
Parasitism and Suitability of Aprostocetus brevipedicellus on Chinese Oak Silkworm, Antheraea pernyi, a Dominant Factitious Host

 ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Parasitoids
2.2. Host
2.3. Antheraea pernyi Egg Treatments
2.4. Suitability of A. pernyi Egg with Different Treatments on Parasitism by A. brevipedicellus
2.4.1. No-Choice Test
2.4.2. Choice Test
2.5. Statistical Analysis
3. Results
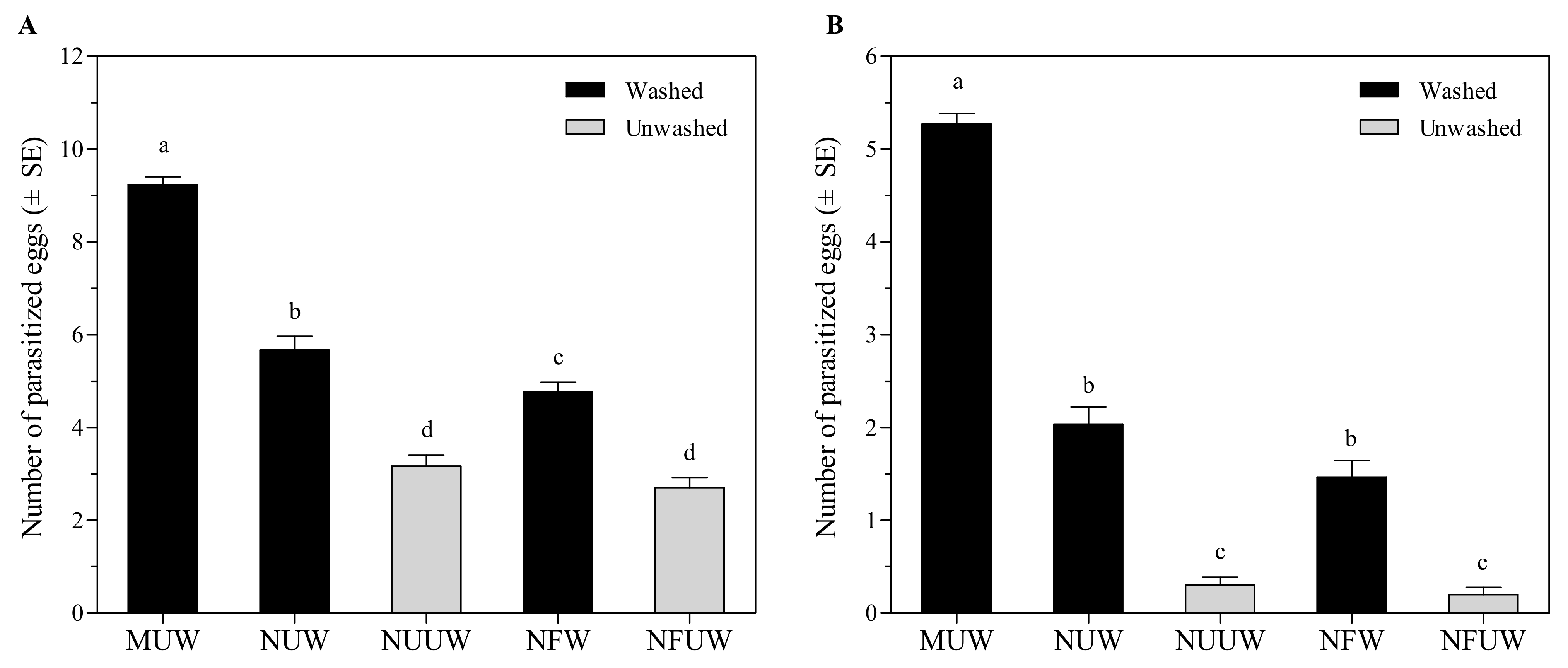
3.1. Suitability of A. pernyi Egg with Different Treatments on Parasitism by A. brevipedicellus
3.1.1. No-Choice Test
3.1.2. Choice Test
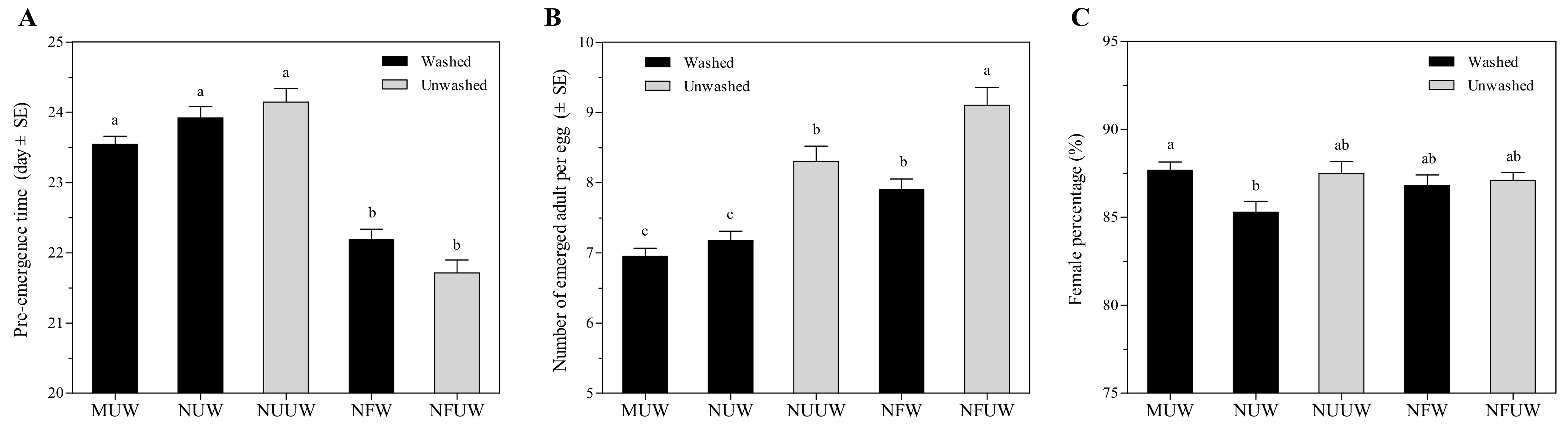
3.2. Suitability of A. pernyi Egg with Different Treatments on Development of A. brevipedicellus Offspring
3.2.1. Pre-Emergence Time
3.2.2. Emergence of Aprostocetus brevipedicellus
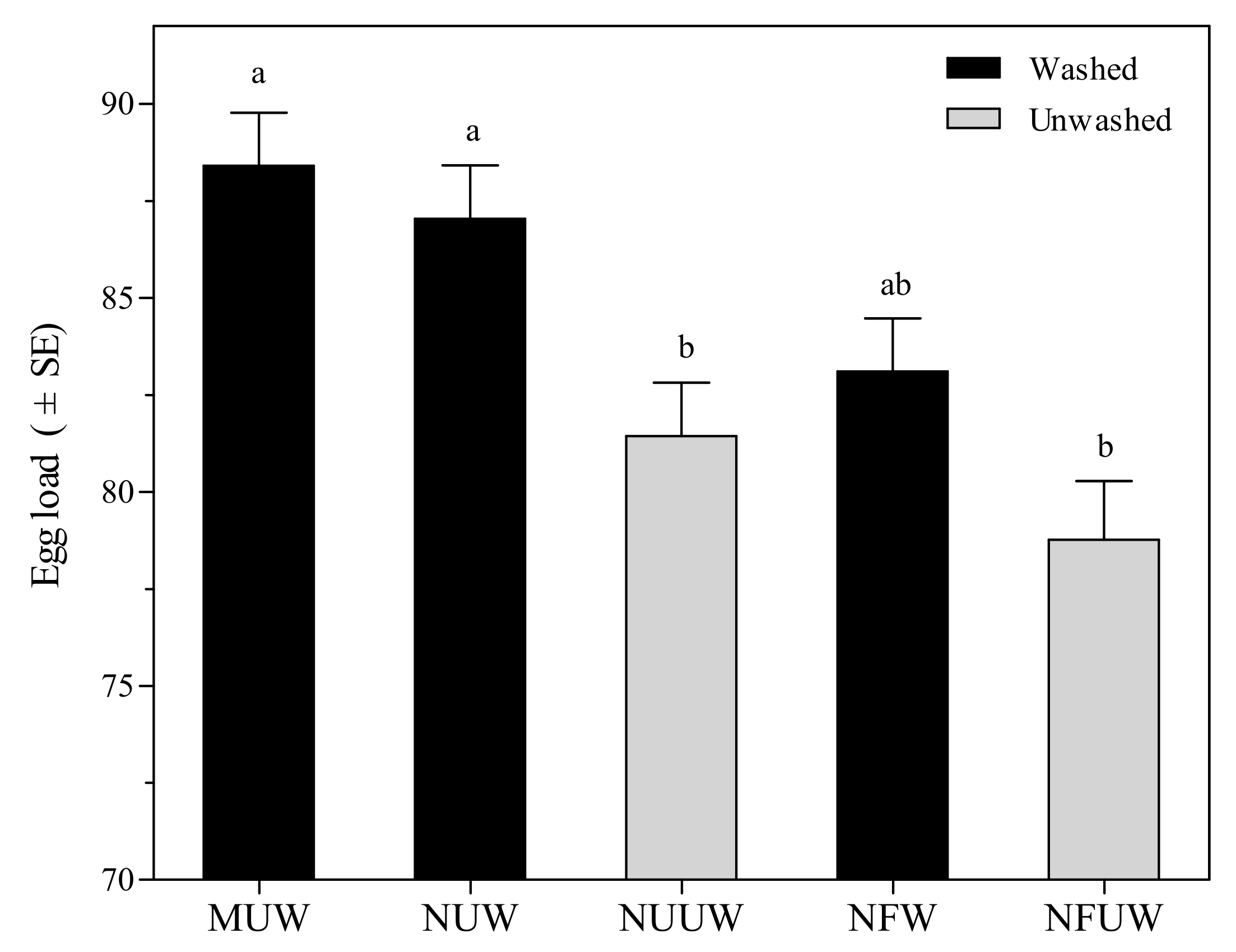
3.2.3. Female Hind Tibia Length and Egg Load of Aprostocetus brevipedicellus Offspring
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Song, Y.S.; Dong, Y.Q.; Cui, D.Y.; Yue, F.Z.; Zhu, N.B.; Bai, H.Y. Species analysis of forest pest in China, V. Insect. Forest Pest. Dis. 2019, 38, 26–30. (In Chinese) [Google Scholar]
- Yang, Z.-Q.; Wang, X.-B.Y.; Zhang, Y.-N. Recent advances in biological control of important native and invasive forest pests in China. Biol. Control 2014, 68, 117–128. [Google Scholar] [CrossRef]
- Xu, M.; Xu, F.Y.; Liu, Y.P.; Pan, Y.S.; Wu, X.Q. Assessment of Metarhizium anisopliae (Clavicipitaceae) and its vector, Scleroderma guani (Hymenoptera: Bethylidae), for the control of Monochamus alternatus (Coleoptera: Cerambycidae). Can. Entomol. 2015, 147, 628–634. [Google Scholar] [CrossRef]
- Li, D.S.; Liao, C.Y.; Zhang, B.X.; Song, Z.W. Biological control of insect pests in litchi orchards in China. Biol. Control. 2014, 68, 23–36. [Google Scholar] [CrossRef]
- Chen, Y.M.; Gibson, G.A.P.; Peng, L.F.; Iqbal, A.; Zang, L.S. Anastatus Motschulsky (Hymenoptera, Eupelmidae): Egg parasitoids of Caligula japonica Moore (Lepidoptera, Saturniidae) in China. ZooKeys 2019, 881, 109–134. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Desneux, N.; Wajnberg, E.; Fauvergue, X.; Privet, S.; Kaiser, L. Oviposition behaviour and patch-time allocation in two aphid parasitoids exposed to deltamethrin residues. Entomol. Exp. Appl. 2004, 112, 227–235. [Google Scholar] [CrossRef]
- Desneux, N.; Ramirez-Romero, R.; Kaiser, L. Multistep bioassay to predict recolonization potential of emerging parasitoids after a pesticide treatment. Environ. Toxicol. Chem. 2006, 25, 2675–2682. [Google Scholar] [CrossRef] [Green Version]
- Taning, C.N.T.; Vanommeslaeghe, A.; Smagghe, G. With or without foraging for food, field-realistic concentrations of sulfoxaflor are equally toxic to bumblebees (Bombus terrestris). Entomol. Gen. 2019, 39, 151–155. [Google Scholar] [CrossRef]
- Varikou, K.; Garantonakis, N.; Birouraki, A. Exposure of Bombus terrestris L. to three different active ingredients and two application methods for olive pest control. Entomol. Gen. 2019, 39, 53–60. [Google Scholar] [CrossRef]
- Desneux, N.; Wajnberg, E.; Wyckhuys, K.A.G.; Burgio, G.; Arpaia, S.; Narváez-Vasquez, C.A.; González-Cabrera, J.; Ruescas, D.C.; Tabone, E.; Frandon, J.; et al. Biological invasion of European tomato crops by Tuta absoluta: Ecology, geographic expansion and prospects for biological control. J. Pest. Sci. 2010, 83, 197–215. [Google Scholar] [CrossRef]
- Huang, N.X.; Jaworski, C.C.; Desneux, N.; Zhang, F.; Yang, P.Y.; Wang, S. Long-term and large-scale releases of Trichogramma promote pesticide decrease in maize in northeastern China. Entomol. Gen. 2020, 40, 331–335. [Google Scholar] [CrossRef]
- Zang, L.S.; Wang, S.; Zhang, F.; Desneux, N. Biological control with Trichogramma in China: History, present status and perspectives. Annu. Rev. Entomol. 2021, 66, 463–484. [Google Scholar] [CrossRef]
- Chan, M.S.; Godfray, H.C.J. Host-feeding strategies of parasitoid wasps. Evol. Ecol. 1993, 7, 593–604. [Google Scholar] [CrossRef]
- Pennacchio, F.; Strand, M.R. Evolution of developmental strategies in parasitic Hymenoptera. Annu. Rev. Entomol. 2006, 49, 233–258. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.J.; Bauer, L.S.; Abell, K.J.; Lelito, J.P.; Van Driesche, R. Establishment and abundance of Tetrastichus planipennisi (Hymenoptera: Eulophidae) in Michigan: Potential for success in classical biocontrol of the invasive emerald ash borer (Coleoptera: Buprestidae). J. Econ. Entomol. 2013, 106, 1145–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.H.; Che, P.F.; Yang, X.B.; Song, L.W.; Zhang, C.R.; Benelli, G.; Desneux, N.; Zang, L.S. Optimized pupal age of Tenebrio molitor L. (Coleoptera: Tenebrionidae) enhanced mass rearing efficiency of Chouioia cunea Yang (Hymenoptera: Eulophidae). Sci. Rep. 2019, 9, 3229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morais, W.C.C.; Plata-Rueda, A.; Martínez, L.C.; Zanuncio, A.J.V.; Fernandes, F.L.; Wilcken, C.F.; Zanuncio, J.C.; Serrão, J.E. Potential of Diaphania hyalinata and Tenebrio molitor as alternative host for mass rearing of Palmistichus elaeisis (Hymenoptera: Eulophidae). Entomol. Gen. 2019, 39, 285–294. [Google Scholar] [CrossRef]
- Voegele, J.M. Biological control of Brontispa longissima in Western Samoa: An ecological and economic evaluation. Agric. Ecosyst. Environ. 1989, 27, 315–329. [Google Scholar] [CrossRef]
- Yao, Y.X. Taxonomic Study on Chalcidoids Parasitizing Forest Defoliators in China (Hymenoptera). Ph.D. Thesis, Chinese Academy of Forestry Sciences, Beijing, China, 2005. (In Chinese). [Google Scholar]
- Yang, Z.Q.; Yao, Y.X.; Cao, L.M. Chalcidoidea Parasitizing Forest Defoliators (Hymenoptera); Science Press: Beijing, China, 2015; p. 283. (In Chinese) [Google Scholar]
- Jacas, J.A.; Peña, J.E.; Duncan, R.E. Successful oviposition and reproductive biology of Aprostocetus vaquitarum (Hymenoptera: Eulophidae): A predator of Diaprepes abbreviatus (Coleoptera: Curculionidae). Biol. Control 2005, 33, 352–359. [Google Scholar] [CrossRef]
- Ulmer, B.J.; Jacas, J.A.; Peña, J.E.; Duncan, R.E.; Castillo, J. Effect of temperature on life history of Aprostocetus vaquitarum (Hymenoptera: Eulophidae), an egg parasitoid of Diaprepes abbreviatus (Coleoptera: Curculionidae). Biol. Control 2006, 39, 19–25. [Google Scholar] [CrossRef]
- Li, J.Q.; Yang, Y.; Wang, S.X.; Feng, H.C.; Huang, D.Z.; Jin, Y.J. Host selection and location behavior of Aprostocetus prolixus LaSalle et Huang (Hymenoptera: Eulophidae), an egg parasitoid of Apriona germari (Hope) (Coleoptera: Cerambycidae). Acta Entomol. Sin. 2007, 50, 1122–1128. (In Chinese) [Google Scholar]
- Nacro, S.; Nénon, J.P. Female reproductive biology of Platygaster diplosisae (Hymenoptera: Platygastridae) and Aprostocetus procerae (Hymenoptera: Eulophidae), two parasitoids associated with the African rice gall midge, Orseolia oryzivora (Diptera: Cecidomyiidae). Entomol. Sci. 2008, 11, 231–237. [Google Scholar] [CrossRef]
- Ouattara, D.; Nacro, S.; Latévi, K.; Coulibaly, A. Ecology of Platygaster Diplosisae (Hymenoptera: Platygasteridae) and Aprostocetus Procerae (Hymenoptera: Eulophidae), parasitoids of Orseolia Oryzivora (Diptera: Cecidomyiidae). Int. J. Curr. Adv. Res. 2019, 8, 20482–20487. [Google Scholar]
- Sampson, B.J.; Roubos, C.R.; Stringer, S.J.; Marshall, D.; Liburd, O.E. Biology and efficacy of Aprostocetus (Eulophidae: Hymenoptera) as a parasitoid of the blueberry gall midge complex: Dasineura oxycoccana and Prodiplosis vaccinii (Diptera: Cecidomyiidae). J. Econ. Entomol. 2013, 106, 73–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tee, H.S.; Saad, A.R.; Lee, C.Y. Suitability of heat- and freeze-killed oothecae of the American Cockroach (Dictyoptera: Blattidae) as hosts for an Oothecal parasitoid, Aprostocetus hagenowii (Hymenoptera: Eulophidae). J. Econ. Entomol. 2010, 103, 1770–1774. [Google Scholar] [CrossRef]
- Liu, P.C.; Men, J.; Zhao, B.; Wei, J.R. Fitness-related offspring sex allocation of Anastatus disparis, a gypsy moth egg parasitoid, on different-sized host species. Entomol. Exp. Appl. 2017, 163, 281–286. [Google Scholar] [CrossRef]
- Li, T.H.; Tian, C.Y.; Zang, L.S.; Hou, Y.Y.; Ruan, C.C.; Yang, X.B.; Lucie, M.; Desneux, N. Multiparasitism with Trichogramma dendrolimi on egg of Chinese oak silkworm, Antheraea pernyi, enhances emergence of Trichogramma ostriniae. J. Pest. Sci. 2019, 92, 707–713. [Google Scholar] [CrossRef]
- Li, X.-B.Y.; Lei, Q.; Hua, H.Q.; Song, H.F.; Wang, S.; Ramirez-Romero, R.; Dai, H.J.; Li, J.T.; Li, Y.X. Impact of host suitability on oviposition preference toward fertilized and unfertilized host eggs in two Trichogramma parasitoid species. Entomol. Gen. 2019, 39, 313–323. [Google Scholar] [CrossRef]
- Iqbal, A.; Chen, Y.M.; Hou, Y.Y.; Zhang, L.S.; Desneux, N.; Zang, L.S. Factitious host species impact on the outcome of multiparasitism between egg parasitoids. J. Pest. Sci. 2019, 92, 1261–1269. [Google Scholar] [CrossRef]
- Wang, Y.; Zou, Z.P.; Hou, Y.Y.; Yang, X.B.; Wang, S.; Dai, H.J.; Xu, Y.Y.; Zang, L.S. Manually-extracted unfertilized eggs of Chinese oak silkworm, Antheraea pernyi, enhance mass production of Trichogramma parasitoids. Entomol. Gen. 2020, 40, 397–406. [Google Scholar] [CrossRef]
- Krugner, R. Suitability of non-fertilized eggs of Homalodisca vitripennis for the egg parasitoid Gonatocerus morrilli. BioControl 2014, 59, 167–174. [Google Scholar] [CrossRef]
- Xu, J.; Yang, X.B.; Lin, Y.; Zang, L.S.; Tian, C.Y.; Ruan, C.C. Effect of fertilized, unfertilized, and UV-irradiated hosts on parasitism and suitability for Trichogramma parasitoids. Entomol. Exp. Appl. 2016, 161, 50–56. [Google Scholar] [CrossRef]
- Yang, X.; Qu, Y.L.; Wu, Z.Y.; Lin, Y.; Ruan, C.C.; Desneux, N.; Zang, L.S. Parasitism and suitability of fertilized and nonfertilized eggs of the rice striped stem borer, Chilo suppressalis (Lepidoptera: Crambidae), for Trichogramma parasitoids. J. Econ. Entomol. 2016, 109, 1524–1528. [Google Scholar] [CrossRef]
- Du, W.M.; Xu, J.; Hou, Y.Y.; Lin, Y.; Zang, L.S.; Yang, X.B.; Zhang, J.J.; Ruan, C.C.; Desneux, N. Trichogramma parasitoids can distinguish between fertilized and unfertilized host eggs. J. Pest. Sci. 2018, 91, 771–780. [Google Scholar] [CrossRef]
- Yang, S.Y.; Zhan, H.X.; Zhang, F.; Dirk, B.; Zhong, Y.Z.; Lou, Q.Z.; Zhong, Y.; Zhang, J.P. Development and fecundity of Trissolcus japonicus on fertilized and unfertilized eggs of the brown marmorated stink bug, Halyomorpha halys. J. Pest. Sci. 2018, 91, 1–9. [Google Scholar] [CrossRef]
- Bjorksten, T.A.; Hoffmann, A.A. Plant cues influence searching behaviour and parasitism in the egg parasitoid Trichogramma nr. brassicae. Ecol. Entomol. 1998, 23, 355–362. [Google Scholar] [CrossRef]
- DeLury, N.C.; Gries, R.; Gries, G.; Judd, G.J.R.; Khaskin, G. Moth scale-derived kairomones used by egg-larval parasitoid Ascogaster quadridentatato locate eggs of its host, Cydia pomonella. J. Chem. Ecol. 1999, 25, 2419–2431. [Google Scholar] [CrossRef]
- Meiners, T.; Westerhaus, C.; Hilker, M. Specificity of chemical cues used by a specialist egg parasitoid during host location. Entomol. Exp. Appl. 2000, 95, 151–159. [Google Scholar] [CrossRef]
- Boyle, S.M.; Weber, D.C.; Hough-Goldstein, J.; Hoelmer, K.A. Host kairomones influence searching behavior of Trissolcus japonicus (Hymenoptera: Scelionidae), a parasitoid of Halyomorpha halys (Heteroptera: Pentatomidae). Environ. Entomol. 2020, 49, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pak, G.A.; van Dalen, A.; Kaashoek, N.; Dijkman, H. Host egg chorion structure influencing host suitability for the egg parasitoid Trichogramma Westwood. J. Insect Physiol. 1990, 36, 869–875. [Google Scholar] [CrossRef]
- Karamaouna, F.; Copland, M.J.W. Host suitability, quality and host size preference of Leptomastix epona and Pseudaphycus flavidulus, two endoparasitoids of the mealybug Pseudococcus viburni, and host size effect on parasitoid sex ratio and clutch size. Entomol. Exp. Appl. 2000, 96, 149–158. [Google Scholar] [CrossRef]
- Godfray, H.C.J. Parasitoids. Behavior and Evolutionary Ecology; Princeton University Press: Princeton, NJ, USA, 1994. [Google Scholar]
- Desneux, N.; Barta, R.J.; Hoelmer, K.A.; Hopper, K.R.; Heimpel, G.E. Multifaceted determinants of host specificity in an aphid parasitoid. Oecologia 2009, 160, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Biondi, A.; Desneux, N.; Amiens-Desneux, E.; Siscaro, G.; Zappalà, L. Biology and developmental strategies of the palaearctic parasitoid Bracon nigricans (Hymenoptera: Braconidae) on the neotropical moth Tuta absoluta (Lepidoptera: Gelechiidae). J. Econ. Entomol. 2013, 106, 1638–1647. [Google Scholar] [CrossRef]
- Monticelli, L.S.; Nguyen, L.T.; Amiens-Desneux, E.; Luo, C.; Lavoir, A.V.; Gatti, J.L.; Desneux, N. The preference-performance relationship as a means of classifying parasitoids according to their specialization degree. Evol. Appl. 2019, 12, 1626–1640. [Google Scholar] [CrossRef]
- Van Alphen, J.J.M.; Visser, M.E. Superparasitism as an adaptive strategy for insect parasitoids. Annu. Rev. Entomol. 1990, 35, 59–79. [Google Scholar] [CrossRef]
- Gu, H.; Wang, Q.; Dorn, S. Superparasitism in Cotesia glomerata: Response of hosts and consequences for parasitoids. Ecol. Entomol. 2003, 28, 422–431. [Google Scholar] [CrossRef]
- King, B.H. Offspring sex ratio and number in response to proportion of host sizes and ages in the parasitoid wasp Spalangia cameroni (Hymenoptera: Pteromalidae). Environ. Entomol. 2002, 31, 505–508. [Google Scholar] [CrossRef] [Green Version]
- Visser, M.E. The importance of being large: The relationship between size and fitness in females of the parasitoid Aphaereta minuta (Hymenoptera: Brachonidae). J. Anim. Ecol. 1994, 63, 963–978. [Google Scholar] [CrossRef]
- West, S.A.; Flanagan, K.E.; Godfray, H.C.J. The relationship between parasitoid size and fitness in the field, a study of Achrysocharoides zwoelferi (Hymenoptera: Eulophidae). J. Anim. Ecol. 1996, 65, 631–639. [Google Scholar] [CrossRef]
- Ellers, J.; Van Alphen, J.J.M.; Sevenster, J.G. A field study of size-fitness relationships in the parasitoid Asobara tabida. J. Anim. Ecol. 1998, 67, 318–324. [Google Scholar] [CrossRef]
- Durocher-Granger, L.; Martel, V.; Boivin, G. Gamete number and size correlate with adult size in the egg parasitoid Trichogramma euproctidis. Entomol. Exp. Appl. 2011, 140, 262–268. [Google Scholar] [CrossRef]
- Yanagi, S.; Tuda, M. Female size constrains egg size via the influence of reproductive organ size and resource storage in the seed beetle Callosobruchus chinensis. J. Insect Physiol. 2012, 58, 1432–1437. [Google Scholar] [CrossRef]
- Wang, J.J.; Liu, X.B.; Zhang, Y.A.; Wen, C.; Wei, J.R. The reproductive capability of Ooencyrtus kuvanae reared on eggs of the factitious host Antheraea pernyi. J. Appl. Entomol. 2014, 138, 267–272. [Google Scholar] [CrossRef]
- Drost, Y.C.; Cardé, R.T. Influence of host deprivation on egg load and oviposition behaviour of Brachymeria intermedia, a parasitoid of gypsy moth. Physiol. Entomol. 1992, 17, 230–234. [Google Scholar] [CrossRef]
- Li, X.W.; Zhu, L.T.; Meng, L.; Li, B.P. Brood size and sex ratio in response to host quality and wasp traits in the gregarious parasitoid Oomyzus sokolowskii (Hymenoptera: Eulophidae). PeerJ 2017, 5, e2919. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Chen, Y.-M.; Yang, X.-B.; Lv, R.-E.; Desneux, N.; Zang, L.-S. Parasitism and Suitability of Aprostocetus brevipedicellus on Chinese Oak Silkworm, Antheraea pernyi, a Dominant Factitious Host. Insects 2021, 12, 694. https://doi.org/10.3390/insects12080694
Wang J, Chen Y-M, Yang X-B, Lv R-E, Desneux N, Zang L-S. Parasitism and Suitability of Aprostocetus brevipedicellus on Chinese Oak Silkworm, Antheraea pernyi, a Dominant Factitious Host. Insects. 2021; 12(8):694. https://doi.org/10.3390/insects12080694
Chicago/Turabian StyleWang, Jing, Yong-Ming Chen, Xiang-Bing Yang, Rui-E Lv, Nicolas Desneux, and Lian-Sheng Zang. 2021. "Parasitism and Suitability of Aprostocetus brevipedicellus on Chinese Oak Silkworm, Antheraea pernyi, a Dominant Factitious Host" Insects 12, no. 8: 694. https://doi.org/10.3390/insects12080694
APA StyleWang, J., Chen, Y.-M., Yang, X.-B., Lv, R.-E., Desneux, N., & Zang, L.-S. (2021). Parasitism and Suitability of Aprostocetus brevipedicellus on Chinese Oak Silkworm, Antheraea pernyi, a Dominant Factitious Host. Insects, 12(8), 694. https://doi.org/10.3390/insects12080694