
Chromosomal and Molecular Diversity in the Simulium ornatum Group (Diptera: Simuliidae) in the Western Tian Shan Range of Central Asia



Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Sampling and Identification
2.2. Chromosomal Conventions and Procedures
2.3. Molecular Procedures
3. Results
3.1. Sampling Sites
3.2. Morphology
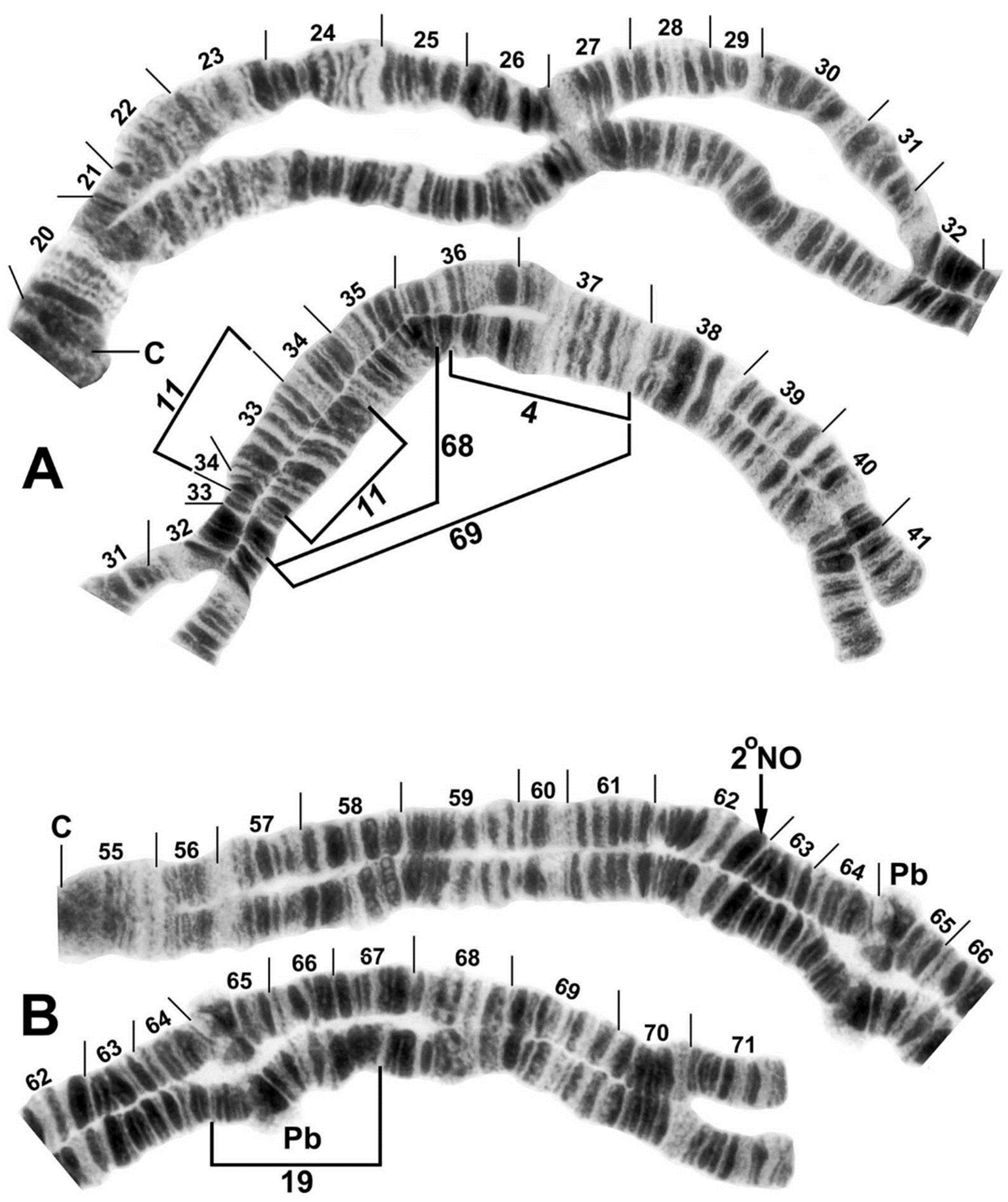
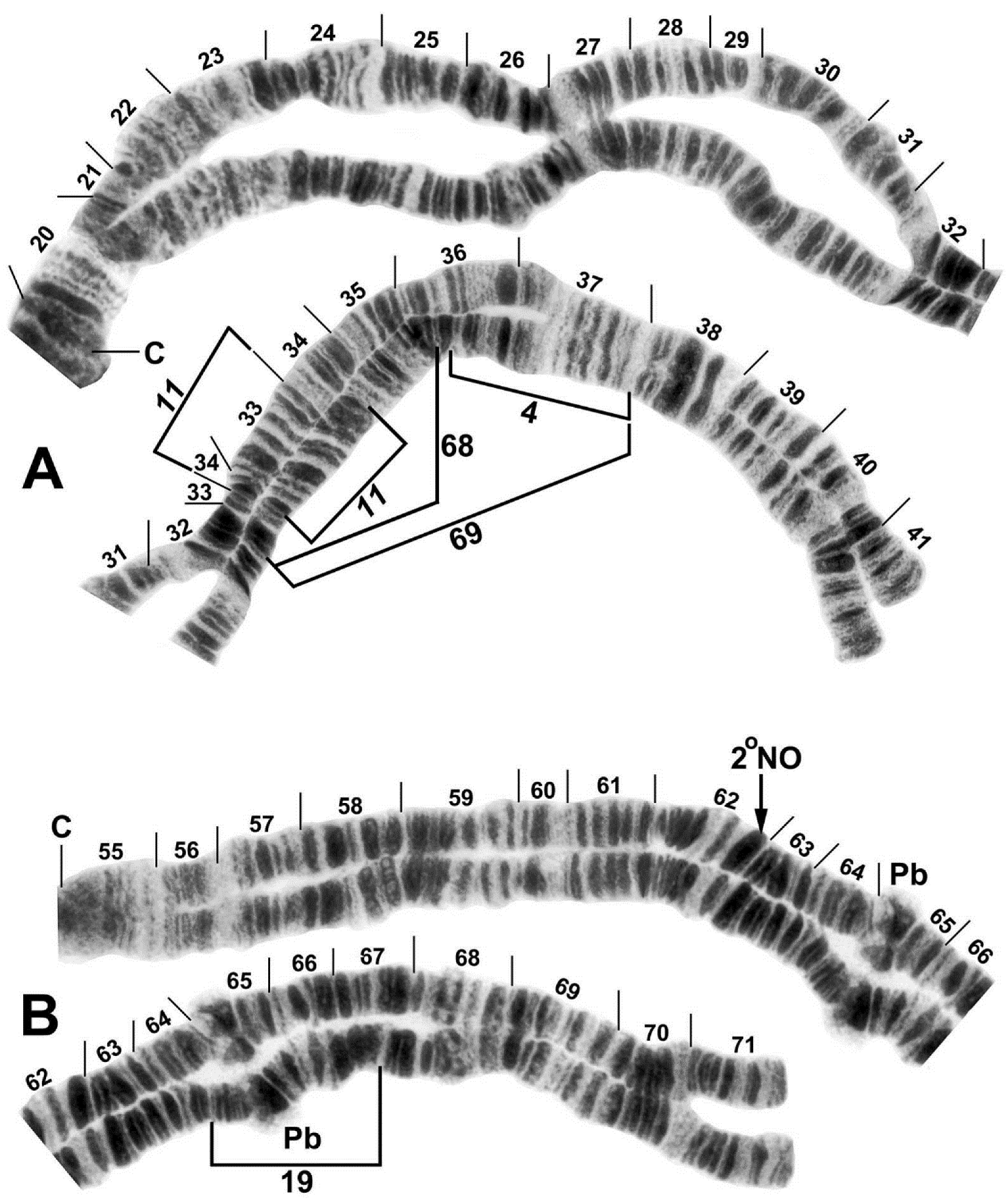
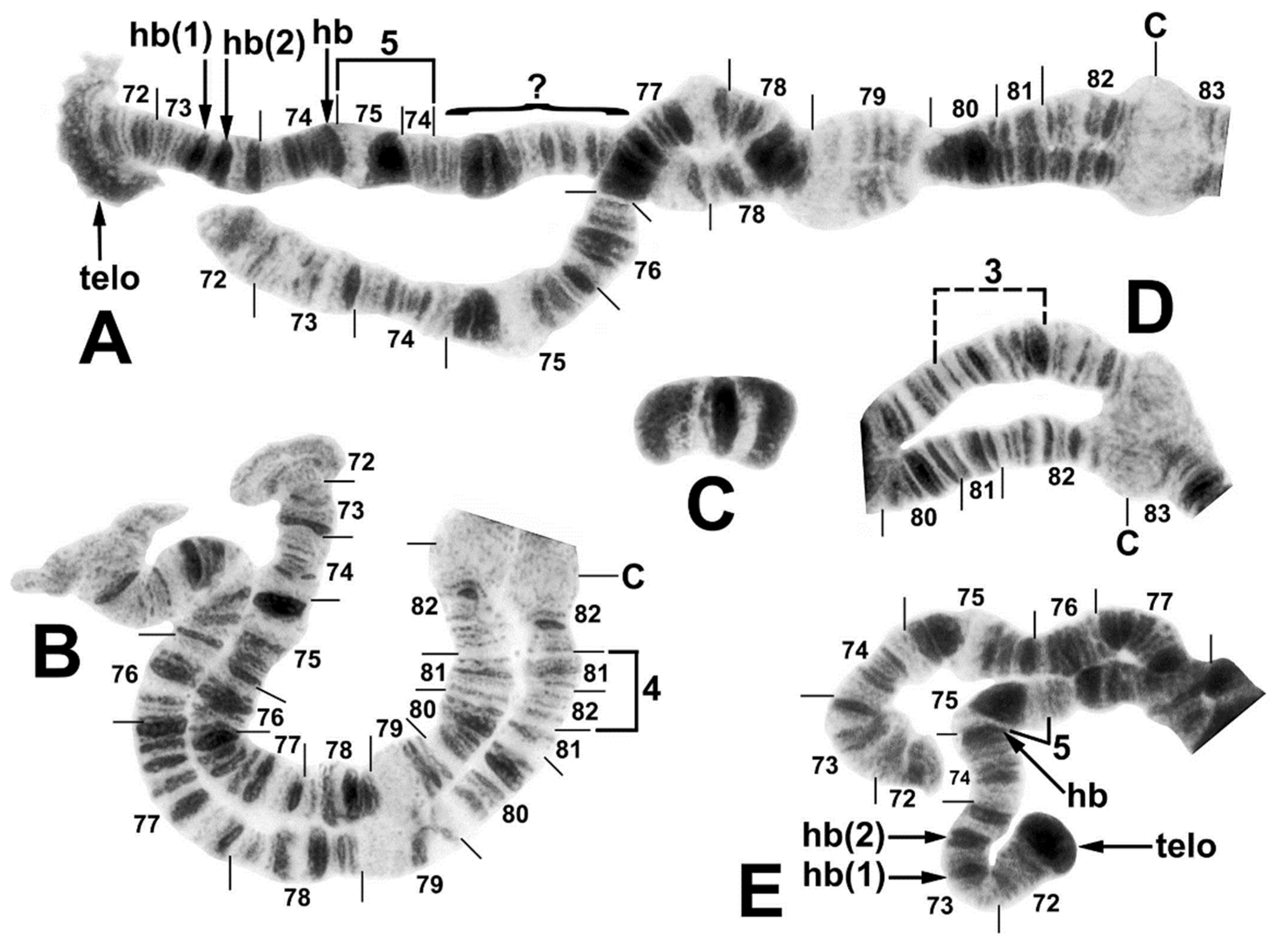
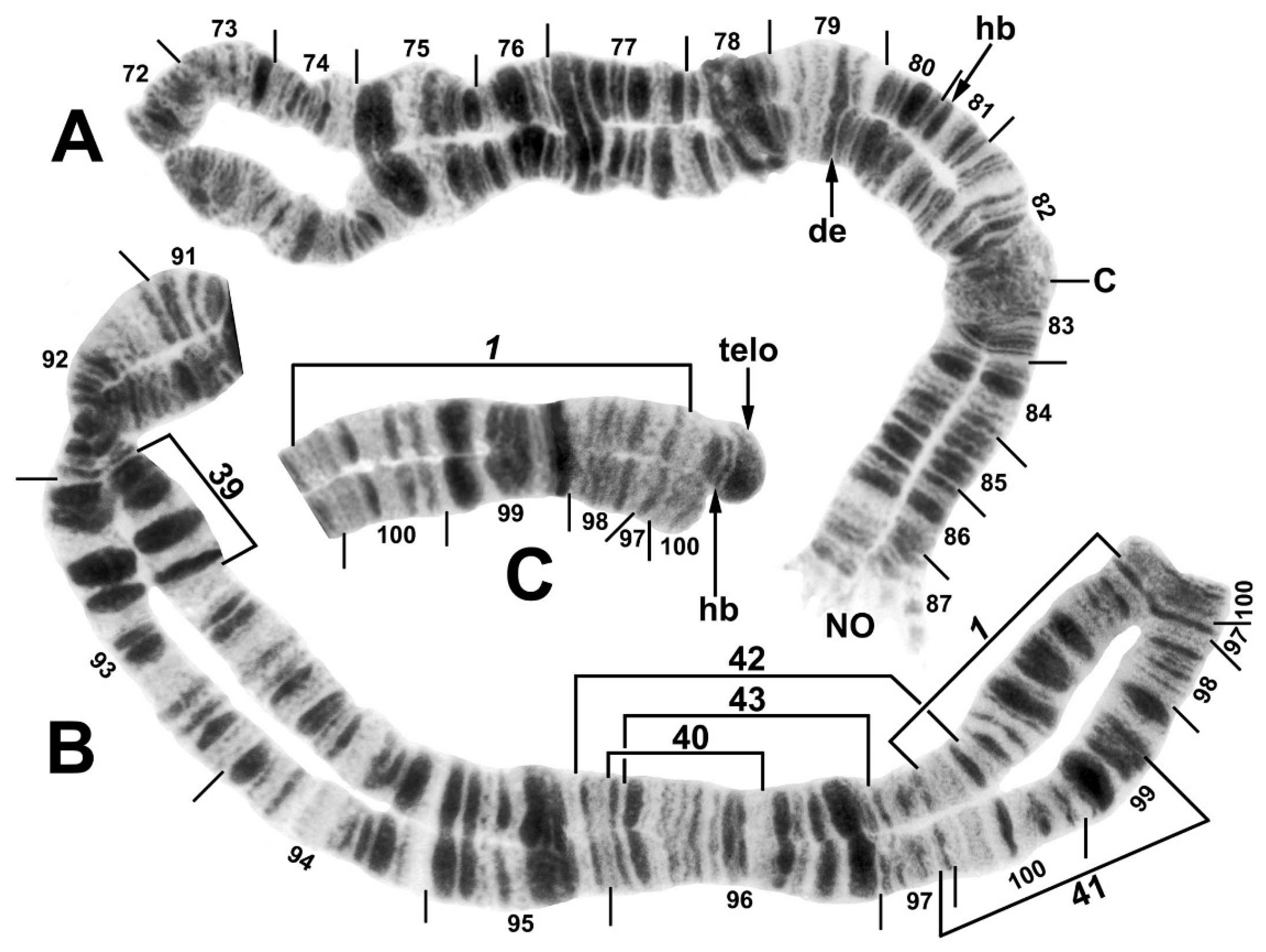
3.3. Chromosomes
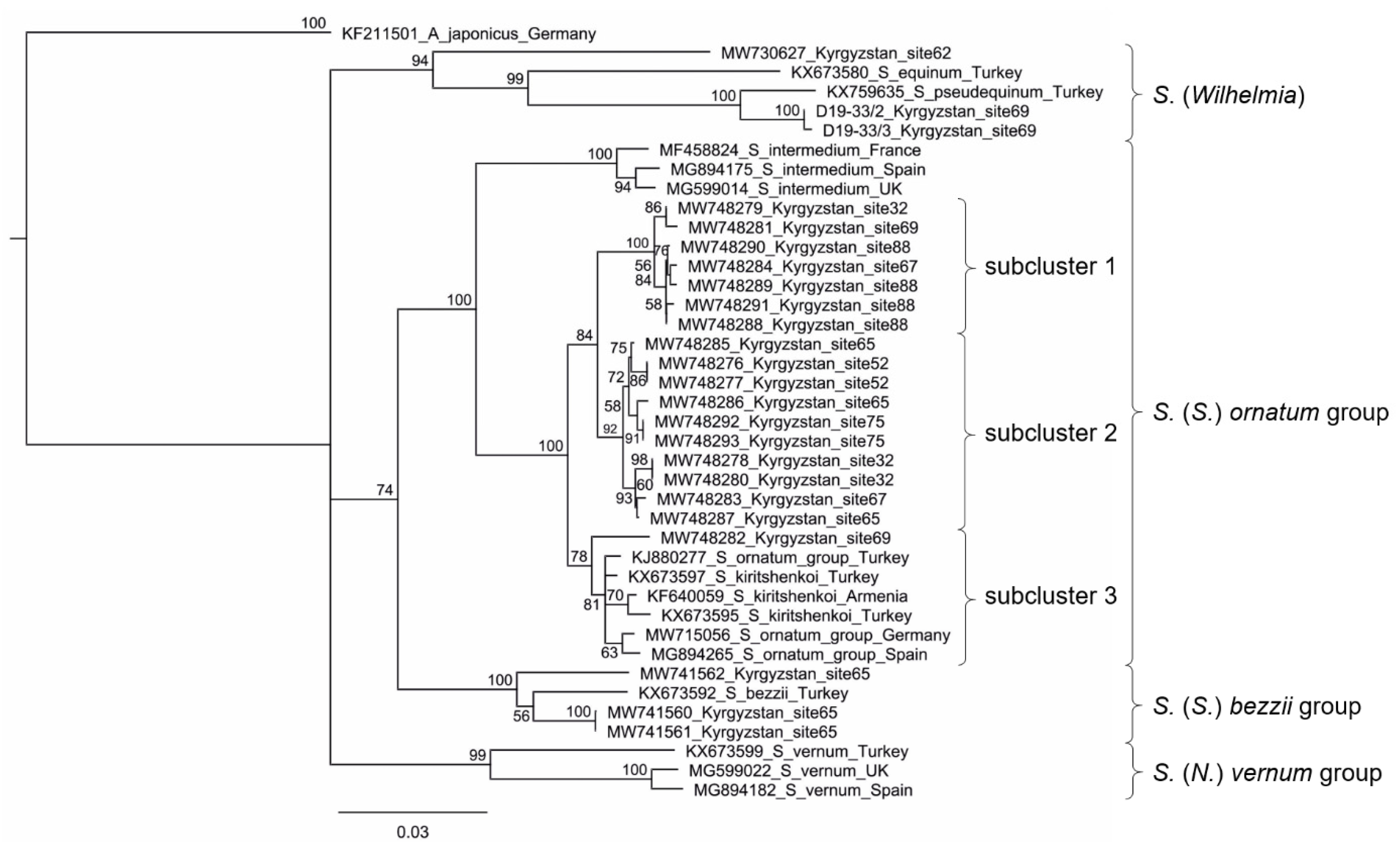
3.4. CO1 Sequences
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adler, P.H. World Blackflies (Diptera: Simuliidae): A Comprehensive Revision of the Taxonomic and Geographical Inventory. 2021. Available online: http://biomia.sites.clemson.edu/pdfs/blackflyinventory.pdf (accessed on 12 June 2021).
- Crosskey, R.W.; Crosskey, M.E. An investigation of the blackfly fauna of Andalusia, southern Spain (Diptera: Simuliidae). J. Nat. Hist. 2000, 34, 895–951. [Google Scholar] [CrossRef]
- Pfenninger, M.; Schwenk, K. Cryptic animal species are homogeneously distributed among taxa and biogeographical regions. BMC Evol. Biol. 2007, 7, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, P.H. Ecology of black fly sibling species. In Black Flies: Ecology, Population Management and Annotated World List; Kim, K.C., Merritt, R.W., Eds.; Pennsylvania State University Press: University Park, PA, USA, 1988; pp. 63–76. [Google Scholar]
- Pramual, P.; Adler, P.H. DNA barcoding of tropical black flies (Diptera: Simuliidae) of Thailand. Mol. Ecol. Res. 2013, 14, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Adler, P.H.; Crosskey, R.W. Cytotaxonomy of the Simuliidae (Diptera): A systematic and bibliographic conspectus. Zootaxa 2015, 3975, 1–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Triana, L.M.; Chaverri, L.G.; Rodríguez-Pérez, M.A.; Prosser, S.W.J.; Hebert, P.D.N.; Gregory, T.R.; Johnson, N. DNA barcoding of Neotropical black flies (Diptera: Simuliidae): Species identification and discovery of cryptic diversity in Mesoamerica. Zootaxa 2015, 3936, 93–114. [Google Scholar] [CrossRef] [Green Version]
- Adler, P.H.; Currie, D.C.; Wood, D.M. The Black Flies (Simuliidae) of North America; Cornell University Press: Ithaca, NY, USA, 2004; xv + 941 pp. + 24 color plates. [Google Scholar]
- Adler, P.H.; Takaoka, H.; Sofian-Azirun, M.; Low, V.L.; Ya’cob, Z.; Chen, C.D.; Lau, K.W.; Pham, X.D. Vietnam, a hotspot for chromosomal diversity and cryptic species in black flies (Diptera: Simuliidae). PLoS ONE 2016, 11, e0163881. [Google Scholar] [CrossRef]
- Tangkawanit, U.; Kuvangkadilok, C.; Baimai, V.; Adler, P.H. Cytosystematics of the Simulium tuberosum group (Diptera: Simuliidae) in Thailand. Zool. J. Linn. Soc. 2009, 155, 289–315. [Google Scholar] [CrossRef] [Green Version]
- Takaoka, H.; Sofian-Azirun, M.; Hashim, R.; Otsuka, Y.; Belabut, D.M.; Ean, T.P. Relationships of black-fly species of the Simulium tuberosum species-group (Diptera: Simuliidae) in Peninsular Malaysia, with keys to ten Malaysian species. Raffles Bull. Zool. 2012, 60, 530–538. [Google Scholar]
- Post, R.J. Cytotaxonomy of the Simulium ornatum species-group in Britain. Newsl. Brit. Simuliid Group 1980, 3, 3–5. [Google Scholar]
- Rubtsov, I.A. Moshki (sem. Simuliidae) [Blackflies (fam. Simuliidae)]. In Fauna of the USSR. New Series No. 64, Insects, Diptera 6 (6); Akademii Nauk SSSR: Moscow, Russia; Petersburg, Russia, 1956; p. 860. (In Russian) English translation: Blackflies (Simuliidae). Fauna of the USSR. Diptera, 2nd ed.; E.J. Brill: Leiden, Netherlands, 1990; p. 1042. [Google Scholar]
- Konurbaev, E.O. Moshki (Diptera: Simuliidae) Srednei Azii; Akademiia Nauk Kirgizckoi SSR, Izdatel’stvo “Ilim”: Frunze [Bishkek], Kyrgyzstan, 1984; p. 232. (In Russian) [Google Scholar]
- Brown, B.V. A further chemical alternative to critical-point-drying for preparing small (or large) flies. Fly Times 1993, 11, 10. [Google Scholar]
- Adler, P.H.; Kúdelová, T.; Kúdela, M.; Seitz, G.; Ignjatović-Ćupina, A. Cryptic biodiversity and the origins of pest status revealed in the macrogenome of Simulium colombaschense (Diptera: Simuliidae), history’s most destructive black fly. PLoS ONE 2016, 11, e0147673. [Google Scholar] [CrossRef]
- Rothfels, K.H.; Feraday, R.; Kaneps, A. A cytological description of sibling species of Simulium venustum and S. verecundum with standard maps for the subgenus Simulium Davies [sic] (Diptera). Can. J. Zool. 1978, 56, 1110–1128. [Google Scholar] [CrossRef]
- Rothfels, K.H. Cytological approaches to black fly taxonomy. In Black Flies: Ecology, Population Management, and Annotated World List; Kim, K.C., Merritt, R.W., Eds.; Pennsylvania State University Press: University Park, PA, USA, 1988; pp. 39–52. [Google Scholar]
- Hébert, P.D.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. London. Ser. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Kampen, H.; Kronefeld, M.; Zielke, D.; Werner, D. Three rarely encountered and one new Culiseta species (Diptera: Culicidae) in Germany. J. Eur. Mosq. Control Assoc. 2013, 31, 36–39. [Google Scholar]
- Onder, A.; Yildirim, A.; Duzlu, O.; Arslan, M.O.; Sari, B.; Tasci, G.T.; Ciloglu, A.; Aydin, N.P.; Inci, A.; Adler, P.H. Molecular characterization of black flies (Diptera: Simuliidae) in areas with pest outbreaks and simuliotoxicosis in Northeast Anatolia Region, Turkey. Acta Trop. 2019, 199, 105149. [Google Scholar] [CrossRef]
- Khanzadeh, F.; Khaghaninia, S.; Maleki-Ravasan, N.; Oshaghi, M.A.; Adler, P.H. Black flies (Diptera: Simuliidae) of the Aras River Basin: Species composition and floral visitation. Acta Trop. 2020, 209, 105536. [Google Scholar] [CrossRef] [PubMed]
- Ilmonen, J.; Adler, P.H.; Malmqvist, B.; Cywinska, A. The Simulium vernum group (Diptera: Simuliidae) in Europe: Multiple character sets for assessing species status. Zool. J. Linn. Soc. 2009, 156, 847–863. [Google Scholar] [CrossRef] [Green Version]
- Pangjanda, S.; Pramual, P. Tests of conspecificity for closely related black flies (Diptera: Simuliidae) species of the Simulium striatum group in Thailand. Zootaxa 2017, 4231, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Pramual, P.; Kuvangkadilok, C. Integrated cytogenetic, ecological, and DNA barcode study reveals cryptic diversity in Simulium (Gomphostilbia) angulistylum (Diptera: Simuliidae). Genome 2012, 55, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Adler, P.H.; Srisuka, W.; Low, V.L.; Takaoka, H.; Saeung, A. High-elevation chromosomal diversity of black flies (Diptera: Simuliidae) in Thailand. Insect Syst. Divers. 2019, 3, 1–10. [Google Scholar] [CrossRef]
- Shelley, A.J.; Arzube, M.; Couch, C.A. The Simuliidae (Diptera) of the Santiago onchocerciasis focus of Ecuador. Bull. Brit. Mus. Nat. Hist. (Entomol.) 1989, 58, 79–130. [Google Scholar]
- Wilson, M.D.; Post, R.J.; Boakye, D.A. Studies on environmentally-induced colour variation in Simulium sirbanum (Diptera: Simuliidae) using a portable rearing system. Ann. Trop. Med. Parasitol. 1992, 86, 169–174. [Google Scholar] [CrossRef]
- Moulton, J.K. Reexamination of Simulium (Psilopelmia) Enderlein (Diptera: Simuliidae) of America north of Mexico. Proc. Entomol. Soc. Wash. 1998, 100, 50–71. [Google Scholar]
- Konurbaev, E.O. Variability of some of the quantitative characters of simuliid larvae (Diptera, Simuliidae) in the mountains of Soviet Central Asia. Entomol. Oboz. 1973, 52, 915–923, (In Russian; English translation in Entomol. Rev. 1973, 52, 590–595). [Google Scholar]
- Zhang, Y.; Malmqvist, B. Phenotypic plasticity in a suspension-feeding insect, Simulium lundstromi (Diptera: Simuliidae), in response to current velocity. Oikos 1997, 78, 503–510. [Google Scholar] [CrossRef]
- Palmer, R.W.; Craig, D.A. An ecological classification of primary labral fans of filter-feeding black fly (Diptera: Simuliidae) larvae. Can. J. Zool. 2000, 78, 199–218. [Google Scholar] [CrossRef]
- Zettler, J.A.; Adler, P.H.; McCreadie, J.W. Factors influencing larval color in the Simulium vittatum complex (Diptera: Simuliidae). Invert. Biol. 1998, 117, 245–252. [Google Scholar] [CrossRef]
- Adler, P.H.; Kachvorian, E.A. Cytogenetics of the Holarctic black fly Simulium noelleri (Diptera: Simuliidae). Can. J. Zool. 2001, 79, 1972–1979. [Google Scholar] [CrossRef]
- Chubareva, L.A.; Petrova, N.A. Tsitologicheskie Karty Politennykh Khromosom i Nekotorye Morfologicheskie Osobennosti Krovososushchikh Moshek Rossii i Sopredel’nykh Stran (Diptera: Simuliidae): Atlas [Cytological Maps of Polytene Chromosomes and Some Morphological Features of Bloodsucking Black Flies of Russia and Adjacent Countries (Diptera: Simuliidae): Atlas]; Tovarishchestvo Nauchnykh Izdanii KMK: St. Petersburg, Russia, 2008; Available online: https://scholar.google.com/scholar?q=+Chubareva+LA+Petrova+NA++(+2008+)++Tsitologicheskie+karty+politennykh+khromosom+i+nekotorye+morfologicheskie+osobennosti+krovososushchikh+moshek+Rossii+i+sopredel’nykh+stran+(+Diptera+:++Simuliidae+):+atlas++[+Cytological+maps+of+polytene+chromosomes+and+some+morphological+features+of+bloodsucking+black+flies+of+Russia+and+adjacent+countries+(+Diptera+:++Simuliidae+):+atlas+].++Tovarishchestvo+Nauchnykh+Izdanii+KMK+,++St.+Petersburg++[In+Russian]+ (accessed on 9 September 2021).
- Landau, R. Four forms of Simulium tuberosum (Lundstr.) in southern Ontario: A salivary gland chromosome study. Can. J. Zool. 1962, 40, 921–939. [Google Scholar] [CrossRef]
- Rothfels, K.H.; Freeman, D.M. The salivary gland chromosomes of seven species of Prosimulium (Diptera, Simuliidae) in the mixtum (IIIL-1) group. Can. J. Zool. 1977, 55, 482–507. [Google Scholar] [CrossRef]
- Shields, G.F.; Procunier, W.S. A cytological description of sibling species of Simulium (Gnus) arcticum (Diptera: Simuliidae). Polar Biol. 1982, 1, 181–192. [Google Scholar] [CrossRef]
- Procunier, W.S. The interdependence of B chromosomes, nucleolar organizer expression, and larval development in the blackfly species Cnephia dacotensis and Cnephia ornithophilia (Diptera: Simuliidae). Can. J. Zool. 1982, 60, 2879–2896. [Google Scholar] [CrossRef]
- Jones, N.; Ruban, A. Are B chromosomes useful for crop improvement? Plants People Planet 2019, 1, 84–92. [Google Scholar] [CrossRef]
- Meier, R.; Shiyang, K.; Vaidya, G.; Ng, P.K.L. DNA barcoding and taxonomy in Diptera: A tale of high intraspecific variability and low identification success. Syst. Biol. 2006, 55, 715–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, P.H.; Cheke, R.A.; Post, R.J. Evolution, epidemiology, and population genetics of black flies (Diptera: Simuliidae). Infect. Genet. Evol. 2010, 10, 846–865. [Google Scholar] [CrossRef] [PubMed]
- Rubtsov, I.A. Blackflies (fam. Simuliidae). In Fauna of the USSR. New Series No. 23, Insects, Diptera 6 (6); Akademii Nauk SSSR: Moscow, Russia; Petersburg, Russia, 1940; ix + 533 pp. (In Russian, English keys and descriptions of new species). [Google Scholar]
- Rubtsov, I.A. The composition, relationships, geographical distribution and probable origin of the black fly fauna of Central Asia. Izv. Akad. Nauk SSSR Ser. Biol. 1947, 1, 87–116, (In Russian, summary and species descriptions in English). [Google Scholar]
- Yankovsky, A.V. Family Simuliidae Newman. In Katalog Tipovykh Ekzemplyarov Kollektsii Zoologicheckogo Instituta RAN, Diptera. 5. Simuliidae, Culicidae [Catalogue of Type Specimens in the Collection of the Zoological Institute, Russian Academy of Sciences, Diptera. 5. Simuliidae, Culicidae]; Yankovsky, A.V., Ulyanov, K.N., Eds.; Rossiiskaya Akademiya Nauk, Zoologicheskii Institut: St. Petersburg, Russia, 1995; pp. 1–61. (In Russian) [Google Scholar]
- Siviglia, A.; Repetto, R.; Zolezzi, G.; Tubino, M. River bed evolution due to channel expansion: General behaviour and application to a case study (Kugart River, Kyrgyz Republic). River Res. Applic. 2008, 24, 1271–1287. [Google Scholar] [CrossRef]
- Adler, P.H.; Malmqvist, B.; Zhang, Y. Black flies (Diptera: Simuliidae) of northern Sweden: Taxonomy, chromosomes, and bionomics. Entomol. Scand. 1999, 29, 361–382. [Google Scholar] [CrossRef]
- Kachvorian, E.A.; Adler, P.H. Black flies (Diptera, Simuliidae) of Armenia. Biol. J. Armen. 1999, 52, 299–301. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Location 1 (Stream Width) | Latitude and Longitude | Elevation (m Above Sea Level) | Date | Larvae Collected: Chromosome Preparations 2 | CO1 Sequences 3 |
|---|---|---|---|---|---|---|
| 5 | Issyk-Kul region, roadside stream (1 m) | 42°26′48″ N 75°50′35″ E | 1757 | 18 June | 32:23 | - |
| 18 | Naryn region, rocky roadside stream (1.5 m) | 41°31′24″ N 75°02′10″ E | 1912 | 21 June | 10:0 | - |
| 20 | Naryn region, meadow stream (1 m) | 41°12′32″ N 74°45′42″ E | 2080 | 22 June | 20:4 | - |
| 21 | Naryn region, polluted roadside stream (0.75 m) | 41°13′54″ N 74°41′52″ E | 1995 | 22 June | 8:0 | - |
| 32 | Naryn region, muddy roadside stream (2 m) | 41°21′52″ N 73°44′40″ E | 1827 | 23 June | 55:11 | 3 |
| 34 | Naryn region, roadside trickle (0.3 m) | 41°19′17″ N 73°39′48″ E | 2052 | 23 June | 45:22 | - |
| 46 | Jalal-Abad region, grassy mountainside stream (0.3 m) | 40°33′05″ N 73°58′26″ E | 1805 | 26 June | 11:4 | - |
| 51 | Osh region, Hwy. M41, 3 km W of Sary-Tash (3 m) | 39°44′38″ N 73°12′25″ E | 3270 | 28 June | 1:1 | - |
| 52 | Osh region, Hwy. A371, Tajikistan border, Kara-Kindik River (3–4 m) | 39°42′00” N 73°27′16” E | 3246 | 28 June | 6:6 | 2 |
| 60 | Osh region, Hwy. M41, Mashrapsay River (4–8 m) | 40°17′09″ N 73°16′11″ E | 2072 | 29 June | 1:1 | - |
| 62 | Osh region, wide valley stream (1 m) | 40°16′01″ N 73°03′20″ E | 1736 | 29 June | 23:10 | - |
| 65 | Jalal-Abad region, 0.25 km N of site 67 (1 m) | 41°17′39″ N 72°40′39″ E | 1762 | 1 July | 34:23 | 3 |
| 67 | Jalal-Abad region, ca. 3 km N of village of Alash (3 m) | 41°17′32″ N 72°40′34″ E | 1743 | 1 July | 13:10 | 2 |
| 69 | Jalal-Abad region, springfed stream with strong flow ca. 6 km NE of Maylisuu (0.15 m) | 41°18′31″ N 72°29′51″ E | 1098 | 2 July | 21:7 | 2 |
| 75 | Talas region, tumbling mountain stream ca. 8 km W of Toluk (1–3 m) | 41°55′32″ N 73°28′45″ E | 1870 | 3 July | 8:5 | 2 |
| 78 | Naryn region, roadside meadow trickle (0.15 m) | 41°52′46″ N 73°41′43″ E | 2348 | 4 July | 10:5 | - |
| 85 | Naryn region, Kekemeren River valley, tumbling stream ca. 2 km S of Kyzyl-Oi (10–15 m) | 41°55′52″ N 74°09′11″ E | 1737 | 5 July | 2:2 | - |
| 88 | Naryn region, roadside stream, ca. 5 km NW of Dzholkara (1.5 m) | 41°53′15″ N 74°39′46″ E | 2185 | 5 July | 36:18 | 4 |
| 89 | Naryn region, roadside stream ca. 1 km S of Bashkaingdy (15–20 m) | 41°59′13″ N 74°39′41″ E | 1804 | 5 July | 4:4 | - |
| 95 | Naryn region, Hwy. A367, roadside stream, ca. 6 km W of Uzunbulak (0.5 m) | 42°06′07″ N 75°11′02″ E | 2512 | 6 July | 1:1 | - |
| Site | 5 | 20 | 32 | 34 | 46 | 51 | 52 | 60 | 62 | 65 | 67 | 69 | 75 | 78 | 85 | 88 | 89 | 95 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Females: Males | 10:12 | 3 1:1 | 5:2 | 12:10 | 3 2:0 | 1:0 | 5:1 | 1:0 | 4 1:2 | 12:10 | 7 1:3 | 0:2 | 1:3 | 2:3 | 1:1 | 9:8 | 2:2 | 0:1 2 |
| IL-4 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.35 | - | 0.50 |
| IL-11 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| IL-68 | - | - | - | 0.02 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| IL-69 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.03 | - | - |
| IIL-19 | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.10 | - | - | - | - |
| IIL 2°NO 3 | - | - | - | - | - | - | - | - | - | 0.02 | - | - | - | - | - | - | - | - |
| IIIS-3 | - | - | - | * 4 | - | - | - | - | - | * 4 | * 4 | * 4 | * 4 | - | - | - | - | - |
| IIIS-4 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.25 | - | - | - |
| IIIS-5 5 | 0.02 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.03 | - | - |
| IIIS hb81 | - | - | ** 6 | ** 6 | - | - | - | - | ** 6 | ** 6 | ** 6 | - | - | - | - | - | - | - |
| IIIS de79 | - | - | - | - | - | - | - | 0.50 | - | - | - | - | - | - | - | - | - | - |
| IIIS hb73(1) 5 | 0.02 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.03 | - | - |
| IIIS hb73(2) 5 | 0.02 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.03 | - | - |
| IIIS hb74 5 | 0.02 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.03 | - | - |
| IIIS hb telo 5 | 0.02 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.12 | - | - |
| IIIL-1 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| IIIL-39 | - | - | 0.07 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| IIIL-40 | - | - | - | 0.02 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| IIIL-41 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.09 | - | - |
| IIIL-42 | 0.02 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| IIIL-43 7 | 0.02 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| IIIL hb100 + telo 8 | 0.11 | - | - | - | - | - | - | - | 0.08 | 0.02 | 0.10 | - | - | - | - | - | - | - |
| Bs (%) | - | - | - | - | - | - | 0.33 | - | - | - | - | - | - | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adler, P.H.; Werner, D.; Kampen, H. Chromosomal and Molecular Diversity in the Simulium ornatum Group (Diptera: Simuliidae) in the Western Tian Shan Range of Central Asia. Insects 2021, 12, 817. https://doi.org/10.3390/insects12090817
Adler PH, Werner D, Kampen H. Chromosomal and Molecular Diversity in the Simulium ornatum Group (Diptera: Simuliidae) in the Western Tian Shan Range of Central Asia. Insects. 2021; 12(9):817. https://doi.org/10.3390/insects12090817
Chicago/Turabian StyleAdler, Peter H., Doreen Werner, and Helge Kampen. 2021. "Chromosomal and Molecular Diversity in the Simulium ornatum Group (Diptera: Simuliidae) in the Western Tian Shan Range of Central Asia" Insects 12, no. 9: 817. https://doi.org/10.3390/insects12090817
APA StyleAdler, P. H., Werner, D., & Kampen, H. (2021). Chromosomal and Molecular Diversity in the Simulium ornatum Group (Diptera: Simuliidae) in the Western Tian Shan Range of Central Asia. Insects, 12(9), 817. https://doi.org/10.3390/insects12090817