
Nesting, Sex Ratio and Natural Enemies of the Giant Resin Bee in Relation to Native Species in Europe



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Organism
2.2. Study Area and Sampling
2.3. Data Analysis
2.3.1. Offspring Traits and Parasitism
3. Results
3.1. Above-ground Nesting Community
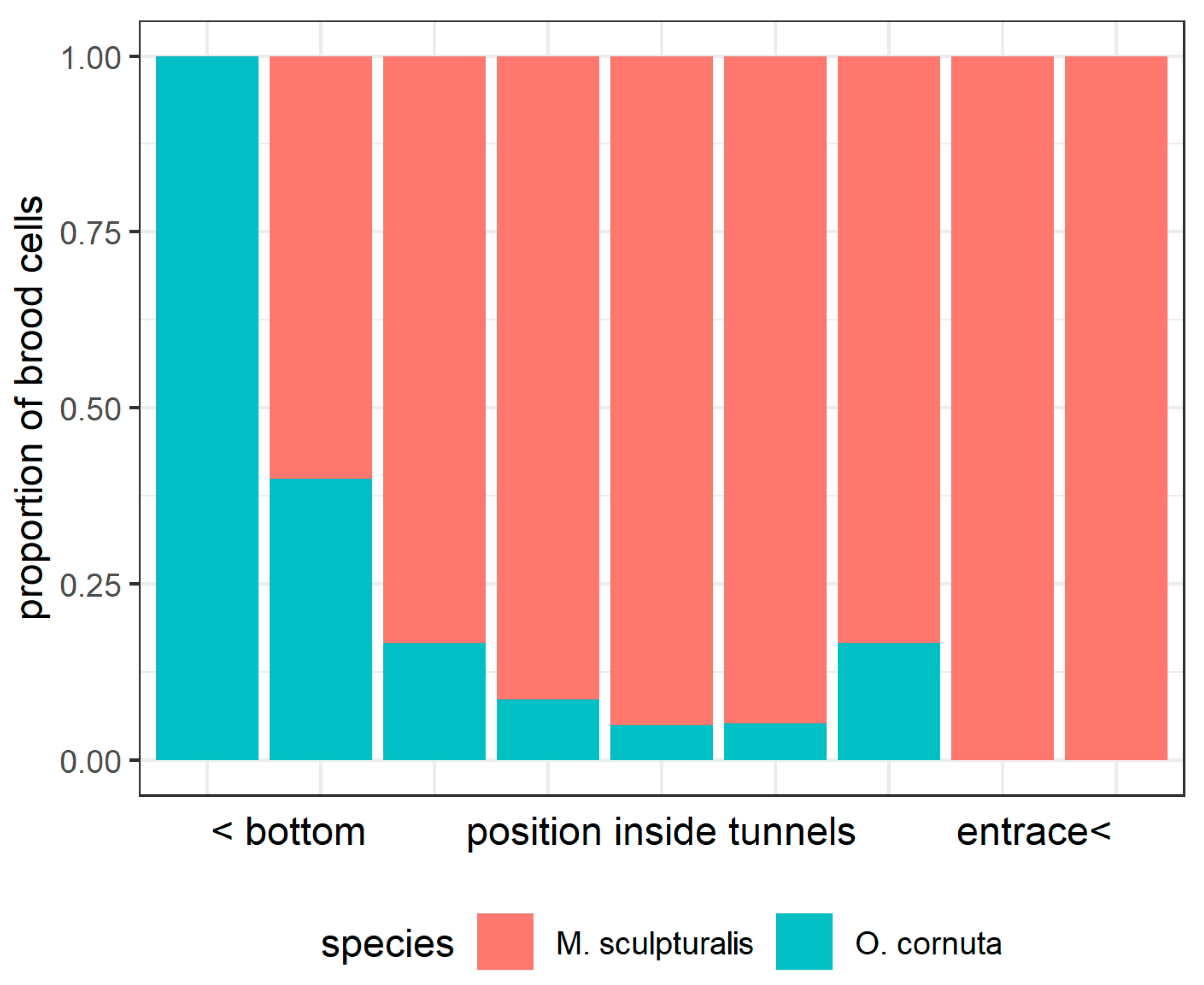
3.2. Interaction between the Exotic and the Native Bee
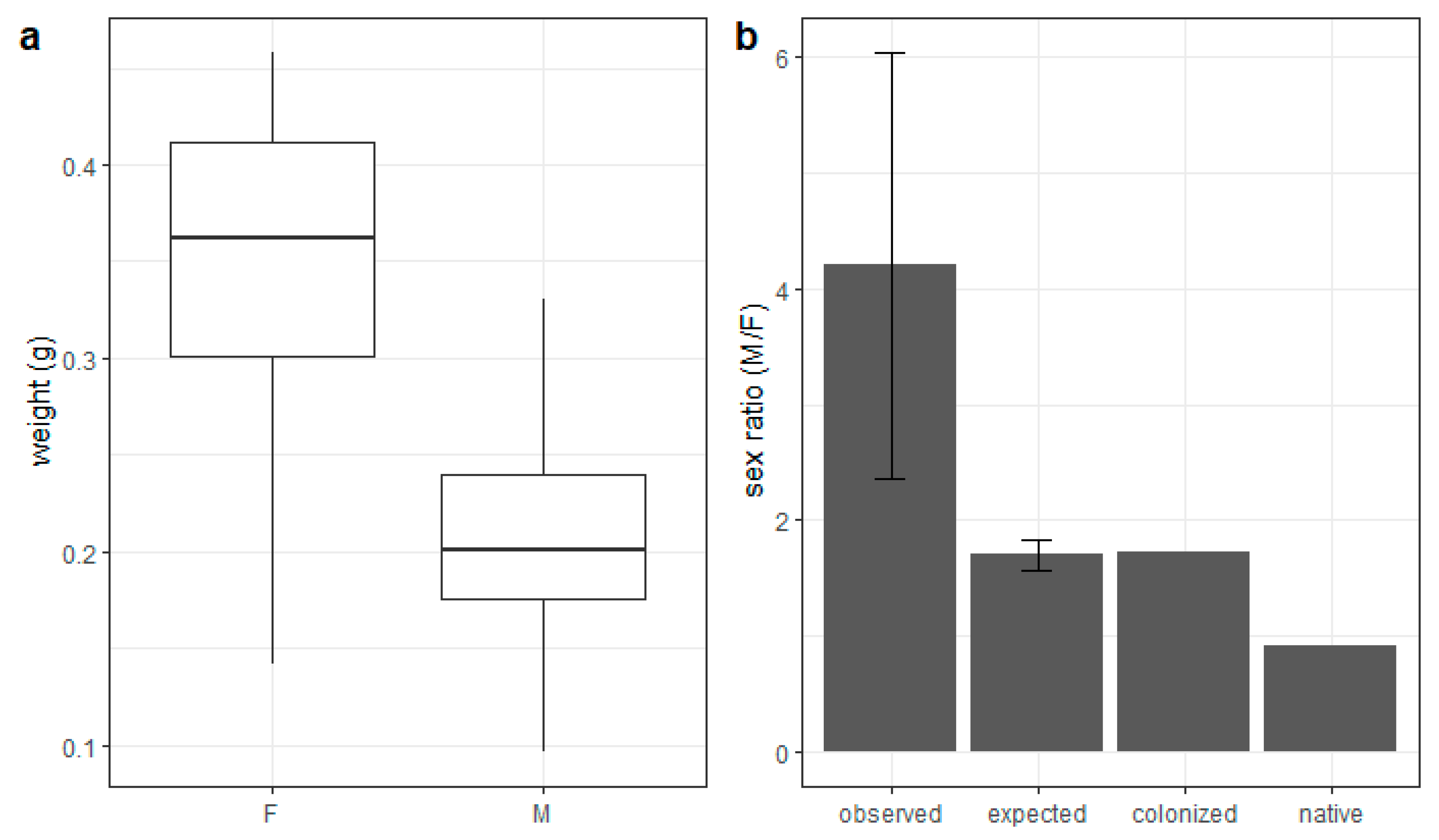
3.3. Progeny Weight and Sex Ratio of Exotic Bee
3.4. Natural Enemies
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nest | F | Mean Weight (g) | M | Mean Weight (g) | Observed Sex Ratio | Expected Sex Ratio |
|---|---|---|---|---|---|---|
| A | 11 | 0.353 | 46 | 0.205 | 4.2 | 1.7 |
| B | 10 | 0.352 | 39 | 0.215 | 3.9 | 1.6 |
| C | 15 | 0.341 | 53 | 0.199 | 3.5 | 1.7 |
| D | 4 | 0.370 | 30 | 0.199 | 7.5 | 1.9 |
| Total | 40 | 0.350 | 168 | 0.203 | 4.2 | 1.7 |
References
- Russo, L. Positive and Negative Impacts of Non-Native Bee Species around the World. Insects 2016, 7, 69. [Google Scholar] [CrossRef]
- Goulson, D. Effects of Introduced Bees on Native Ecosystems. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 1–26. [Google Scholar] [CrossRef]
- Rasmussen, C.; Carríon, A.L.; Castro-Urgal, R.; Chamorro, S.; Gonzalez, V.H.; Griswold, T.L.; Herrera, H.W.; McMullen, C.K.; Olesen, J.M.; Traveset, A. Megachile Timberlakei Cockerell (Hymenoptera: Megachilidae): Yet Another Adventive Bee Species to the Galápagos Archipelago. Pan-Pac. Entomol. 2012, 88, 98–102. [Google Scholar] [CrossRef]
- Vereecken, N.J.; Barbier, É. Premières Données Sur La Présence de l’abeille Asiatique Megachile (Callomegachile) Sculpturalis Smith (Hymenoptera, Megachilidae) En Europe. Osmia 2009, 3, 4–6. [Google Scholar] [CrossRef]
- Quaranta, M.; Sommaruga, A.; Balzarini, P.; Felicioli, A. A New Species for the Bee Fauna of Italy: Megachile Sculpturalis Continues Its Colonization of Europe. Bull. Insectol. 2014, 67, 287–293. [Google Scholar]
- Kovács, T. Megachile Sculpturalis Smith, 1853 in Hungary (Hymenoptera, Megachilidae). Folia Hist.-Nat. Musei Matra. 2015, 39, 73–76. [Google Scholar]
- Westrich, P. Megachile Sculpturalis Smith 1853 (Hymenoptera, Apidae), a New Species for the Bee Fauna of Germany, Now North of the Alps. Eucera 2015, 9, 3–10. [Google Scholar]
- Le Féon, V.; Aubert, M.; Genoud, D.; Andrieu-Ponel, V.; Westrich, P.; Geslin, B. Range Expansion of the Asian Native Giant Resin Bee Megachile Sculpturalis (Hymenoptera, Apoidea, Megachilidae) in France. Ecol. Evol. 2018, 8, 1534–1542. [Google Scholar] [CrossRef] [PubMed]
- Guariento, E.; Lanner, J.; Staggl, M.A.; Kranebitter, P. Megachile Sculpturalis (Hymenoptera: Megachilidae), the Giant Resin Bee New to South Tyrol. Gredleriana 2019, 19, 209–215. [Google Scholar] [CrossRef]
- Lanner, J.; Huchler, K.; Pachinger, B.; Sedivy, C.; Meimberg, H. Dispersal Patterns of an Introduced Wild Bee, Megachile Sculpturalis Smith, 1853 (Hymenoptera: Megachilidae) in European Alpine Countries. PLoS ONE 2020, 15, e0236042. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, S.P.; Fateryga, A.V. First Record of the Invasive Giant Resin Bee Megachile (Callomegachile) Sculpturalis Smith, 1853 (Hymenoptera: Megachilidae) in the Crimea. Far East. Entomol. 2019, 395, 7–13. [Google Scholar] [CrossRef]
- Aguado, O.; Hernández-Castellano, C.; Bassols, E.; Miralles, M.; Navarro, D.; Stefanescu, C.; Vicens, N. Megachile (Callomegachile) Sculpturalis Smith, 1853 (Apoidea: Megachilidae): A New Exotic Species in the Iberian Peninsula, and Some Notes about Its Biology. Butlletí Inst. Catalana D’Història Nat. 2018, 82, 157–162. [Google Scholar]
- Ortiz-Sánchez, F.J.N.; Varro, J.F.; Taeger, U. Megachile (Callomegachile) Sculpturalis Smith, 1853, Nueva. Bol. Soc. Entomológica Aragon. 2019, 63, 259–261. [Google Scholar]
- Polidori, C.; Sánchez-Fernández, D. Environmental Niche and Global Potential Distribution of the Giant Resin Bee Megachile Sculpturalis, a Rapidly Spreading Invasive Pollinator. Glob. Ecol. Conserv. 2020, 24, e01365. [Google Scholar] [CrossRef]
- Hinojosa-Díaz, I.I.; Yañez-Ordoñez, O.; Chen, G.; Peterson, A.T.; Engel, M.S. The North American Invasion of the Giant Resisn Bee. J. Hym. Res. 2005, 14, 69–77. [Google Scholar]
- Hinojosa-Díaz, I. The Giant Resin Bee Making Its Way West: First Record in Kansas (Hymenoptera: Megachilidae). ZooKeys 2008, 1, 67–71. [Google Scholar] [CrossRef]
- Parys, K.A.; Tripodi, A.D.; Sampson, B.J. The Giant Resin Bee, Megachile Sculpturalis Smith: New Distributional Records for the Mid- and Gulfsouth USA. Biodivers. Data J. 2015, 3. [Google Scholar] [CrossRef]
- Greenleaf, S.S.; Williams, N.M.; Winfree, R.; Kremen, C. Bee Foraging Ranges and Their Relationship to Body Size. Oecologia 2007, 153, 589–596. [Google Scholar] [CrossRef]
- Ruzzier, E.; Ruzzier, E.; Menchetti, M.; Bortolotti, L.; Selis, M.; Monterastelli, E. Updated Distribution of the Invasive Megachile Sculpturalis (Hymenoptera: Megachilidae) in Italy and Its First Record on a Mediterranean Island Updated Distribution of the Invasive Megachile Sculpturalis (Hymenoptera: Megachilidae) in Italy and Its. Biodivers. Data J. 2020, 8, e57783. [Google Scholar] [CrossRef] [PubMed]
- Andrieu-Ponel, V.; Ponel, P.; Le Féon, V.; Geslin, B.; Duvallet, G. A Propos Du Comportement de Butinage de Megachile Sculpturalis Smith, 1853, En France Méditerranéenne (Nîmes et Montpellier) (Hymenoptera, Megachilidae). Bull. Société Entomol. Fr. 2018, 123, 49–54. [Google Scholar]
- Geslin, B.; Gachet, S.; Deschamps-Cottin, M.; Flacher, F.; Ignace, B.; Knoploch, C.; Meineri, É.; Robles, C.; Ropars, L.; Schurr, L.; et al. Bee Hotels Host a High Abundance of Exotic Bees in an Urban Context. Acta Oecologica 2020, 105, 103556. [Google Scholar] [CrossRef]
- Rosas-Ramos, N.; Baños-Picón, L.; Tobajas, E.; Tormos, J.; Asís, J.D. Both Landscape and Local Scale Factors Matter for the Parental Investment Strategies of the Pollinator Osmia Caerulescens. J. Apic. Res. 2017, 56, 1–12. [Google Scholar] [CrossRef]
- Torchio, P.F.; Tepedino, V.J. Sex Ratio, Body Size and Seasonality in a Solitary Bee, Osmia Lignaria Propinqua Cresson (Hymenoptera: Megachilidae). Evolution 1980, 34, 993–1003. [Google Scholar] [CrossRef]
- Bosch, J.; Vicens, N. Sex Allocation in the Solitary Bee Osmia Cornuta: Do Females Behave in Agreement with Fisher’s Theory? Behav. Ecol. Sociobiol. 2005, 59, 124–132. [Google Scholar] [CrossRef]
- Ulbrich, K.; Seidelmann, K. Modeling Population Dynamics of Solitary Bees in Relation to Habitat Quality. Web Ecol. 2001, 2, 57–64. [Google Scholar] [CrossRef]
- Fitch, G.; Glaum, P.; Simao, M.C.; Vaidya, C.; Matthijs, J.; Iuliano, B.; Perfecto, I. Changes in Adult Sex Ratio in Wild Bee Communities Are Linked to Urbanization. Sci. Rep. 2019, 9, 3767. [Google Scholar] [CrossRef]
- Collet, M.; Vayssade, C.; Auguste, A.; Mouton, L.; Desouhant, E.; Malausa, T.; Fauvergue, X. Diploid Male Production Correlates with Genetic Diversity in the Parasitoid Wasp Venturia Canescens: A Genetic Approach with New Microsatellite Markers. Ecol. Evol. 2016, 6, 6721–6734. [Google Scholar] [CrossRef]
- Laport, R.G.; Minckley, R.L. Occupation of Active Xylocopa Virginica Nests by the Recently Invasive Megachile Sculpturalis in Upstate New York. J. Kans. Entomol. Soc. 2012, 85, 384–386. [Google Scholar] [CrossRef]
- Zandigiacomo, P.; Grion, M. First Finding of Megachile Sculpturalis Smith (Hymenoptera, Megachilidae) in Friuli Venezia Giulia (North-Eastern Italy). Gortania Bot. Zool. 2017, 39, 37–40. [Google Scholar]
- Dunn, A.M. Chapter 7 Parasites and Biological Invasions. In Advances in Parasitology: Natural History of Host-Parasite Interactions; Advances in Parasitology; Academic Press: Cambridge, MA, USA, 2009; Volume 68, pp. 161–184. [Google Scholar]
- Parker, J.D.; Torchin, M.E.; Hufbauer, R.A.; Lemoine, N.P.; Alba, C.; Blumenthal, D.M.; Bossdorf, O.; Byers, J.E.; Dunn, A.M.; Heckman, R.W.; et al. Do Invasive Species Perform Better in Their New Ranges? Ecology 2013, 94, 985–994. [Google Scholar] [CrossRef] [PubMed]
- Michener, C.D. The Bees of the World, 2nd ed.; The Johns Hopkins University Press: Baltimore, MD, USA, 2007. [Google Scholar]
- Bucci, A.; Barbero, D.; Lasagna, M.; Forno, M.G.; De Luca, D.A. Shallow Groundwater Temperature in the Turin Area (NW Italy): Vertical Distribution and Anthropogenic Effects. Environ. Earth Sci. 2017, 76, 221. [Google Scholar] [CrossRef]
- Acquaotta, F.; Fratianni, S.; Venema, V. Assessment of Parallel Precipitation Measurements Networks in Piedmont, Italy. Int. J. Climatol. 2016, 36, 3963–3974. [Google Scholar] [CrossRef]
- Bosch, J.; Kemp, W.P. Developing and Establishing Bee, Species as Crop Pollinators: The Example of Osmia Spp. (Hymenoptera: Megachilidae) and Fruit Trees. Bull. Entomol. Res. 2002, 92, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Seidelmann, K.; Bienasch, A.; Pröhl, F. The Impact of Nest Tube Dimensions on Reproduction Parameters in a Cavity Nesting Solitary Bee, Osmia Bicornis (Hymenoptera: Megachilidae). Apidologie 2016, 47, 114–122. [Google Scholar] [CrossRef]
- Fliszkiewicz, M.; Kuśnierczak, A.; Szymaś, B. The Accompanying Fauna of Solitary Bee Osmia Bicornis (L.) Syn. Osmia Rufa (L.) Nests Settled in Different Biotopes. J. Apic. Sci. 2012, 56, 51–58. [Google Scholar] [CrossRef]
- Krunić, M.; Stanisavljević, L.; Pinzauti, M.; Felicioli, A. The Accompanying Fauna of Osmia Cornuta and Osmia Rufa and Effective Measures of Protection. Bull. Insectology 2005, 58, 141–152. [Google Scholar]
- Zajdel, B.; Kucharska, K.; Kucharski, D.; Fliszkiewicz, M.; Gabka, J. Accompanying Fauna of Red Mason Bees (Osmia Bicornis L.) in Annual and Perennial Nesting Sites. Med. Weter. 2014, 70, 746. [Google Scholar]
- Majka, C.G.; Philips, T.K.; Sheffield, C. Ptinus Sexpunctatus Panzer (Coleoptera: Anobiidae, Ptininae) Newly Recorded in North America. Entomol. News 2007, 118, 73–76. [Google Scholar] [CrossRef]
- Seidelmann, K. Open-Cell Parasitism Shapes Maternal Investment Patterns in the Red Mason Bee Osmia Rufa. Behav. Ecol. 2006, 17, 839–848. [Google Scholar] [CrossRef]
- Fisher, R.A. The Genetical Theory of Natural Selectiontle; Oxford University Press: Oxford, UK, 1930. [Google Scholar]
- Global Biodiversity Information Facility. Free and Open Access to Biodiversity Data. GBIF Occurrence Download. Available online: https://www.gbif.org/occurrence/download/0176594-200613084148143 (accessed on 22 February 2021).
- Bates, A.J.; Sadler, J.P.; Fairbrass, A.J.; Falk, S.J.; Hale, J.D.; Matthews, T.J. Changing Bee and Hoverfly Pollinator Assemblages along an Urban-Rural Gradient. PLoS ONE 2011, 6, e23459. [Google Scholar] [CrossRef]
- Le Féon, V.; Geslin, B. Écologie et Distribution de l’Abeille Originaire d’Asie Megachile Sculpturalis S MITH 1853 (Apoidea—Megachilidae—Megachilini): Un État Des Connaissances Dix Ans Après Sa Première Observation En Europe. Osmia 2018, 7, 31–39. [Google Scholar] [CrossRef]
- Roulston, T.; Malfi, R. Aggressive Eviction of the Eastern Carpenter Bee (Xylocopa Virginica (Linnaeus)) from Its Nest by the Giant Resin Bee (Megachile Sculpturalis Smith). J. Kans. Entomol. Soc. 2002, 85, 387–388. [Google Scholar] [CrossRef]
- Quaranta, M.; Ambroselli, S.; Barro, P.; Bella, S.; Carini, A.; Celli, G.; Cogoi, P.; Comba, L.; Comoli, R.; Felicioli, A.; et al. Wild Bees in Agroecosystems and Semi-Natural Landscapes. 1997–2000 Collection Period in Italy. Bull. Insectology 2004, 57, 11–61. [Google Scholar]
- Lanner, J.; Gstöttenmayer, F.; Curto, M.; Geslin, B.; Huchler, K.; Orr, M.C.; Pachinger, B.; Sedivy, C.; Meimberg, H. Evidence for Multiple Introductions of an Invasive Wild Bee Species Currently under Rapid Range Expansion in Europe. BMC Ecol. Evol. 2021, 21, 17. [Google Scholar] [CrossRef]
- Gloag, R.; Ding, G.; Christie, J.R.; Buchmann, G.; Beekman, M.; Oldroyd, B.P. An Invasive Social Insect Overcomes Genetic Load at the Sex Locus. Nat. Ecol. Evol. 2016, 1, 11. [Google Scholar] [CrossRef]
- Darrouzet, E.; Gévar, J.; Guignard, Q.; Aron, S. Production of Early Diploid Males by European Colonies of the Invasive Hornet Vespa Velutina Nigrithorax. PLoS ONE 2015, 10, e0136680. [Google Scholar] [CrossRef] [PubMed]
- Russo, L.; de Keyzer, C.W.; Harmon-Threatt, A.N.; LeCroy, K.A.; MacIvor, J.S. The Managed-to-Invasive Species Continuum in Social and Solitary Bees and Impacts on Native Bee Conservation. Curr. Opin. Insect Sci. 2021. [Google Scholar] [CrossRef]
- Tepedino, V.J.; Parker, F.D. Alternation of Sex Ratio in a Partially Bivoltine Bee, Megachile Rotundata (Hymenoptera: Megachilidae). Ann. Entomol. Soc. Am. 1988, 81, 467–476. [Google Scholar] [CrossRef]
- Jones, C.M.; Brown, M.J.F. Parasites and Genetic Diversity in an Invasive Bumblebee. J. Anim. Ecol. 2014, 83, 1428–1440. [Google Scholar] [CrossRef]
- de Jong, Y.; Verbeek, M.; Michelsen, V.; de Place Bjørn, P.; Los, W.; Steeman, F.; Bailly, N.; Basire, C.; Chylarecki, P.; Stloukal, E.; et al. Fauna Europaea—All European Animal Species on the Web. Biodivers. Data J. 2014, 2, e4034. [Google Scholar] [CrossRef]




| Species | Nests (Occupied Tunnels) | Total Brood Cells | Mean Number of Cells per Nest | Maximum Number of Cells per Nest * | % of Parasitized Brood Cells | Emerged Adults | Natural Enemies |
|---|---|---|---|---|---|---|---|
| Osmia cornuta | 171 | 1013 | 5.9 | 15 | 93 | 12 | Cacoxenus indagator Chaetodactylus osmiae Melittobia acasta Monodontomerus obscurus Ptinus sexpunctatus |
| Megachile sculpturalis | 58 | 244 | 4.9 | 7 | 7 | 213 | Cacoxenus indagator Monodontomerus obscurus Ptinus sexpunctatus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Straffon-Díaz, S.; Carisio, L.; Manino, A.; Biella, P.; Porporato, M. Nesting, Sex Ratio and Natural Enemies of the Giant Resin Bee in Relation to Native Species in Europe. Insects 2021, 12, 545. https://doi.org/10.3390/insects12060545
Straffon-Díaz S, Carisio L, Manino A, Biella P, Porporato M. Nesting, Sex Ratio and Natural Enemies of the Giant Resin Bee in Relation to Native Species in Europe. Insects. 2021; 12(6):545. https://doi.org/10.3390/insects12060545
Chicago/Turabian StyleStraffon-Díaz, Sara, Luca Carisio, Aulo Manino, Paolo Biella, and Marco Porporato. 2021. "Nesting, Sex Ratio and Natural Enemies of the Giant Resin Bee in Relation to Native Species in Europe" Insects 12, no. 6: 545. https://doi.org/10.3390/insects12060545
APA StyleStraffon-Díaz, S., Carisio, L., Manino, A., Biella, P., & Porporato, M. (2021). Nesting, Sex Ratio and Natural Enemies of the Giant Resin Bee in Relation to Native Species in Europe. Insects, 12(6), 545. https://doi.org/10.3390/insects12060545