
Cuticle Hydrocarbons Show Plastic Variation under Desiccation in Saline Aquatic Beetles



Abstract
Simple Summary
Abstract

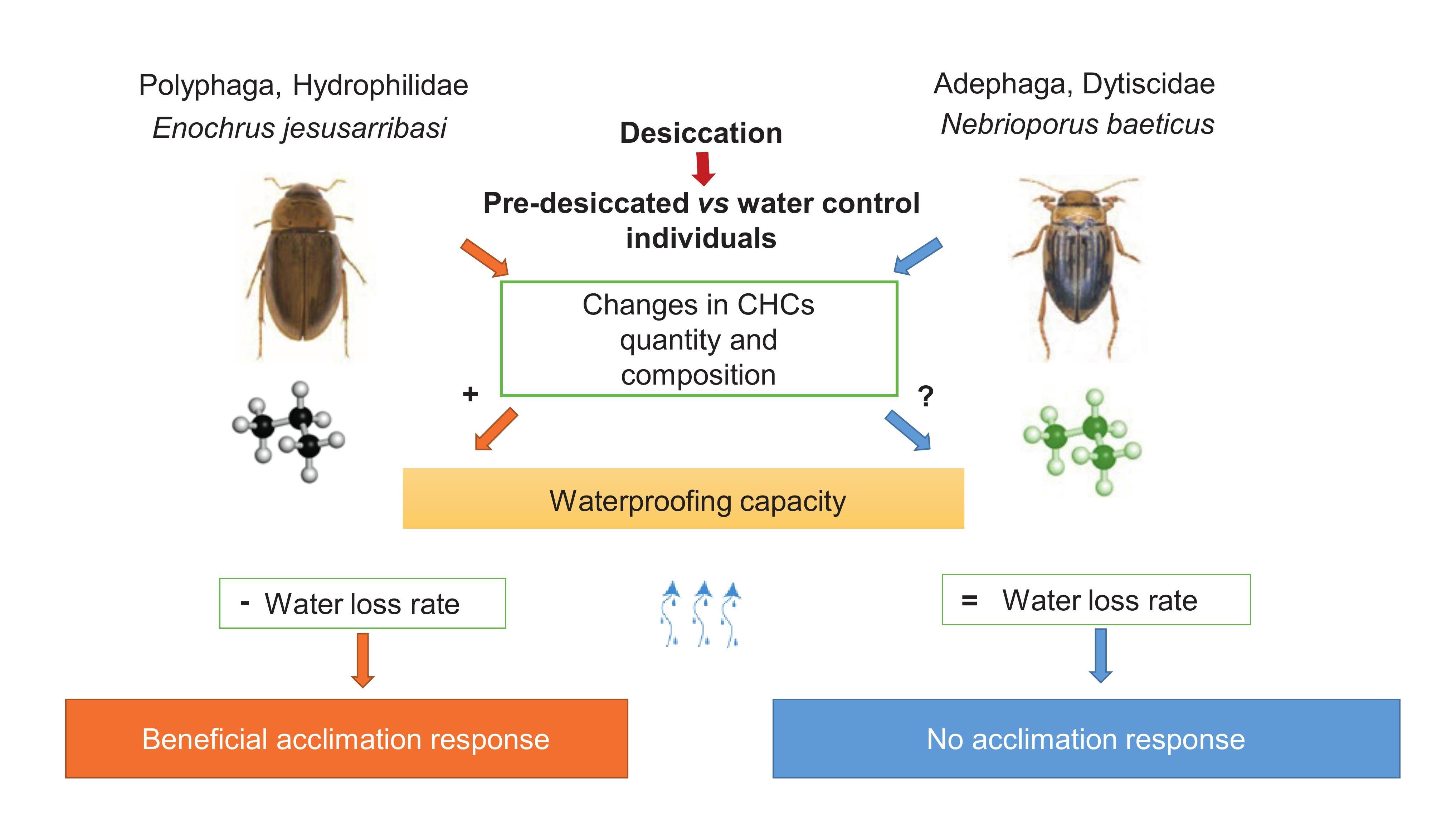
1. Introduction
2. Materials and Methods
2.1. Study Species, Field Collection and Maintenance
2.2. Desiccation Experimental Procedures
2.3. CHC Extraction
2.4. Cuticular Water Loss
2.5. Data Analysis
3. Results
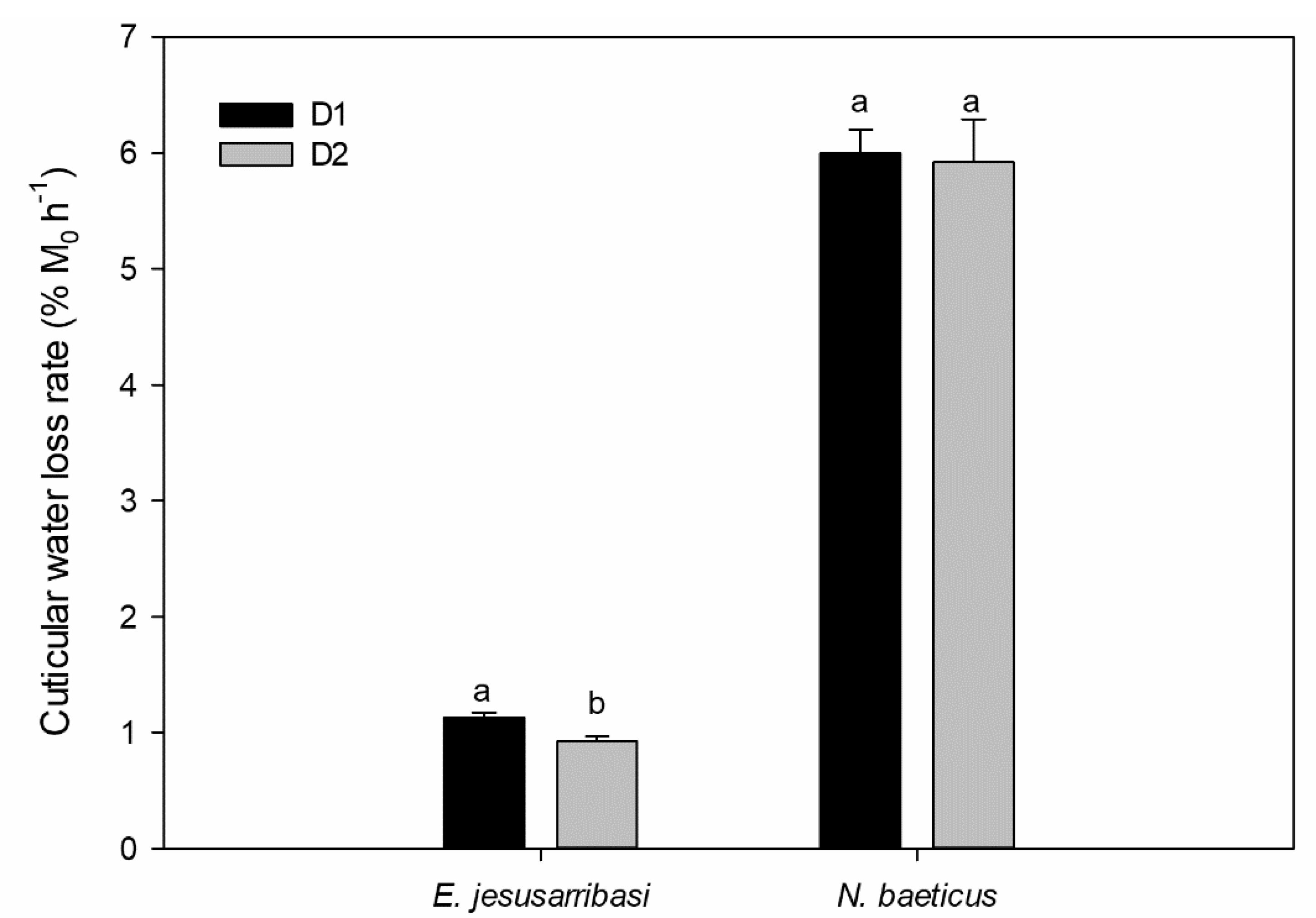
3.1. Variation in Cuticular Water Loss Rates
3.2. CHC Traits
3.3. CHC Profiles
4. Discussion
5. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chown, S.L.; Sørensen, J.G.; Terblanche, J.S. Water loss in insects: An environmental change perspective. J. Insect Physiol. 2011, 57, 1070–1084. [Google Scholar] [CrossRef] [PubMed]
- Bazinet, A.L.; Marshall, K.E.; MacMillan, H.A.; Williams, C.M.; Sinclair, B.J. Rapid changes in desiccation resistance in Drosophila melanogaster are facilitated by changes in cuticular permeability. J. Insect Physiol. 2010, 56, 2006–2012. [Google Scholar] [CrossRef] [PubMed]
- Reidenbach, K.R.; Cheng, C.; Liu, F.; Liu, C.; Besansky, N.J.; Syed, Z. Cuticular differences associated with aridity acclimation in African malaria vectors carrying alternative arrangements of inversion 2La. Parasites Vectors 2014, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Stinziano, J.R.; Sové, R.J.; Rundle, H.D.; Sinclair, B.J. Rapid desiccation hardening changes the cuticular hydrocarbon profile of drosophila melanogaster. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 180, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Menzel, F.; Zumbusch, M.; Feldmeyer, B. How ants acclimate: Impact of climatic conditions on the cuticular hydrocarbon profile. Funct. Ecol. 2018, 32, 657–666. [Google Scholar] [CrossRef]
- Bond, N.R.; Lake, P.S.; Arthington, A.H. The impacts of drought on freshwater ecosystems: An Australian perspective. Hydrobiologia 2008, 1–14. [Google Scholar] [CrossRef]
- Woodward, G.; Perkins, D.M.; Brown, L.E. Climate change and freshwater ecosystems: Impacts across multiple levels of organization. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2093–2106. [Google Scholar] [CrossRef] [PubMed]
- Velasco, J.; Millan, A.; Url, S. Insect Dispersal in a Drying Desert Stream: Effects of Temperature and Water Loss. Naturalist 1998, 43, 80–87. [Google Scholar]
- Dudley, R. The Biomechanics of Insect Flight: Form, Function, Evolution; Princeton University Press: Princeton, NJ, USA, 2000; ISBN 9780691094915. [Google Scholar]
- Strachan, S.R.; Chester, E.T.; Robson, B.J. Freshwater Invertebrate Life History Strategies for Surviving Desiccation. Springer Sci. Rev. 2015, 3, 57–75. [Google Scholar] [CrossRef]
- Chester, E.T.; Miller, A.D.; Valenzuela, I.; Wickson, S.J.; Robson, B.J. Drought survival strategies, dispersal potential and persistence of invertebrate species in an intermittent stream landscape. Freshw. Biol. 2015, 60, 2066–2083. [Google Scholar] [CrossRef]
- Cañedo-Argüelles, M.; Gutiérrez-Cánovas, C.; Acosta, R.; Castro-López, D.; Cid, N.; Fortuño, P.; Munné, A.; Múrria, C.; Pimentão, A.R.; Sarremejane, R.; et al. As time goes by: 20 years of changes in the aquatic macroinvertebrate metacommunity of Mediterranean river networks. J. Biogeogr. 2020, 47, 1861–1874. [Google Scholar] [CrossRef]
- Hering, D.; Haidekker, A.; Schmidt-Kloiber, A.; Barker, T.; Buisson, L.; Graf, W.; Grenouillet, G.; Lorenz, A.; Sandin, L.; Stendera, S. Monitoring the Responses of Freshwater Ecosystems to Climate Change. In Climate Change Impacts on Freshwater Ecosystems; Blackwell Publishing Ltd.: Oxford, UK, 2010; pp. 84–118. ISBN 9781405179133. [Google Scholar]
- Chown, S.L.; Nicolson, S. Insect Physiological Ecology: Mechanisms and Patterns; Oxford University Press: Oxford, UK, 2004; ISBN 9788578110796. [Google Scholar]
- Blomquist, G.J.; Bagnères, A.G. Insect Hydrocarbons Biology, Biochemistry, and Chemical Ecology; Cambridge University Press: Cambridge, UK, 2010; ISBN 9780511711909. [Google Scholar]
- Hadley, N. Water Relations of Terrestrial Arthropods; Academic Press: San Diego, CA, USA, 1994; ISBN 9780080918525. [Google Scholar]
- Gibbs, A.G. Lipid melting and cuticular permeability: New insights into an old problem. J. Insect Physiol. 2002, 48, 391–400. [Google Scholar] [CrossRef]
- Gibbs, A.; Pomonis, J.G. Physical properties of insect cuticular hydrocarbons: The effects of chain length, methyl-branching and unsaturation. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1995, 112, 243–249. [Google Scholar] [CrossRef]
- Benoit, J.B. Water Management by Dormant Insects: Comparisons between Dehydration Resistance During Summer Aestivation and Winter Diapause. In Aestivation Molecular and Physiological Aspects; Springer: New York, NY, USA, 2010; ISBN 9783642024207. [Google Scholar]
- Gibbs, A.G.; Rajpurohit, S. Cuticular lipids and water balance Cuticular lipids and water balance. Insect Hydrocarb. Biol. Biochem. Chem. Ecol. 2010, 100–120. [Google Scholar] [CrossRef]
- Rajpurohit, S.; Hanus, R.; Vrkoslav, V.; Behrman, E.L.; Bergland, A.O.; Petrov, D.; Cvačka, J.; Schmidt, P.S. Adaptive dynamics of cuticular hydrocarbons in Drosophila. J. Evol. Biol. 2017, 30, 66–80. [Google Scholar] [CrossRef] [PubMed]
- Sprenger, P.P.; Burkert, L.H.; Abou, B.; Federle, W.; Menzel, F. Coping with climate: Cuticular hydrocarbon acclimation of ants under constant and fluctuating conditions. J. Exp. Biol. 2018, 221, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Hadley, N. Epicuticular lipids of the desert tenebrionid beetle, Eleodes armata: Seasonal and acclimatory effects on composition. Insect Biochem. 1977, 7, 277–283. [Google Scholar] [CrossRef]
- Noorman, N.; Den Otter, C.J. Effects of relative humidity, temperature, and population density on production of cuticular hydrocarbons in housefly Musca domestica L. J. Chem. Ecol. 2002, 28, 1819–1829. [Google Scholar] [CrossRef]
- Gefen, E.; Talal, S.; Brendzel, O.; Dror, A.; Fishman, A. Variation in quantity and composition of cuticular hydrocarbons in the scorpion Buthus occitanus (Buthidae) in response to acute exposure to desiccation stress. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 182, 58–63. [Google Scholar] [CrossRef]
- Sprenger, P.P.; Menzel, F. Cuticular hydrocarbons in ants (Hymeno ptera: Formicidae) and other insects: How and why they differ among individuals, colonies, and species. Myrmecol. News 2020, 29, 1–26. [Google Scholar] [CrossRef]
- Hoffmann, A.A. Acclimation for desiccation resistance in Drosophila melanogaster and the association between acclimation responses and genetic variation. J. Insect Physiol. 1990, 36, 885–891. [Google Scholar] [CrossRef]
- Hoffmann, A.A. Acclimation for desiccation resistance in Drosophila: Species and population comparisons. J. Insect Physiol. 1991, 37, 757–762. [Google Scholar] [CrossRef]
- Aggarwal, D.D.; Ranga, P.; Kalra, B.; Parkash, R.; Rashkovetsky, E.; Bantis, L.E. Rapid effects of humidity acclimation on stress resistance in Drosophila melanogaster. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 166, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, B.J.; Ferguson, L.V.; Salehipour-Shirazi, G.; Macmillan, H.A. Cross-tolerance and cross-talk in the cold: Relating low temperatures to desiccation and immune stress in insects. Integr. Comp. Biol. 2013, 53, 545–556. [Google Scholar] [CrossRef]
- Pallarés, S.; Arribas, P.; Bilton, D.T.; Millán, A.; Velasco, J.; Ribera, I. The chicken or the egg? Adaptation to desiccation and salinity tolerance in a lineage of water beetles. Mol. Ecol. 2017, 26, 5614–5628. [Google Scholar] [CrossRef]
- Janzen, D.H. Why Mountain Passes are Higher in the Tropics. Am. Nat. 1967, 101, 233–249. [Google Scholar] [CrossRef]
- Stevens, G.C. The latitudinal gradient in geographical range: How so many species coexist in the tropics. Am. Nat. 1989, 133, 240–256. [Google Scholar] [CrossRef]
- Rohr, J.R.; Civitello, D.J.; Cohen, J.M.; Roznik, E.A.; Sinervo, B.; Dell, A.I. The complex drivers of thermal acclimation and breadth in ectotherms. Ecol. Lett. 2018, 21, 1425–1439. [Google Scholar] [CrossRef]
- Botella-Cruz, M.; Pallarés, S.; Millán, A.; Velasco, J. Role of cuticle hydrocarbons composition in the salinity tolerance of aquatic beetles. J. Insect Physiol. 2019, 117, 103899. [Google Scholar] [CrossRef]
- Arribas, P.; Andújar, C.; Abellán, P.; Velasco, J.; Millán, A.; Ribera, I. Tempo and mode of the multiple origins of salinity tolerance in a water beetle lineage. Mol. Ecol. 2014, 23, 360–373. [Google Scholar] [CrossRef]
- Pallarés, S.; Botella-Cruz, M.; Arribas, P.; Millán, A.; Velasco, J. Aquatic insects in a multistress environment: Cross-tolerance to salinity and desiccation. J. Exp. Biol. 2017, 220, 1277–1286. [Google Scholar] [CrossRef]
- Bilton, D.T.; Ribera, I.; Short, A.E.Z. Water Beetles as Models in Ecology and Evolution. Annu. Rev. Entomol. 2019, 64, 359–377. [Google Scholar] [CrossRef]
- Millán, A.; Sánchez-fernández, D.; Abellán, P.; Picazo, F.; Carbonell, J.A.; Lobo, J.M.; Ribera, I. Atlas de Los Coleópteros Acuáticos de España Peninsular; Ministerio de Agricultura, Alimentación y Medio Ambiente: Madrid, Spain, 2014; ISBN 9788449114182.
- Céspedes, V.; Pallarés, S.; Arribas, P.; Millán, A.; Velasco, J. Water beetle tolerance to salinity and anionic composition and its relationship to habitat occupancy. J. Insect Physiol. 2013, 59, 1076–1084. [Google Scholar] [CrossRef]
- Pallarés, S.; Arribas, P.; Bilton, D.T.; Millán, A.; Velasco, J. The comparative osmoregulatory ability of two water beetle genera whose species span the fresh-hypersaline gradient in inland waters (Coleoptera: Dytiscidae, Hydrophilidae). PLoS ONE 2015, 10, e0124299. [Google Scholar] [CrossRef] [PubMed]
- Yee, D.A. Ecology, Systematics, and the Natural History of Predaceous Diving Beetles (Coleoptera: Dytiscidae); Yee, D.A., Ed.; Springer: Dordrecht, The Netherlands, 2014; ISBN 9789401791083. [Google Scholar]
- Botella-Cruz, M.; Villastrigo, A.; Pallarés, S.; López-Gallego, E.; Millán, A.; Velasco, J. Cuticle hydrocarbons in saline aquatic beetles. PeerJ 2017, 5, e3562. [Google Scholar] [CrossRef] [PubMed]
- Schilman, P.E.; Lighton, J.R.B.; Holway, D.A. Respiratory and cuticular water loss in insects with continuous gas exchange: Comparison across five ant species. J. Insect Physiol. 2005, 51, 1295–1305. [Google Scholar] [CrossRef] [PubMed]
- Zuur, A.; Ieno, E.; Walker, N.; Saveliev, A.; Smith, G. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009; p. 574. ISBN 978-0-387-87457-9. [Google Scholar]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, 1–128. [Google Scholar] [CrossRef]
- Menzel, F.; Blaimer, B.B.; Schmitt, T. How do cuticular hydrocarbons evolve? Physiological constraints and climatic and biotic selection pressures act on a complex functional trait. Proc. R. Soc. B Biol. Sci. 2017, 284. [Google Scholar] [CrossRef]
- Crowson, R. The Biology of the Coleoptera; Academic Press: London, UK, 1981; ISBN 0121960501. [Google Scholar]
- Dulbecco, A.B.; Moriconi, D.E.; Lynn, S.; McCarthy, A.; Juárez, M.P.; Girotti, J.R.; Calderón-Fernández, G.M. Deciphering the role of Rhodnius prolixus CYP4G genes in straight and methyl-branched hydrocarbon formation and in desiccation tolerance. Insect Mol. Biol. 2020. [Google Scholar] [CrossRef]
- Engl, T.; Eberl, N.; Gorse, C.; Krüger, T.; Schmidt, T.H.P.; Plarre, R.; Adler, C.M. Kaltenpoth Ancient symbiosis confers desiccation resistance to stored grain pest beetles. Mol. Ecol. 2018, 27, 2095–2108. [Google Scholar] [CrossRef]
- Leeson, S.A.; Kennington, W.J.; Evans, T.A.; Simmons, L.W. Phenotypic plasticity but no adaptive divergence in cuticular hydrocarbons and desiccation resistance among translocated populations of dung beetles. Evol. Ecol. 2020. [Google Scholar] [CrossRef]
- Buellesbach, J.; Whyte, B.A.; Cash, E.; Gibson, J.D.; Scheckel, K.J.; Sandidge, R.; Tsutsui, N.D. Desiccation Resistance and Micro-Climate Adaptation: Cuticular Hydrocarbon Signatures of Different Argentine Ant Supercolonies Across California. J. Chem. Ecol. 2018, 44, 1101–1114. [Google Scholar] [CrossRef] [PubMed]
- Hunt, T.; Bergsten, J.; Levkanicova, Z.; Papadopoulou, A.; St. John, O.; Wild, R.; Hammond, P.M.; Ahrens, D.; Balke, M.; Caterino, M.S.; et al. A comprehensive phylogeny of beetles reveals the evolutionary origins of a superradiation. Science 2007, 318, 1913–1916. [Google Scholar] [CrossRef] [PubMed]
- Kellermann, V.; Hoffmann, A.A.; Overgaard, J.; Loeschcke, V.; Sgrò, C.M. Plasticity for desiccation tolerance across drosophila species is affected by phylogeny and climate in complex ways. Proc. R. Soc. B Biol. Sci. 2018, 285, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Feyereisen, R. Origin and evolution of the CYP4G subfamily in insects, cytochrome P450 enzymes involved in cuticular hydrocarbon synthesis. Mol. Phylogenet. Evol. 2020, 143. [Google Scholar] [CrossRef] [PubMed]
- Abellán, P.; Sánchez-Fernández, D.; Picazo, F.; Millán, A.; Lobo, J.M.; Ribera, I. Preserving the evolutionary history of freshwater biota in Iberian National Parks. Biol. Conserv. 2013, 162, 116–126. [Google Scholar] [CrossRef]
- Bernhard, D.; Schmidt, C.; Korte, A.; Fritzsch, G.; Beutel, R.G. From terrestrial to aquatic habitats and back again—Molecular insights into the evolution and phylogeny of Hydrophiloidea (Coleoptera) using multigene analyses. Zool. Scr. 2006, 35, 597–606. [Google Scholar] [CrossRef]
- Short, A.E.Z.; Fikáček, M. Molecular phylogeny, evolution and classification of the Hydrophilidae (Coleoptera). Syst. Entomol. 2013, 38, 723–752. [Google Scholar] [CrossRef]
- Bloom, D.D.; Fikáček, M.; Short, A.E.Z. Clade age and diversification rate variation explain disparity in species richness among water scavenger beetle (Hydrophilidae) lineages. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Shull, V.L.; Vogler, A.P.; Baker, M.D.; Maddison, D.R.; Hammond, P.M. Sequence alignment of 18S ribosomal RNA and the basal relationships of adephagan beetles: Evidence for monophyly of aquatic families and the placement of trachypachidae. Syst. Biol. 2001, 50, 945–969. [Google Scholar] [CrossRef]
- Gunderson, A.R.; Armstrong, E.J.; Stillman, J.H. Multiple Stressors in a Changing World: The Need for an Improved Perspective on Physiological Responses to the Dynamic Marine Environment. Ann. Rev. Mar. Sci. 2016, 8, 357–378. [Google Scholar] [CrossRef] [PubMed]
- Bogert, C.M. Thermoregulation in reptiles; a factor in evolution. Evolution 1949, 3, 195–211. [Google Scholar] [CrossRef] [PubMed]
- Huey, R.B.; Hertz, P.E.; Sinervo, B. Behavioral drive versus behavioral inertia in evolution: A null model approach. Am. Nat. 2003, 161, 357–366. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Treatment | Total n° CHC | Median CL ± SE | Alkanes | Unsaturated | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total Abundance (ng) | n-Alkanes | Branched Alkanes | |||||||||||
| n° | % | Median CL ± SE | n° | % | Median CL ± SE | n° | % | Median CL ± SE | |||||
| E. jesusarribasi | Control | 1315.92 ± 177.04 a | 42 | 25.62 ± 0.09 | 8 | 16.91 | 25.96 ± 0.15 a | 26 | 39.16 a | 26.06 ± 0.06 a | 8 | 43.93 a | 23.83 ± 0.25 |
| D1 | 2036.90 ± 339.58 b | 42 | 25.44 ± 0.16 | 9 | 17.12 | 26.23 ± 0.15 a | 25 | 35.45 a | 26.76 ± 0.25 a | 9 | 47.43 a | 23.33 ± 0.17 | |
| D2 | 1294.13 ± 171.13 a | 40 | 26.56 ± 0.20 | 7 | 21.52 | 27.97 ± 0.68 b | 26 | 56.65 b | 27.72 ± 0.11 b | 7 | 21.83 b | 23.98 ± 0.22 | |
| N. baeticus | Control | 7936.46 ± 1548.11 a | 48 | 24.70 ± 0.06 | 8 | 32.11 | 25.99 ±0.05 | 26 | 27.24 | 25.01 ± 0.12 | 14 | 40.65 | 23.21 ± 0.05 a |
| D1 | 6333.31 ± 379.04 b | 47 | 24.81 ± 0.03 | 8 | 37.91 | 25.96 ± 0.10 | 25 | 27.40 | 25.12 ± 0.09 | 12 | 34.69 | 23.35 ± 0.06 a | |
| D2 | 3505.50 ± 618.20 c | 41 | 24.66 ± 0.16 | 10 | 32.34 | 25.04 ± 0.16 | 21 | 25.97 | 25.10 ± 0.37 | 10 | 41.67 | 23.85 ± 0.29 b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Botella-Cruz, M.; Velasco, J.; Millán, A.; Hetz, S.; Pallarés, S. Cuticle Hydrocarbons Show Plastic Variation under Desiccation in Saline Aquatic Beetles. Insects 2021, 12, 285. https://doi.org/10.3390/insects12040285
Botella-Cruz M, Velasco J, Millán A, Hetz S, Pallarés S. Cuticle Hydrocarbons Show Plastic Variation under Desiccation in Saline Aquatic Beetles. Insects. 2021; 12(4):285. https://doi.org/10.3390/insects12040285
Chicago/Turabian StyleBotella-Cruz, María, Josefa Velasco, Andrés Millán, Stefan Hetz, and Susana Pallarés. 2021. "Cuticle Hydrocarbons Show Plastic Variation under Desiccation in Saline Aquatic Beetles" Insects 12, no. 4: 285. https://doi.org/10.3390/insects12040285
APA StyleBotella-Cruz, M., Velasco, J., Millán, A., Hetz, S., & Pallarés, S. (2021). Cuticle Hydrocarbons Show Plastic Variation under Desiccation in Saline Aquatic Beetles. Insects, 12(4), 285. https://doi.org/10.3390/insects12040285