
Pupal Cues Increase Sperm Production but Not Testis Size in an Insect



Abstract
Simple Summary
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Insects
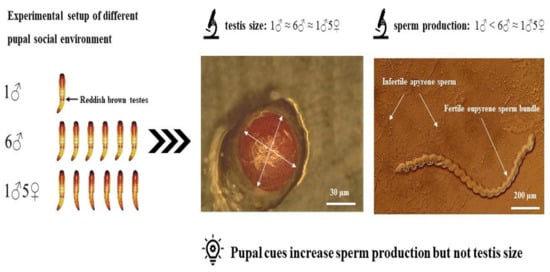
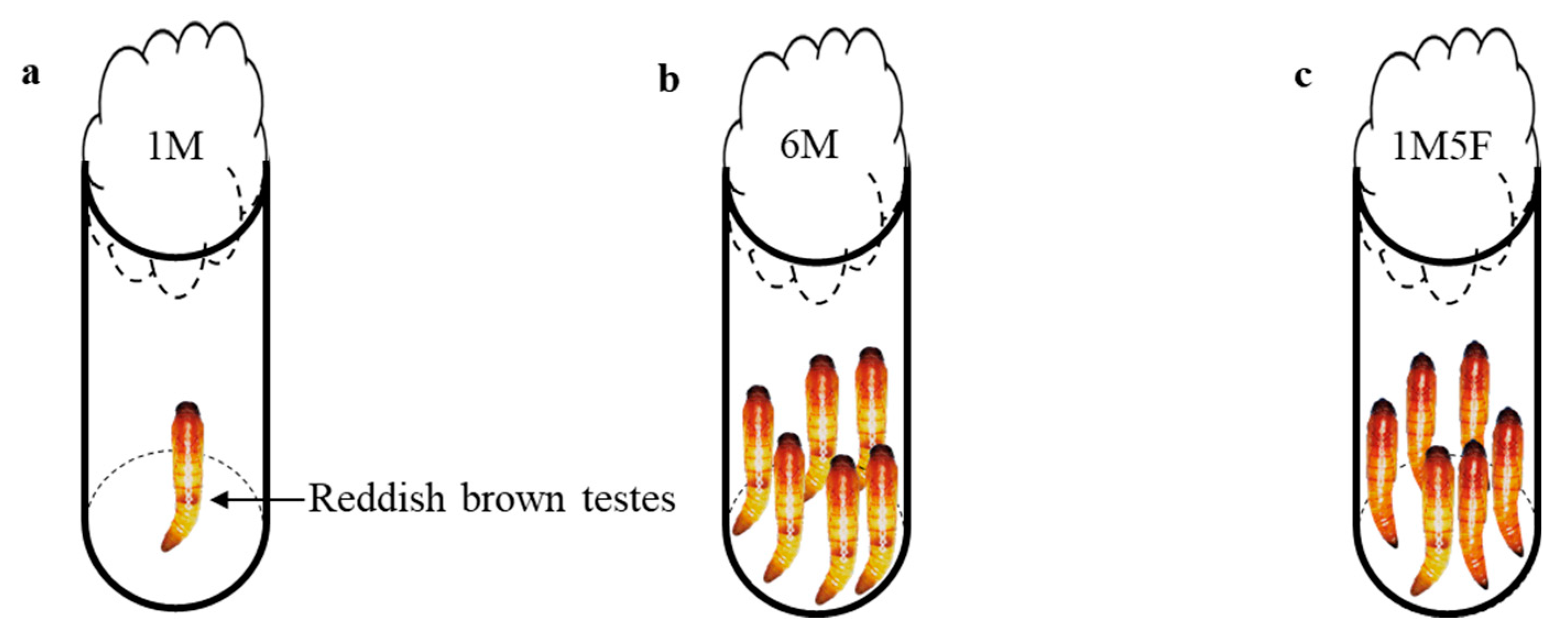
2.2. Experimental Setup and Data Collection
2.3. Statistical Analysis
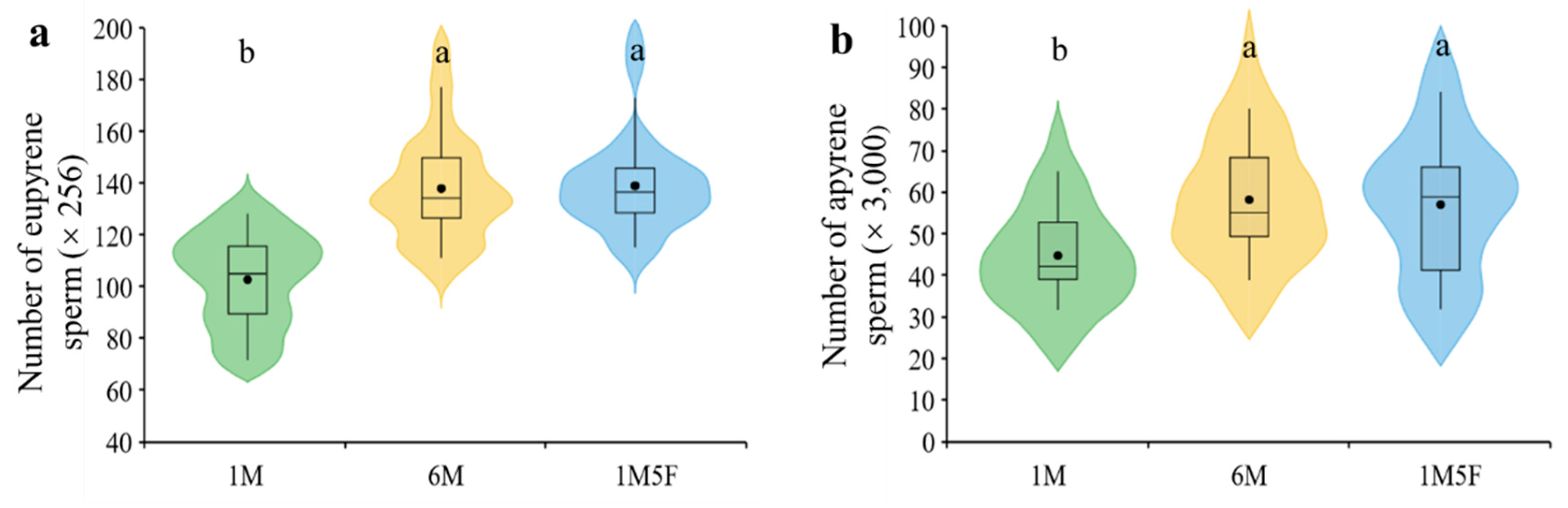
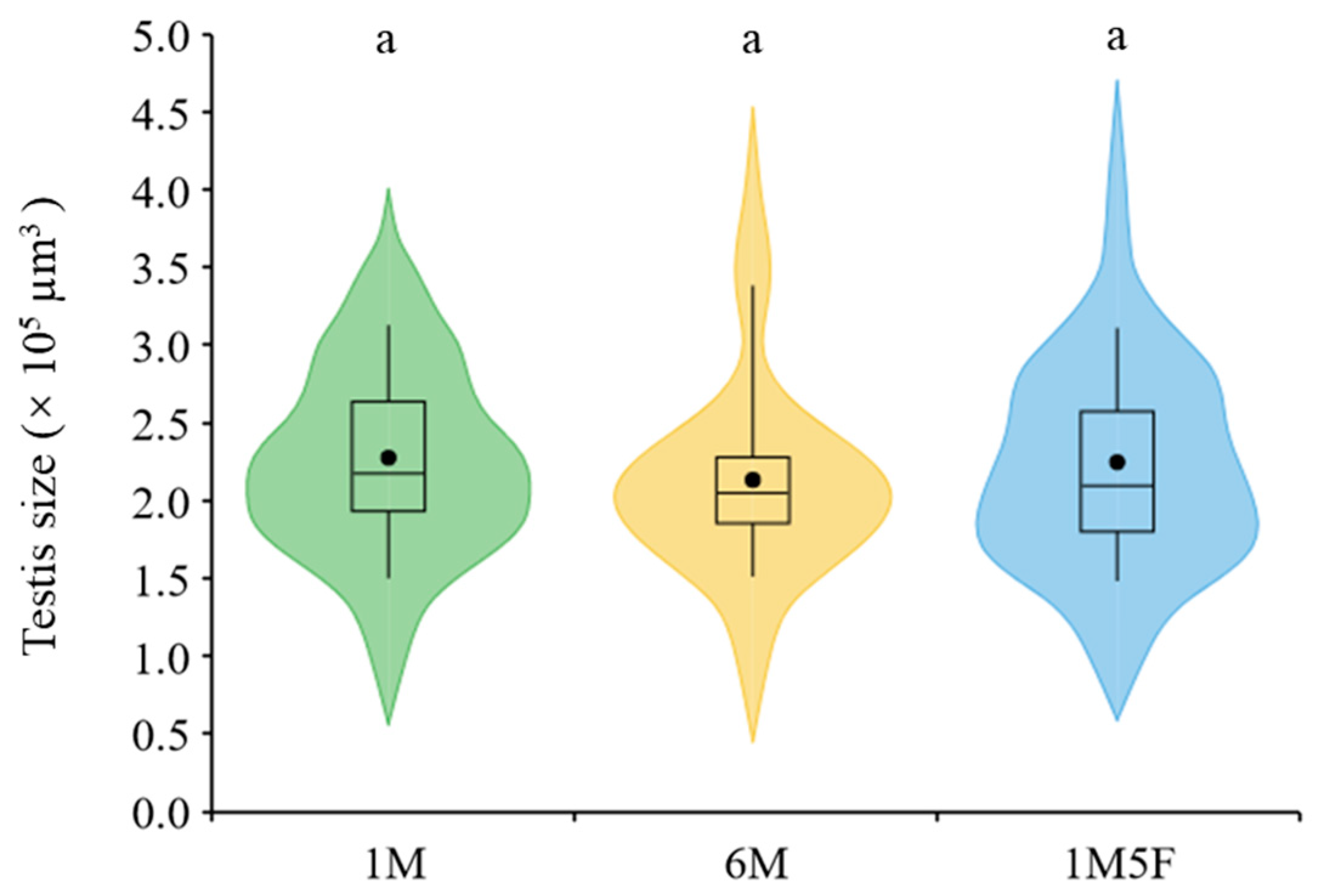
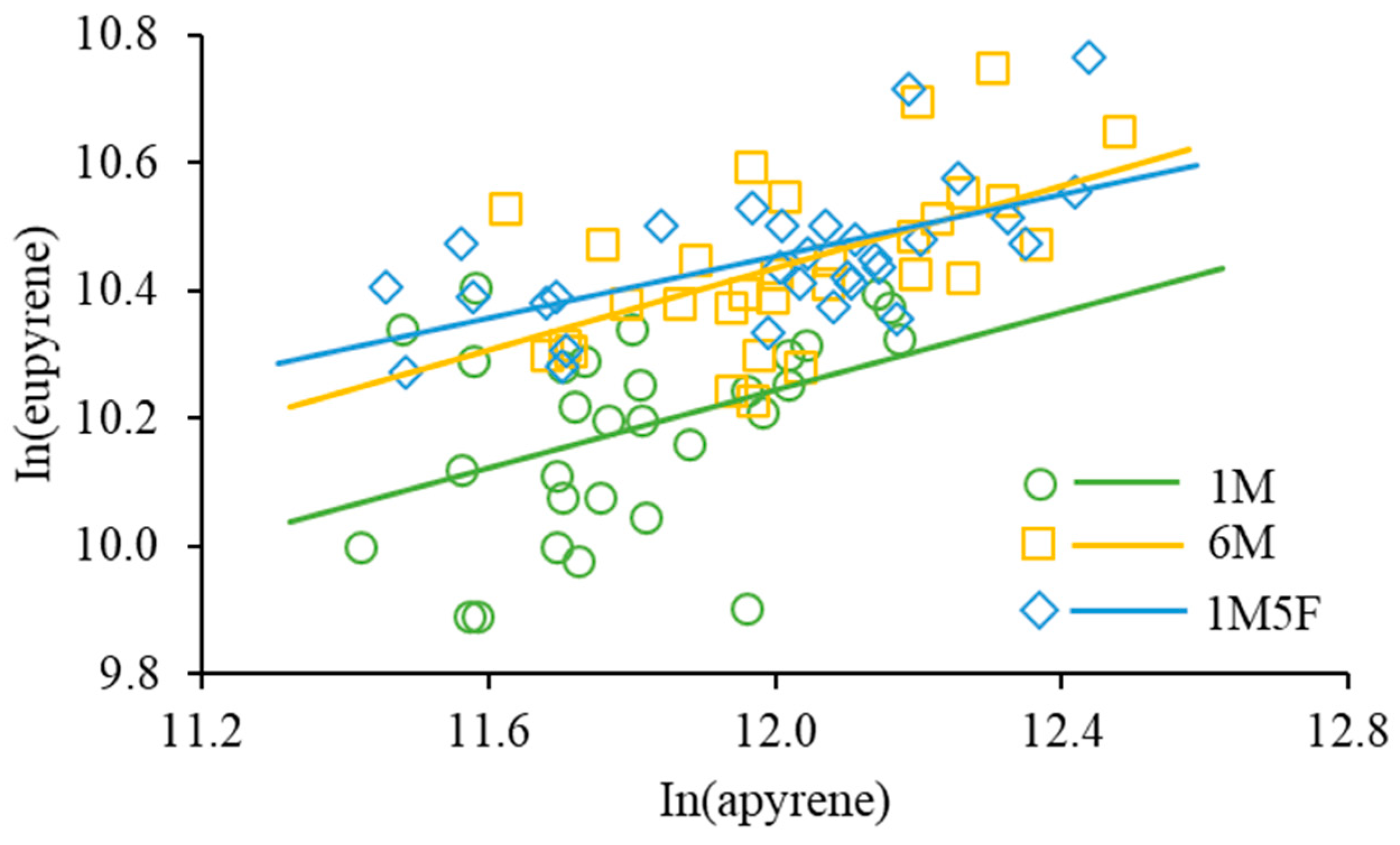
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dewsbury, D.A. Ejaculate cost and male choice. Am. Nat. 1982, 119, 601–610. [Google Scholar] [CrossRef]
- Wedell, N.; Gage, M.J.G.; Parker, G.A. Sperm competition, male prudence and sperm-limited females. Trends Ecol. Evol. 2002, 17, 313–320. [Google Scholar] [CrossRef]
- Dore, A.A.; Bretman, A.; Chapman, T. Fitness consequences of redundant cues of competition in male Drosophila melanogaster. Ecol. Evol. 2020, 10, 5517–5526. [Google Scholar] [CrossRef]
- Bjork, A.; Dallai, R.; Pitnick, S. Adaptive modulation of sperm production rate in Drosophila bifurca, a species with giant sperm. Biol. Lett. 2007, 3, 517–519. [Google Scholar] [CrossRef] [PubMed]
- Parker, G.A.; Pizzari, T. Sperm competition and ejaculate economics. Biol. Rev. 2010, 85, 897–934. [Google Scholar] [CrossRef]
- Kelly, C.D.; Jennions, M.D. Sexual selection and sperm quantity: Meta-analyses of strategic ejaculation. Biol. Rev. 2011, 86, 863–884. [Google Scholar] [CrossRef]
- Firman, R.C.; Garcia-Gonzalez, F.; Simmons, L.W.; Andre, G.I. A competitive environment influences sperm production, but not testes tissue composition, in house mice. J. Evol. Biol. 2018, 31, 1647–1654. [Google Scholar] [CrossRef]
- Ramm, S.A.; Stockley, P. Adaptive plasticity of mammalian sperm production in response to social experience. Proc. Biol. Sci. B Biol. Sci. 2009, 276, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Moatt, J.P.; Dytham, C.; Thom, M.D. Sperm production responds to perceived sperm competition risk in male Drosophila melanogaster. Physiol. Behav. 2014, 131, 111–114. [Google Scholar] [CrossRef]
- Liu, J.Y.; Zhang, Y.J.; Zheng, X.-L.; He, X.Z.; Wang, Q. Combined cues of male competition influence spermatozoal investment in a moth. Funct. Ecol. 2020, 34, 1223–1234. [Google Scholar] [CrossRef]
- Simmons, L.W.; Denholm, A.; Jackson, C.; Levy, E.; Madon, E. Male crickets adjust ejaculate quality with both risk and intensity of sperm competition. Biol. Lett. 2007, 3, 520–522. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, Q. Ejaculate economics: An experimental test in a moth. Biol. Lett. 2014, 10, 20131031. [Google Scholar] [CrossRef] [PubMed]
- Larsdotter-Mellström, H.; Wiklund, C. Different mating expenditure in response to sperm competition risk between generations in the bivoltine butterfly Pieris napi. Behav. Ecol. Sociobiol. 2015, 69, 1067–1074. [Google Scholar] [CrossRef]
- Simmons, L.W.; Lovegrove, M. Socially cued seminal fluid gene expression mediates responses in ejaculate quality to sperm competition risk. Proc. Biol. Sci. B Biol. Sci. 2017, 284, 20171486. [Google Scholar] [CrossRef] [PubMed]
- Lymbery, S.J.; Tomkins, J.L.; Simmons, L.W. Male responses to sperm competition when rivals vary in number and familiarity. Proc. Biol. Sci. B Biol. Sci. 2019, 286, 20182589. [Google Scholar] [CrossRef]
- Esfandi, K.; He, X.Z.; Wang, Q. Sperm allocation strategies in a sperm heteromorphic insect. Curr. Zool. 2020, 66, 285–292. [Google Scholar] [CrossRef]
- Gage, M.J.G. Continuous variation in reproductive strategy as an adaptive response to population density in the moth Plodia interpunctella. Proc. Biol. Sci. B Biol. Sci. 1995, 261, 25–30. [Google Scholar] [CrossRef]
- McNamara, K.B.; Simmons, L.W. Experimental evolution reveals differences between phenotypic and evolutionary responses to population density. J. Evol. Biol. 2017, 30, 1763–1771. [Google Scholar] [CrossRef]
- Liu, J.Y.; Zhang, Y.J.; Zheng, X.-L.; He, X.Z.; Wang, Q. Larval social cues influence testicular investment in an insect. Curr. Zool. 2021, zoab028. [Google Scholar] [CrossRef]
- McNamara, K.B.; Elgar, M.A.; Jones, T.M. Adult responses to larval population size in the almond moth, Cadra cautella. Ethology 2010, 116, 39–46. [Google Scholar] [CrossRef]
- He, Y.; Miyata, T. Variations in sperm number in relation to larval crowding and spermatophore size in the armyworm, Pseudaletia separata. Ecol. Entomol. 1997, 22, 41–46. [Google Scholar] [CrossRef]
- Hinton, H.E. Sound production in lepidopterous pupae. Entomologist 1948, 81, 254–269. [Google Scholar]
- Travassos, M.A.; Pierce, N.E. Acoustics, context and function of vibrational signalling in a lycaenid butterfly-ant mutualism. Anim. Behav. 2000, 60, 13–26. [Google Scholar] [CrossRef]
- Casacci, L.P.; Bonelli, S.; Balletto, E.; Barbero, F. Multimodal signaling in myrmecophilous butterflies. Front. Ecol. Evol. 2019, 7, 454. [Google Scholar] [CrossRef]
- Lin, Y.H.; Liao, Y.C.; Yang, C.S.; Billen, J.; Yang, M.M.; Hsu, Y.F. Vibrational communication between a myrmecophilous butterfly Spindasis lohita (Lepidoptera: Lycaenidae) and its host ant Crematogaster rogenhoferi (Hymenoptera: Formicidae). Sci. Rep. 2019, 9, 18548. [Google Scholar] [CrossRef] [PubMed]
- Downey, J.C. Sound production in pupae of Lycaenidae. J. Lepid. Soc. 1966, 20, 129–155. [Google Scholar]
- Álvarez, M.; Munguira, M.L.; Martinez-Ibanez, M.D. Comparative study of the morphology of stridulatory organs of the Iberian lycaenid butterfly pupae (Lepidoptera). J. Morphol. 2014, 275, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Dolle, P.; Klein, P.; Fischer, O.W.; Schnitzler, H.U.; Gilbert, L.E.; Boppre, M. Twittering pupae of Papilionid and Nymphalid butterflies (Lepidoptera): Novel structures and sounds. Ann. Entomol. Soc. Am. 2018, 111, 341–354. [Google Scholar] [CrossRef]
- Alexander, A.J. A study of the biology and behavior of the caterpillars, pupae and emerging butterflies of the subfamily Heliconiinae in Trinidad, West Indies. II. Molting, and the behavior of pupae and emerging adults. Zoologica 1961, 46, 105–124. [Google Scholar]
- Jefferson, R.N.; Rubin, R.E. Sex pheromones of Lepidoptera. Xxxv. Pupal and postemergence development of the pheromone gland of Trichoplusia ni. Ann. Entomol. Soc Am. 1973, 66, 277–279. [Google Scholar] [CrossRef]
- Choi, M.Y.; Lim, H.; Park, K.C.; Adlof, R.; Wang, S.; Zhang, A.; Jurenka, R. Identification and biosynthetic studies of the hydrocarbon sex pheromone in Utetheisa ornatrix. J. Chem. Ecol. 2007, 33, 1336–1345. [Google Scholar] [CrossRef][Green Version]
- Feng, K.C.; Roelofs, W.L. Sex pheromone gland development in redbanded leafroller moth, Argyrotaenia velutinana, pupae and adults. Ann. Entomol. Soc. Am. 1977, 70, 721–732. [Google Scholar] [CrossRef]
- Tang, J.D.; Wolf, W.A.; Roelofs, W.L.; Knipple, D.C. Development of functionally competent cabbage looper moth sex pheromone glands. Insect Biochem. 1991, 21, 573–581. [Google Scholar] [CrossRef]
- Kwadha, C.A.; Mutunga, J.M.; Irungu, J.; Ongamo, G.; Ndegwa, P.; Raina, S.; Fombong, A.T. Decanal as a major component of larval aggregation pheromone of the greater wax moth, Galleria mellonella. J. Appl. Entomol. 2019, 143, 417–429. [Google Scholar] [CrossRef]
- Duthie, B.; Gries, G.; Gries, R.; Krupke, C.; Derksen, S. Does pheromone-based aggregation of codling moth larvae help procure future mates? J. Chem. Ecol. 2003, 29, 425–436. [Google Scholar] [CrossRef]
- Calvert, I.A.N.; Corbet, S.A. Reproductive maturation and pheromone release in the flour moth Anagasta kuehniella (Zeller). J. Ent. Ser. A-Gen. 1973, 47, 201–209. [Google Scholar] [CrossRef]
- Pontier, S.M.; Schweisguth, F. A wolbachia-sensitive communication between male and female pupae controls gamete compatibility in Drosophila. Curr. Biol. 2015, 25, 2339–2348. [Google Scholar] [CrossRef]
- Estrada, C.; Yildizhan, S.; Schulz, S.; Gilbert, L.E. Sex-specific chemical cues from immatures facilitate the evolution of mate guarding in Heliconius butterflies. Proc. Biol. Sci. B Biol. Sci. 2010, 277, 407–413. [Google Scholar] [CrossRef]
- Stockley, P.; Seal, N.J. Plasticity in reproductive effort of male dung flies (Scatophaga stercoraria) as a response to larval density. Funct. Ecol. 2001, 15, 96–102. [Google Scholar] [CrossRef]
- Johnson, T.L.; Symonds, M.R.E.; Elgar, M.A. Anticipatory flexibility: Larval population density in moths determines male investment in antennae, wings and testes. Proc. Biol. Sci. B Biol. Sci. 2017, 284, 20172087. [Google Scholar] [CrossRef]
- Lüpol, S.; de Boer, R.A.; Evans, J.P.; Tomkins, J.L.; Fitzpatrick, J.L. How sperm competition shapes the evolution of testes and sperm: A meta-analysis. Philos. Trans. R. Soc. B 2020, 375, 20200064. [Google Scholar] [CrossRef] [PubMed]
- Nijhout, H.F.; Emlen, D.J. Competition among body parts in the development and evolution of insect morphology. Proc. Natl. Acad. Sci. USA 1998, 95, 3685–3689. [Google Scholar] [CrossRef] [PubMed]
- Moczek, A.P.; Nijhout, H.F. Trade-offs during the development of primary and secondary sexual traits in a horned beetle. Am. Nat. 2004, 163, 184–191. [Google Scholar] [CrossRef]
- Luecke, D.M.; Kopp, A. Sex-specific evolution of relative leg size in Drosophila prolongata results from changes in the intersegmental coordination of tissue growth. Evolution 2019, 73, 2281–2294. [Google Scholar] [CrossRef]
- Oberlander, H. The imaginal disks. In Comprehensive Insect Physiology, Biochemistry, and Pharmacology, 1st ed.; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon: London, UK, 1985; pp. 151–182. [Google Scholar]
- Shingleton, A.W.; Frankino, W.A.; Flatt, T.; Nijhout, H.F.; Emlen, D.J. Size and shape: The developmental regulation of static allometry in insects. Bioessays 2007, 29, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Rolff, J.; Johnston, P.R.; Reynolds, S. Complete metamorphosis of insects. Philos. Trans. R. Soc. B 2019, 374, 20190063. [Google Scholar] [CrossRef] [PubMed]
- Mirth, C.K.; Saunders, T.E.; Amourda, C. Growing up in a changing world: Environmental regulation of development in insects. Annu. Rev. Entomol. 2021, 66, 81–99. [Google Scholar] [CrossRef]
- Norris, M.J.; Richards, O.W. Contributions towards the study of insect fertility. The structure and operation of the reproductive organs of the genera Ephestia and Plodia (Lepidoptera, Phycitidzae). Proc. Zool. Soc. Lond. 1932, 102, 595–612. [Google Scholar] [CrossRef]
- Esfandi, K.; He, X.Z.; Wang, Q. Flirtation reduces males’ fecundity but not longevity. Evolution 2015, 69, 2118–2128. [Google Scholar] [CrossRef]
- Brindley, T.A. The growth and development of Ephestia kuehniella Zeller (Lepidoptera) and Tribolium confusum Duval (Coleoptera) under controlled conditions of temperature and relative humidity. Ann. Entomol. Soc. Am. 1930, 23, 741–757. [Google Scholar] [CrossRef]
- Jacob, T.A.; Cox, P.D. The influence of temperature and humidity on the life-cycle of Ephestia kuehniella Zeller (Lepidoptera: Pyralidae). J. Stored. Prod. Res. 1977, 13, 107–118. [Google Scholar] [CrossRef]
- Friedländer, M.; Seth, R.K.; Reynolds, S.E. Eupyrene and apyrene sperm: Dichotomous spermatogenesis in Lepidoptera. In Advances in Insect Physiology; Simpson, S.J., Ed.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 206–308. [Google Scholar]
- Garbini, C.P.; Imberski, R.B. Spermatogenesis in Ephestia kuehniella (Lepidoptera, Pyralididae). Trans. Am. Microsc. Soc. 1977, 96, 189–203. [Google Scholar] [CrossRef]
- Cook, P.A.; Wedell, N. Non-fertile sperm delay female remating. Nature 1999, 397, 486. [Google Scholar] [CrossRef]
- Wedell, N.; Wiklund, C.; Bergstrom, J. Coevolution of non-fertile sperm and female receptivity in a butterfly. Biol. Lett. 2009, 5, 678–681. [Google Scholar] [CrossRef] [PubMed]
- Holman, L.; Snook, R.R. A sterile sperm caste protects brother fertile sperm from female-mediated death in Drosophila pseudoobscura. Curr. Biol. 2008, 18, 292–296. [Google Scholar] [CrossRef]
- Sakai, H.; Oshima, H.; Yuri, K.; Gotoh, H.; Daimon, T.; Yaginuma, T.; Sahara, K.; Niimi, T. Dimorphic sperm formation by Sex-lethal. Proc. Natl. Acad. Sci. USA 2019, 116, 10412–10417. [Google Scholar] [CrossRef]
- Konagaya, T.; Idogawa, N.; Watanabe, M. Destination of apyrene sperm following migration from the bursa copulatrix in the monandrous swallowtail butterfly Byasa alcinous. Sci. Rep. 2020, 10, 20907. [Google Scholar] [CrossRef] [PubMed]
- Hague, N.L.; Dickinson, J.L.; Shepherd, J.G. Transfer, subsequent movement, and fate of sperm in the tobacco hornworm moth, Manduca sexta. Physiol. Entomol. 2021. [Google Scholar] [CrossRef]
- Gage, M.J.G.; Cook, P.A. Sperm size or numbers? Effects of nutritional stress upon eupyrene and apyrene sperm production strategies in the moth Plodia interpunctella (Lepidoptera: Pyralidea). Funct. Ecol. 1994, 8, 594–599. [Google Scholar] [CrossRef]
- Sait, S.M.; Gage, M.J.G.; Cook, P.A. Effects of a fertility-reducing baculovirus on sperm numbers and sizes in the Indian meal moth, Plodia interpunctella. Funct. Ecol. 1998, 12, 56–62. [Google Scholar] [CrossRef]
- Koudelová, J.; Cook, P.A. Effect of gamma radiation and sex linked recessive lethal mutations on sperm transfer in Ephestia kuehniella (Lepidoptera: Pyralidae). Fla. Entomol. 2001, 84, 172–182. [Google Scholar] [CrossRef]
- Raichoudhury, D.P. Retardation of spermatogenesis and reduction of motility of sperm in Ephestia kühniella Z. (Lepidoptera, Phycitidae), caused by high temperature. Proc. Zool. Soc. Lond. 1936, 106, 789–805. [Google Scholar] [CrossRef]
- Millar, R.B.; Anderson, M.J. Remedies for pseudoreplication. Fish. Res. 2004, 70, 397–407. [Google Scholar] [CrossRef]
- Harrison, X.; Donaldson, L.; Correa, M.; Evans, J.; Fisher, D.; Goodwin, C.; Robinson, B.; Hodgson, D.; Inger, R. A brief introduction to mixed effects modelling and multi-model inference in ecology. PeerJ 2018, 6, e4794. [Google Scholar] [CrossRef]
- Davies, G.M.; Gray, A. Don’t let spurious accusations of pseudoreplication limit our ability to learn from natural experiments (and other messy kinds of ecological monitoring). Ecol. Evol. 2015, 5, 5295–5304. [Google Scholar] [CrossRef]
- Amiri, A.; Bandani, A.R. Parents’ living conditions influence offspring fitness and competency. J. Stored Prod. Res. 2021, 92, 101795. [Google Scholar] [CrossRef]
- Allen, L.E.; Barry, K.L.; Holwell, G.I.; Herberstein, M.E. Perceived risk of sperm competition affects juvenile development and ejaculate expenditure in male praying mantids. Anim. Behav. 2011, 82, 1201–1206. [Google Scholar] [CrossRef]
- Kasumovic, M.M.; Brooks, R.C. It’s all who you know: The evolution of socially cued anticipatory plasticity as a mating strategy. Q. Rev. Biol. 2011, 86, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Gray, B.; Simmons, L.W. Acoustic cues alter perceived sperm competition risk in the field cricket Teleogryllus oceanicus. Behav. Ecol. 2013, 24, 982–986. [Google Scholar] [CrossRef]
- Silberglied, R.E.; Shepherd, J.G.; Dickinson, J.L. Eunuchs: The role of apyrene sperm in Lepidoptera? Am. Nat. 1984, 123, 255–265. [Google Scholar] [CrossRef]
- Swallow, J.G.; Wilkinson, G.S. The long and short of sperm polymorphisms in insects. Biol. Rev. 2002, 77, 153–182. [Google Scholar] [CrossRef]
- Corbet, S.A. Mandibular gland secretion of larvae of the flour moth, Anagasta kuehniella, contains an epideictic pheromone and elicits oviposition movements in a hymenopteran parasite. Nature 1971, 232, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Mudd, A. Further novel 2-acylcyclohexane-1,3-diones from lepidopteran larvae. J. Chem. Soc. Perkin Trans. 1 1983, 2161–2164. [Google Scholar] [CrossRef]
- Crump, D.; Silverstein, R.M.; Williams, H.J.; Fitzgerald, T.D. Identification of trail pheromone of larva of eastern tent caterpillar Malacosoma americanum (Lepidoptera: Lasiocampidae). J. Chem. Ecol. 1987, 13, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, T.D.; Underwood, D.L.A. Trail marking by the larva of the madrone butterfly Eucheira socialis and the role of the trail pheromone in communal foraging behavior. J. Insect Behav. 1998, 11, 247–263. [Google Scholar] [CrossRef]
- Flowers, R.W.; Costa, J.T. Larval communication and group foraging dynamics in the red-headed pine sawfly, Neodiprion lecontei (Fitch) (Hymenoptera: Symphyta: Diprionidae). Ann. Entomol. Soc. Am. 2003, 96, 336–343. [Google Scholar] [CrossRef]
- Fitzgerald, T.D.; Pescador-Rubio, A. Trail marking and abandonment of depleted feeding sites by the caterpillars of Eutachyptera psidii (Lepidoptera: Lasiocampidae). J. Insect Behav. 2011, 24, 380–392. [Google Scholar] [CrossRef]
- Kojima, W.; Ishikawa, Y.; Takanashi, T. Chemically mediated group formation in soil-dwelling larvae and pupae of the beetle Trypoxylus dichotomus. Naturwissenschaften 2014, 101, 687–695. [Google Scholar] [CrossRef]
- Dombrovski, M.; Poussard, L.; Moalem, K.; Kmecova, L.; Hogan, N.; Schott, E.; Vaccari, A.; Acton, S.; Condron, B. Cooperative behavior emerges among Drosophila larvae. Curr. Biol. 2017, 27, 2821–2826. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; He, X.Z.; Zheng, X.-L.; Zhang, Y.; Wang, Q. Pupal Cues Increase Sperm Production but Not Testis Size in an Insect. Insects 2021, 12, 679. https://doi.org/10.3390/insects12080679
Liu J, He XZ, Zheng X-L, Zhang Y, Wang Q. Pupal Cues Increase Sperm Production but Not Testis Size in an Insect. Insects. 2021; 12(8):679. https://doi.org/10.3390/insects12080679
Chicago/Turabian StyleLiu, Junyan, Xiong Z. He, Xia-Lin Zheng, Yujing Zhang, and Qiao Wang. 2021. "Pupal Cues Increase Sperm Production but Not Testis Size in an Insect" Insects 12, no. 8: 679. https://doi.org/10.3390/insects12080679
APA StyleLiu, J., He, X. Z., Zheng, X.-L., Zhang, Y., & Wang, Q. (2021). Pupal Cues Increase Sperm Production but Not Testis Size in an Insect. Insects, 12(8), 679. https://doi.org/10.3390/insects12080679