
Insect Collections as an Untapped Source of Bioactive Compounds—Fireflies (Coleoptera: Lampyridae) and Cardiotonic Steroids as a Proof of Concept


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Non-Destructive Extraction of Dry Museum Specimens
2.2. DAD-HPLC and DAD-LC/ESI-Qq-TOF-mass Spectrometry of Beetle Extracts
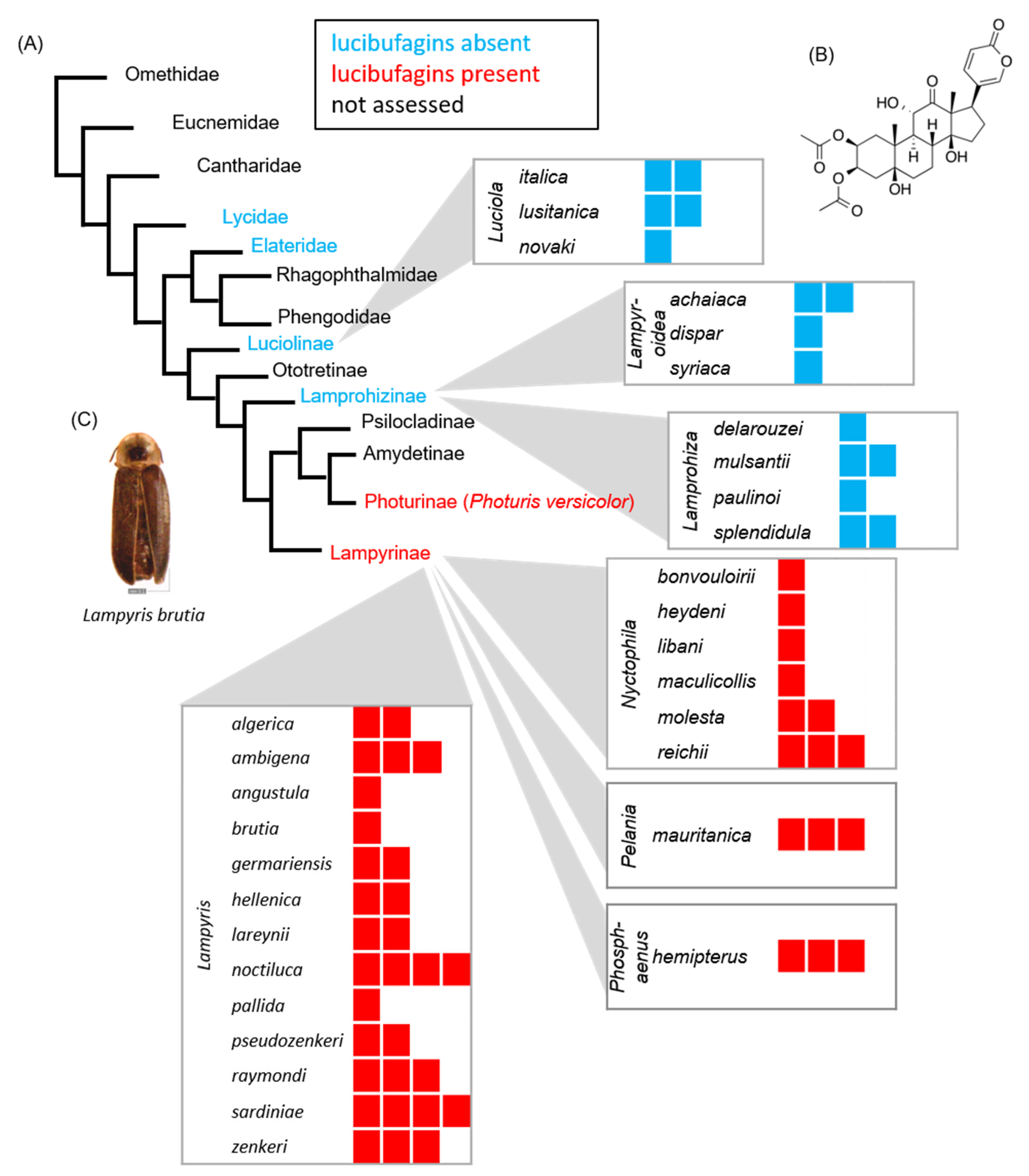
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shaffer, H.B.; Fisher, R.N.; Davidson, C. The role of natural history collections in documenting species declines. Trends Ecol. Evol. 1998, 13, 27–30. [Google Scholar] [CrossRef]
- Lister, A.M. Natural history collections as sources of long-term datasets. Trends Ecol. Evol. 2011, 26, 153–154. [Google Scholar] [CrossRef] [Green Version]
- Holmes, M.W.; Hammond, T.T.; Wogan, G.O.U.; Walsh, R.E.; Labarbera, K.; Wommack, E.A.; Martins, F.M.; Crawford, J.C.; Mack, K.L.; Bloch, L.M.; et al. Natural history collections as windows on evolutionary processes. Mol. Ecol. 2016, 25, 864–881. [Google Scholar] [CrossRef] [Green Version]
- Cook, J.A.; Edwards, S.V.; Lacey, E.A.; Guralnick, R.P.; Soltis, P.S.; Soltis, D.E.; Welch, C.K.; Bell, K.C.; Galbreath, K.E.; Himes, C.; et al. Natural history collections as emerging resources for innovative education. Bioscience 2014, 64, 725–734. [Google Scholar] [CrossRef] [Green Version]
- Scudder, G.G.E.; Duffey, S.S. Cardiac glycosides in the Lygaeinae (Hemiptera: Lygaeidae). Can. J. Zool. 1972, 50, 35–42. [Google Scholar] [CrossRef]
- Zangerl, A.R.; Berenbaum, M.R. Increase in toxicity of an invasive weed after reassociation with its coevolved herbivore. Proc. Natl. Acad. Sci. USA 2005, 102, 15529–15532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petschenka, G.; Halitschke, R.; Roth, A.; Stiehler, S.; Tenbusch, L.; Züst, T.; Hartwig, C.; Gámez, J.F.M.; Trusch, R.; Deckert, J.; et al. Predation drives specialized host plant associations in preadapted milkweed bugs (Heteroptera: Lygaeinae). bioRxiv 2020. [Google Scholar] [CrossRef]
- Branham, M.A. Lampyridae Latreille. In Handbook of Zoology 2010, Volume IV: Arthropoda: Insecta, Part 38. Coleoptera, Beetles, Volume 2. Morphology and Systematics (Polyphaga Partim); Beutel, R.G., Leschen, R.A.B., Lawrence, J.F., Eds.; Walter DeGruyter: Berlin, Germany, 1817; pp. 141–149. [Google Scholar]
- Eisner, T.; Wiemer, D.F.; Haynes, L.W.; Meinwald, J. Lucibufagins: Defensive steroids from the fireflies Photinus ignitus and P. marginellus (Coleoptera: Lampyridae). Proc. Natl. Acad. Sci. USA 1978, 75, 905–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goetz, M.; Wiemer, D.F.; Haynes, L.R.W.; Meinwald, J.; Eisner, T. Lucibufagines. Partie III. Oxo-11-et oxo-12-bufalines, stéroïdes défensifs des lampyres Photinus ignitus et P. marginellus (Coleoptera: Lampyridae). Helv. Chim. Acta 1979, 62, 1396–1400. [Google Scholar] [CrossRef]
- Meinwald, J.; Wiemer, D.F.; Eisner, T. Lucibufagins. 2. Esters of 12-oxo-2β, 5β, 11α-trihydroxybufalin, the major defensive steroids of the firefly Photinus pyralis (Coleoptera: Lampyridae). J. Am. Chem. Soc. 1979, 101, 3055–3060. [Google Scholar] [CrossRef]
- Goetz, M.A.; Meinwald, J.; Eisner, T. Lucibufagins, IV. New defensive steroids and a pterin from the firefly, Photinus pyralis (Coleoptera: Lampyridae). Experientia 1981, 37, 679–680. [Google Scholar] [CrossRef]
- González, A.; Schroeder, F.C.; Attygalle, A.B.; Svatoš, A.; Meinwald, J.; Eisner, T. Metabolic transformations of acquired lucibufagins by firefly “femmes fatales”. Chemoecology 1999, 9, 105–112. [Google Scholar] [CrossRef]
- Eisner, T.; Goetz, M.A.; Hill, D.E.; Smedley, S.R.; Meinwald, J. Firefly “femmes fatales” acquire defensive steroids (lucibufagins) from their firefly prey. Proc. Natl. Acad. Sci. USA 1997, 94, 9723–9728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gronquist, M.; Meinwald, J.; Eisner, T.; Schroeder, F.C. Exploring uncharted terrain in nature’s structure space using capillary NMR spectroscopy: 13 Steroids from 50 fireflies. J. Am. Chem. Soc. 2005, 127, 10810–10811. [Google Scholar] [CrossRef]
- Smedley, S.R.; Risteen, R.G.; Tonyai, K.K.; Pitino, J.C.; Hu, Y.; Ahmed, Z.B.; Christofel, B.T.; Gaber, M.; Howells, N.R.; Mosey, C.F.; et al. Bufadienolides (lucibufagins) from an ecologically aberrant firefly (Ellychnia corrusca). Chemoecology 2017, 27, 141–153. [Google Scholar] [CrossRef]
- Yoshida, T.; Ujiie, R.; Savitzky, A.H.; Jono, T.; Inoue, T.; Yoshinaga, N.; Aburaya, S.; Aoki, W.; Takeuchi, H.; Ding, L.; et al. Dramatic dietary shift maintains sequestered toxins in chemically defended snakes. Proc. Natl. Acad. Sci. USA 2020, 117, 5964–5969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyler, J.; McKinnon, W.; Lord, G.A.; Hilton, P.J. A defensive steroidal pyrone in the glow-worm Lampyris noctiluca L. (Coleoptera: Lampyridae). Physiol. Entomol. 2008, 33, 167–170. [Google Scholar] [CrossRef]
- Agrawal, A.A.; Petschenka, G.; Bingham, R.A.; Weber, M.G.; Rasmann, S. Toxic cardenolides: Chemical ecology and coevolution of specialized plant-herbivore interactions. New Phytol. 2012, 194, 28–45. [Google Scholar] [CrossRef]
- Paula, S.; Tabet, M.R.; Ball, W.J. Interactions between cardiac glycosides and sodium/potassium-ATPase: Three-dimensional structure−activity relationship models for ligand binding to the E2-Pi form of the enzyme versus activity inhibition. Biochemistry 2005, 44, 498–510. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Guo, D. Analysis of bufadienolides in the Chinese drug ChanSu by high-performance liquid chromatography with atmospheric pressure chemical ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2005, 19, 1881–1892. [Google Scholar] [CrossRef]
- Gao, H.; Popescu, R.; Kopp, B.; Wang, Z. Bufadienolides and their antitumor activity. Nat. Prod. Rep. 2011, 28, 953–969. [Google Scholar] [CrossRef]
- Hutchinson, D.A.; Mori, A.; Savitzky, A.H.; Burghardt, G.M.; Wu, X.; Meinwald, J.; Schroeder, F.C. Dietary sequestration of defensive steroids in nuchal glands of the Asian snake Rhabdophis tigrinus. Proc. Natl. Acad. Sci. USA 2007, 104, 2265–2270. [Google Scholar] [CrossRef] [Green Version]
- Fallon, T.R.; Lower, S.E.; Chang, C.-H.; Bessho-Uehara, M.; Martin, G.J.; Bewick, A.J.; Behringer, M.; Debat, H.J.; Wong, I.; Day, J.C.; et al. Firefly genomes illuminate parallel origins of bioluminescence in beetles. Elife 2018, 7, e36495. [Google Scholar] [CrossRef] [PubMed]
- Leavell, B.C.; Rubin, J.J.; McClure, C.J.W.; Miner, K.A.; Branham, M.A.; Barber, J.R. Fireflies thwart bat attack with multisensory warnings. Sci. Adv. 2018, 4, eaat6601. [Google Scholar] [CrossRef] [Green Version]
- Blum, M.S.; Sannasi, A. Reflex bleeding in the lampyrid Photinus pyralis: Defensive function. J. Insect Physiol. 1974, 20, 451–460. [Google Scholar] [CrossRef]
- Brower, L.P.; Glazier, S.C. Localization of heart poisons in the monarch butterfly. Science 1975, 188, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Knight, M.; Glor, R.; Smedley, S.R.; González, A.; Adler, K.; Eisner, T. Firefly toxicosis in lizards. J. Chem. Ecol. 1999, 25, 1981–1986. [Google Scholar] [CrossRef]
- Lloyd, J.E. Firefly parasites and predators. Coleopt. Bull. 1973, 27, 91–106. [Google Scholar]
- Underwood, T.J.; Tallamy, D.W.; Pesek, J.D. Bioluminescence in firefly larvae: A test of the aposematic display hypothesis (Coleoptera: Lampyridae). J. Insect Behav. 1997, 10, 365–370. [Google Scholar] [CrossRef]
- Moosman, P.R.; Cratsley, C.K.; Lehto, S.D.; Thomas, H.H. Do courtship flashes of fireflies (Coleoptera: Lampyridae) serve as aposematic signals to insectivorous bats? Anim. Behav. 2009, 78, 1019–1025. [Google Scholar] [CrossRef]
- Krivoruchko, K.; Goldshtein, A.; Boonman, A.; Eitan, O.; Ben-Simon, J.; Thong, V.D.; Yovel, Y. Fireflies produce ultrasonic clicks during flight as a potential aposematic anti-bat signal. iScience 2021, 24, 102194. [Google Scholar] [CrossRef]
- Pettit, G.R. Higher Plant Steroids BT-Biosynthetic Products for Cancer Chemotherapy; Pettit, G.R., Ed.; Springer US: Boston, MA, USA, 1977; pp. 89–95. Volume 1, ISBN 978-1-4684-7230-1. [Google Scholar]
- Martin, G.J.; Stanger-Hall, K.F.; Branham, M.A.; Da Silveira, L.F.L.; Lower, S.E.; Hall, D.W.; Li, X.-Y.; Lemmon, A.R.; Moriarty Lemmon, E.; Bybee, S.M. Higher-level phylogeny and reclassification of Lampyridae (Coleoptera: Elateroidea). Insect Syst. Divers. 2019, 3. [Google Scholar] [CrossRef]
- Binkert, J.; Angliker, E.; Von Wartburg, A. Über die Autoxydation des Strophanthidins (1. Teil). 49. Mitteilung über Herzglykoside. Helv. Chim. Acta 1962, 45, 2122–2138. (In German) [Google Scholar] [CrossRef]
- Malcolm, S.B. Cardenolide-mediated interactions between plants and herbivores. In Herbivores: Their Interactions with Secondary Plant Metabolites, 2nd ed.; Rosenthal, G.A., Berenbaum, M.R., Eds.; The Chemical Participants; Academic Press: San Diego, CA, USA, 1991; Volume 1, pp. 251–296. [Google Scholar]
- Lüddecke, T.; Vilcinskas, A.; Lemke, S. Phylogeny-guided selection of priority groups for venom bioprospecting: Harvesting toxin sequences in tarantulas as a case study. Toxins 2019, 11, 488. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Species | Total Lucibufagins per Beetle [µg] | Number of Lucibufagin Peaks |
|---|---|---|
| Lampyris algerica levigata (Geisthardt 1983) | 154/165 | 24/25 |
| Lampyris ambigena (Jacquelin du Val 1860) | 7/63/185 | 9/20/22 |
| Lampyris angustula (Fairmaire 1895) | 41 | 21 |
| Lampyris brutia (Costa 1882) | 123 | 24 |
| Lampyris germariensis (Jacquelin du Val 1860) | 3/60 | 6/18 |
| Lampyris hellenica (Geisthardt 1983) | 114/668 | 24/41 |
| Lampyris lareynii (Jacquelin du Val 1859) | 29/78 | 19/21 |
| Lampyris noctiluca (Linnaeus 1767) | 88/118/142/615 | 24/14/22/30 |
| Lampyris pallida (Geisthardt 1987) | 34 | 23 |
| Lampyris pseudozenkeri (Geisthardt 1999) | 11/19 | 21/21 |
| Lampyris raymondi (Mulsant and Rey 1859) | 5/133/313 | 6/21/24 |
| Lampyris sardiniae (Geisthardt 1987) | 2/77/125/138 | 4/26/38/32 |
| Lampyris zenkeri (Germar 1817) | 53/114/210 | 14/21/16 |
| Nyctophila heydeni (Olivier 1884) | 55 | 11 |
| Nyctophila maculicollis (Fairmaire 1866) | 102 | 19 |
| Nyctophila molesta (Jacquelin du Val 1859) | 291/323 | 30/22 |
| Nyctophila reichii (Jacquelin du Val 1859) | 56/217/309 | 11/28/21 |
| Pelania mauritanica (Linnaeus 1767) | 64/159/194 | 16/22/25 |
| Phosphaenus hemipterus (Goeze 1777) | 9/18/38 | 18/24/29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berger, A.; Petschenka, G.; Degenkolb, T.; Geisthardt, M.; Vilcinskas, A. Insect Collections as an Untapped Source of Bioactive Compounds—Fireflies (Coleoptera: Lampyridae) and Cardiotonic Steroids as a Proof of Concept. Insects 2021, 12, 689. https://doi.org/10.3390/insects12080689
Berger A, Petschenka G, Degenkolb T, Geisthardt M, Vilcinskas A. Insect Collections as an Untapped Source of Bioactive Compounds—Fireflies (Coleoptera: Lampyridae) and Cardiotonic Steroids as a Proof of Concept. Insects. 2021; 12(8):689. https://doi.org/10.3390/insects12080689
Chicago/Turabian StyleBerger, Andreas, Georg Petschenka, Thomas Degenkolb, Michael Geisthardt, and Andreas Vilcinskas. 2021. "Insect Collections as an Untapped Source of Bioactive Compounds—Fireflies (Coleoptera: Lampyridae) and Cardiotonic Steroids as a Proof of Concept" Insects 12, no. 8: 689. https://doi.org/10.3390/insects12080689
APA StyleBerger, A., Petschenka, G., Degenkolb, T., Geisthardt, M., & Vilcinskas, A. (2021). Insect Collections as an Untapped Source of Bioactive Compounds—Fireflies (Coleoptera: Lampyridae) and Cardiotonic Steroids as a Proof of Concept. Insects, 12(8), 689. https://doi.org/10.3390/insects12080689