | 67 | 191 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 |
| 50 | 62.0600 | −6.6982 | 5.48 | 15.7 | 0.1 | 0.3 | 34 | 16 | 106 | 6 | 4.56 | 0.76 | 1 | 8 | 12 | 0 | 12 | 13 | 0 | 1 |
| 51 | 62.1186 | −6.6953 | 4.81 | 12.9 | 0 | 0.5 | 66 | 203 | 243 | 6 | 2.93 | 0.49 | 1 | 2 | 4 | 0 | 35 | 12 | 0 | 1 |
| 52 | 62.1259 | −6.6930 | 6.68 | 11.4 | 0.2 | 0.3 | 399 | 172 | 206 | 2 | 1.63 | 0.82 | 0 | 11 | 0 | 0 | 46 | 0 | 0 | 0 |
| 53 | 62.0260 | −6.8318 | 5.9 | 12.7 | 0 | 1 | 144 | 47 | 316 | 0 | – | – | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 54 | 62.0085 | −6.8273 | 5.34 | 3.4 | 0 | 0.4 | 31 | 35 | 193 | 3 | 2.75 | 0.92 | 0 | 14 | 0 | 0 | 18 | 0 | 0 | 6 |
| 55 | 62.0049 | −6.8238 | 6.72 | 14.8 | 0.2 | 0.4 | 28 | 15 | 181 | 5 | 3.25 | 0.65 | 1 | 12 | 2 | 0 | 9 | 0 | 0 | 1 |
| 56 | 62.0051 | −6.8216 | - | 17 | 0.2 | 0.3 | 144 | 10 | 180 | 2 | 1.29 | 0.65 | 0 | 1 | 0 | 0 | 13 | 0 | 0 | 0 |
| 57 | 62.1753 | −6.7622 | 5.26 | 12.3 | 0 | 0.25 | 66 | 1305 | 116 | 6 | 2.84 | 0.47 | 1 | 19 | 2 | 0 | 77 | 10 | 0 | 6 |
| Variable | F Model | r2 | p-Value |
|---|---|---|---|
| Perimeter | 2.936 | 0.060 | 0.012 |
| Water depth | 2.350 | 0.048 | 0.033 |
| Diversity Index | Adjusted r2 | Variable | b | SE | t-Value | p-Value |
|---|---|---|---|---|---|---|
| Species richness | - | Temperature | −0.013 | 0.015 | −0.838 | 0.402 |
| Water depth | −0.009 | 0.003 | −3.203 | 0.002 | ||
| Shannon diversity | 0.139 | Temperature | −0.018 | 0.011 | −1.617 | 0.111 |
| Water depth | −0.004 | 0.001 | −2.858 | 0.006 | ||
| Evenness | 0.093 | Temperature | −0.008 | 0.006 | −1.348 | 0.185 |
| Water depth | 0.002 | 0.001 | 2.297 | 0.027 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hansen, L.J.; Kreiling, A.-K. Small Islands, Small Ponds, Small Communities—Water Beetles and Water Boatmen in the Faroe Islands. Insects 2022, 13, 923. https://doi.org/10.3390/insects13100923
Hansen LJ, Kreiling A-K. Small Islands, Small Ponds, Small Communities—Water Beetles and Water Boatmen in the Faroe Islands. Insects. 2022; 13(10):923. https://doi.org/10.3390/insects13100923
Chicago/Turabian StyleHansen, Leivur Janus, and Agnes-Katharina Kreiling. 2022. "Small Islands, Small Ponds, Small Communities—Water Beetles and Water Boatmen in the Faroe Islands" Insects 13, no. 10: 923. https://doi.org/10.3390/insects13100923
APA StyleHansen, L. J., & Kreiling, A.-K. (2022). Small Islands, Small Ponds, Small Communities—Water Beetles and Water Boatmen in the Faroe Islands. Insects, 13(10), 923. https://doi.org/10.3390/insects13100923