Regulation of Juvenile Hormone on Summer Diapause of Geleruca daurica and Its Pathway Analysis

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. JHA Treatment
2.3. RNA Extraction and cDNA Synthesis
2.4. Transcriptome Sequencing, Assembly and Bioinformatics Analysis
2.5. Quantitative Real-Time PCR
2.6. Total Lipid Content Measurement
2.7. Behavioral and Developmental Observation
2.8. Statistical Analysis
3. Results
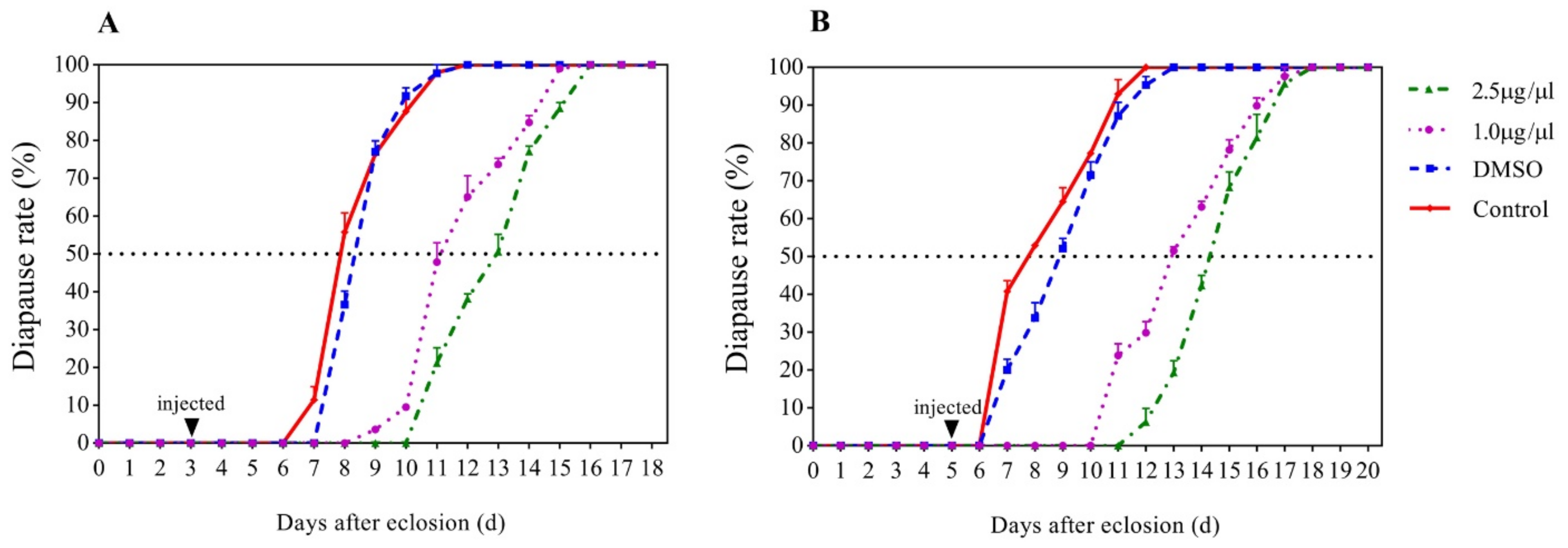
3.1. Effects of JHA on the Development of G. daurica Adults
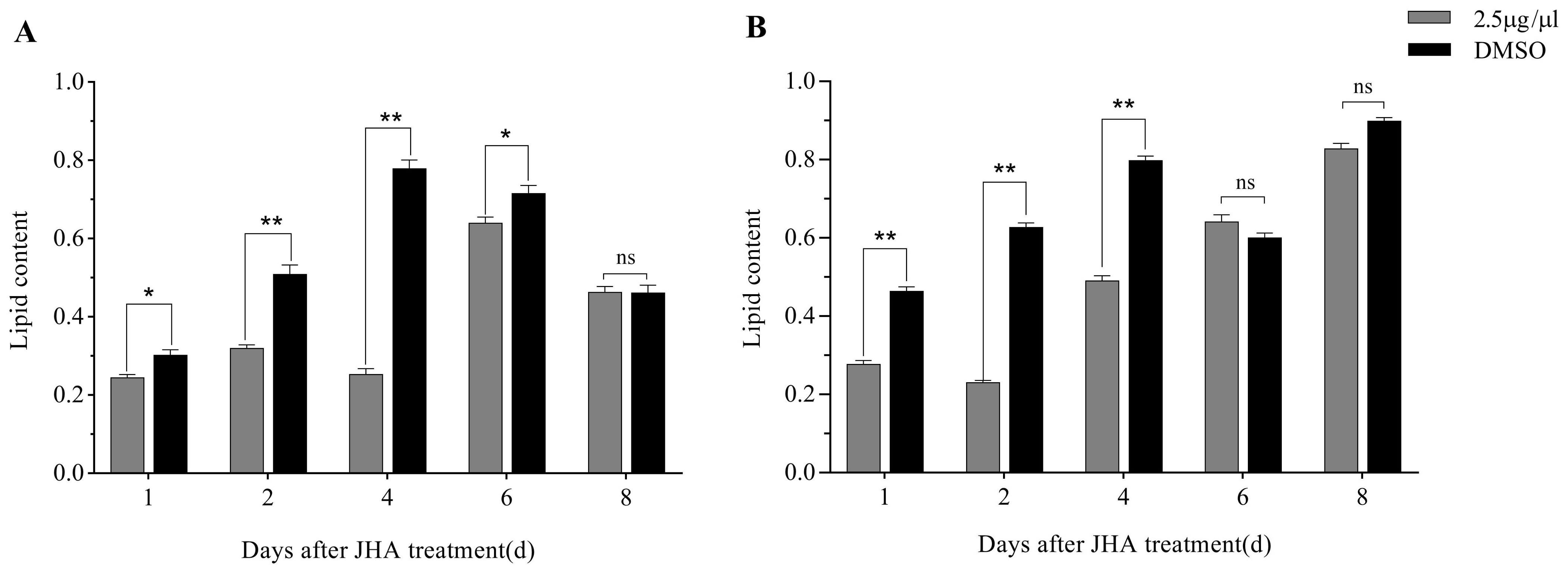
3.2. Effects of JHA on the Total Lipid Content in G. daurica
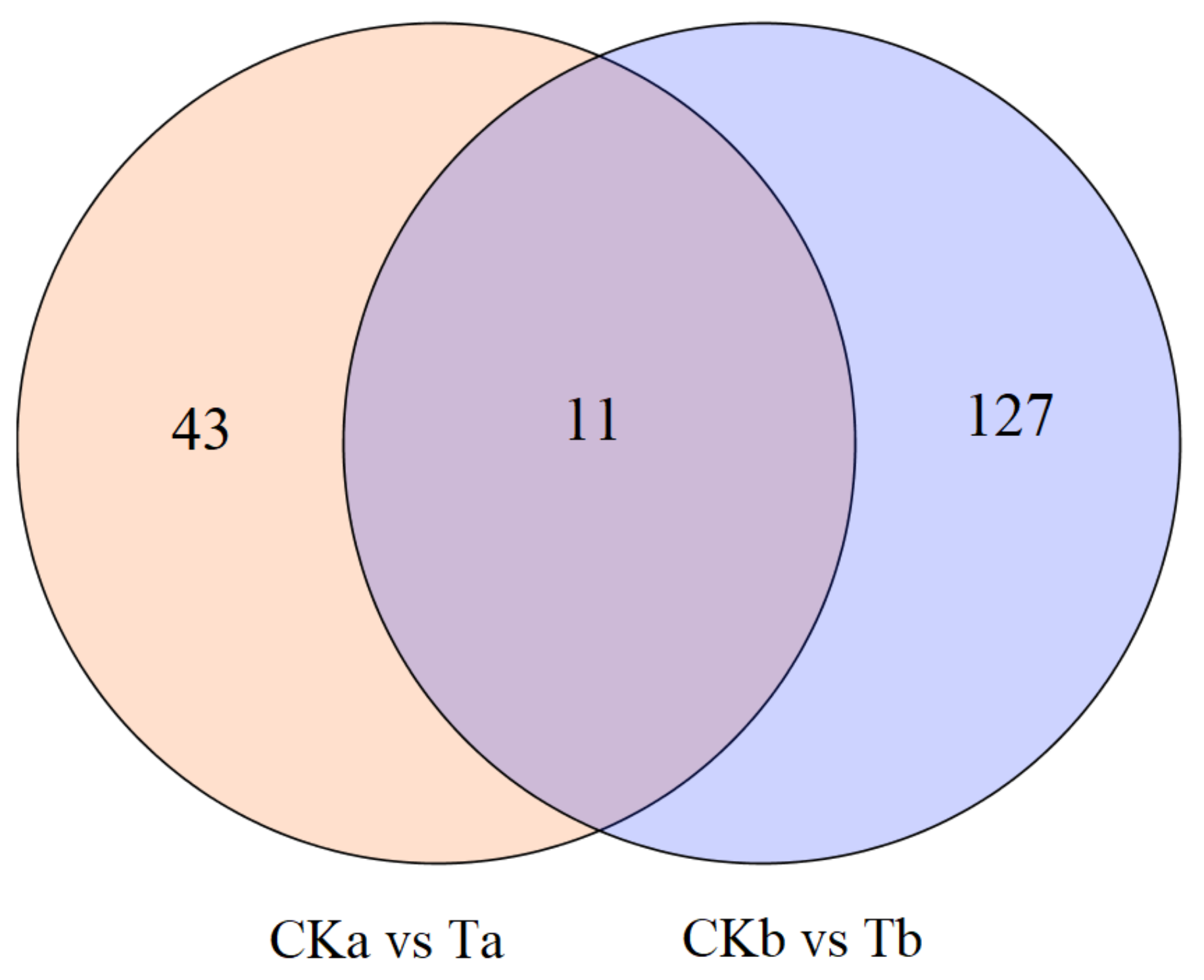
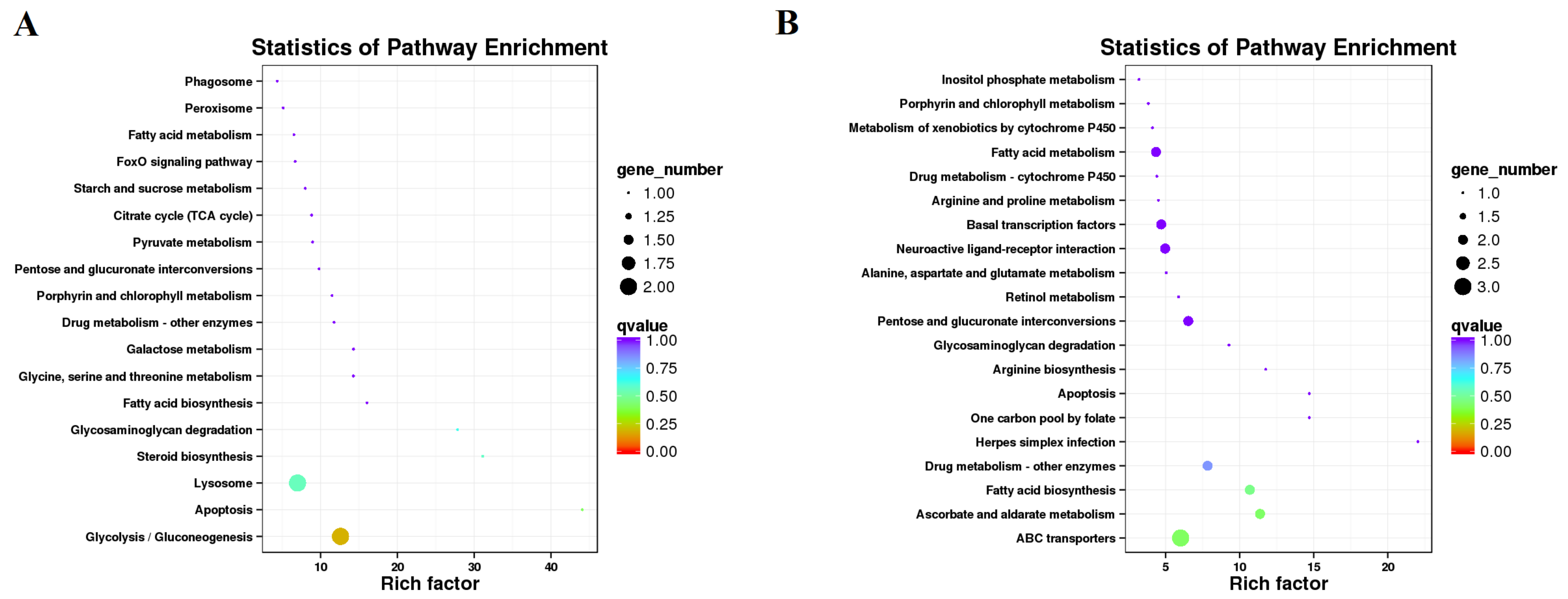
3.3. Transcriptome Changes in Response to JHA Application
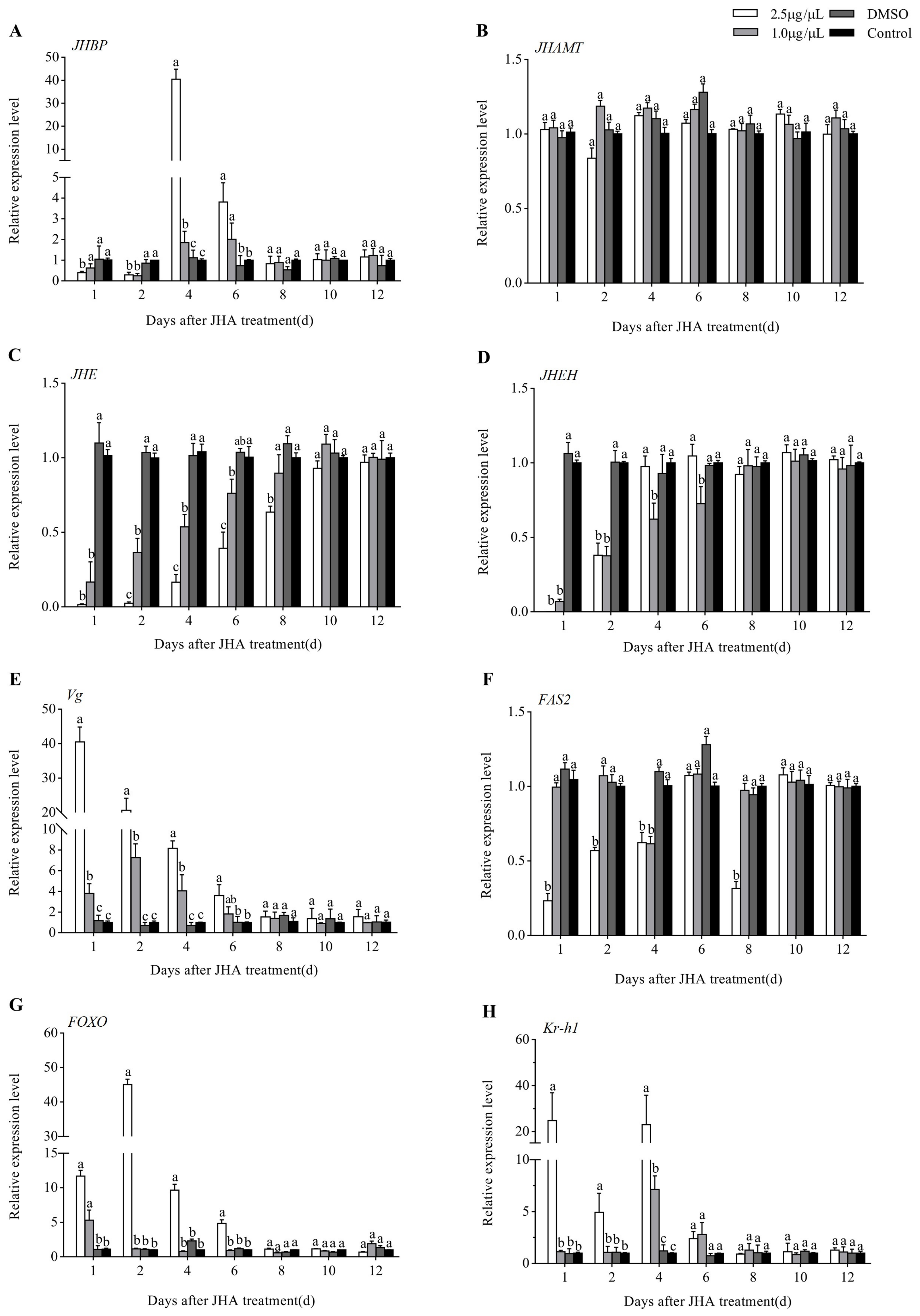
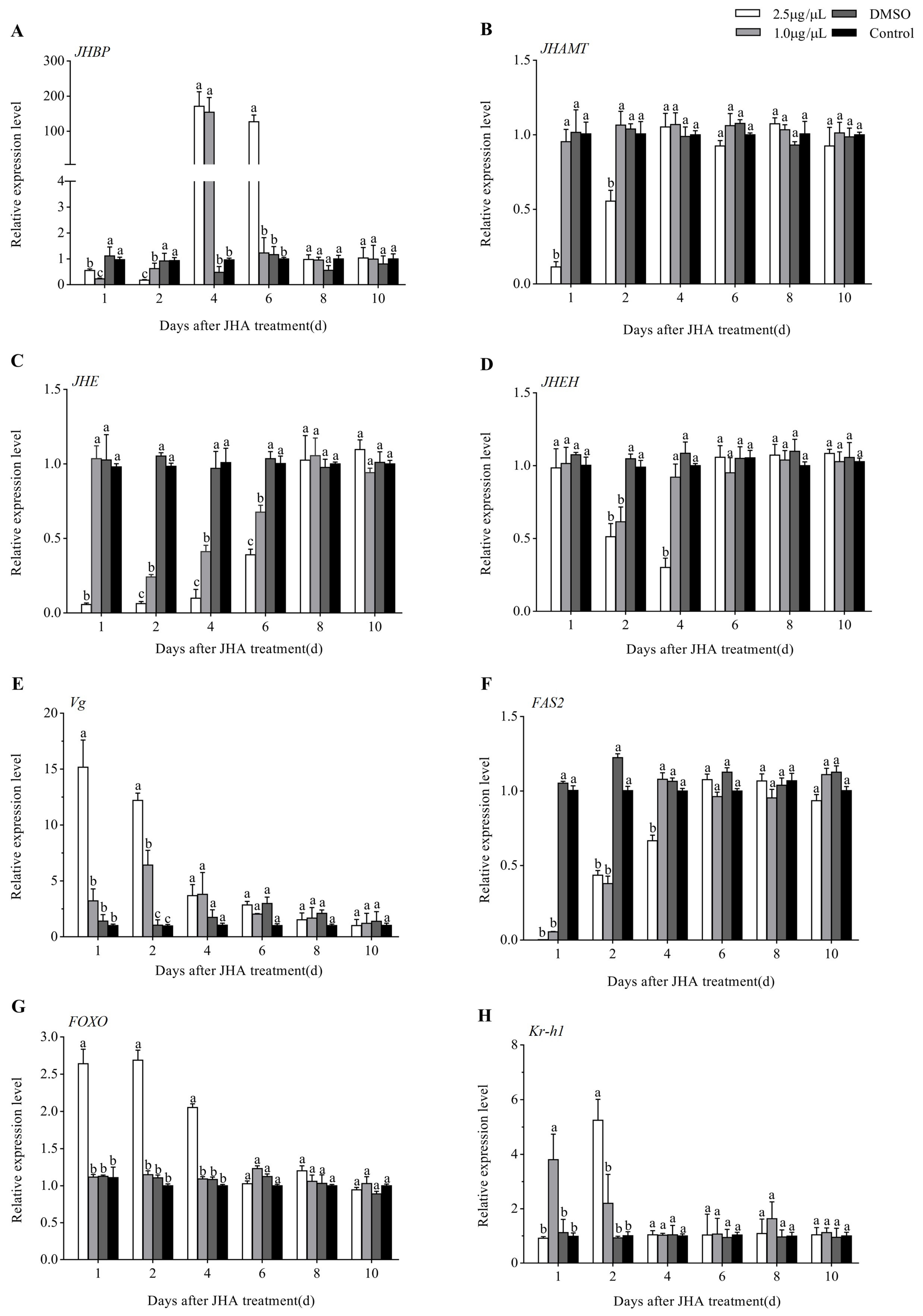
3.4. Effects of JHA Application on JH Signaling Pathway-Related Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kostál, V. Eco-physiological phases of insect diapause. J. Insect Physiol. 2006, 52, 113–127. [Google Scholar] [CrossRef]
- Denlinger, D.L. Regulation of diapause. Annu. Rev. Entomol. 2002, 47, 93–122. [Google Scholar] [CrossRef] [PubMed]
- Hodek, I. Adult diapause in Coleoptera. Psyche 2012, 1–10. [Google Scholar] [CrossRef]
- Denlinger, D.L.; Yocum, G.D.; Rinehart, J.P. Hormonal Control of Diapause. In Insect Endocrinology; Gilbert, L.I., Ed.; Elsevier: London, UK, 2012; pp. 430–463. [Google Scholar]
- Jindra, M.; Palli, S.R.; Riddiford, L.M. The juvenile hormone signaling pathway in insect development. Annu. Rev. Entomol. 2013, 58, 181–204. [Google Scholar] [CrossRef]
- Gujar, H.; Palli, S.R. Juvenile hormone regulation of metamorphosis in the common bed bug, Cimex lectularius. Sci. Rep. 2016, 6, 26092. [Google Scholar] [CrossRef] [PubMed]
- Ashok, M.; Turner, C.; Wilson, T.G. Insect juvenile hormone resistance gene homology with the bHLH-PAS family of transcriptional regulators. Proc. Natl. Acad. Sci. USA 1998, 95, 2761–2766. [Google Scholar] [CrossRef] [PubMed]
- Minakuchi, C.; Namiki, T.; Shinoda, T. Kruppel homolog 1 an early juvenile hormone-response gene downstream of Methoprene-tolerant, mediates its anti-metamorphic action in the red flour beetle Tribolium castaneum. Dev. Biol. 2009, 325, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Charles, J.P.; Iwema, T.; Epa, V.C.; Takaki, K.; Rynes, J.; Jindra, M. Ligand-binding properties of a juvenile hormone receptor, Methoprene-tolerant. Proc. Natl. Acad. Sci. USA 2011, 108, 21128–21133. [Google Scholar] [CrossRef] [PubMed]
- Jindra, M.; Uhlirova, M.; Charles, J.P.; Smykal, V.; Hill, R.J. Genetic evidence for function of the bHLH-PAS protein Gce/Met as a juvenile hormone receptor. PLoS Genet. 2015, 11, e1005394. [Google Scholar] [CrossRef]
- Li, M.; Liu, P.; Wiley, J.D.; Ojani, R.; Bevan, D.R.; Li, J.; Zhu, J. A steroid receptor coactivator acts as the DNA-binding partner of the Methoprene-tolerant protein in regulating juvenile hormone response genes. Mol. Cell. Endocrinol. 2014, 394, 47–58. [Google Scholar] [CrossRef]
- Huang, Z.Y.; Lin, S.; Ahn, K. Methoprene does not affect juvenile hormone titers in honey bee (Apis mellifera) workers. Insect Sci. 2018, 25, 235–240. [Google Scholar] [CrossRef]
- Schooneveld, H.; Sanchez, A.O.; Wilde, J.D. Juvenile hormone—Induced break and termination of diapause in the Colorado potato beetle. J. Insect Physiol. 1977, 23, 689–696. [Google Scholar] [CrossRef]
- Ojima, N.; Ishiguro, S.; An, Y.; Kadosawa, T.; Suzuki, K. Male reproductive maturity and diapause termination in the leaf beetle Gastrophysa atrocyanea. Physiol. Entomol. 2015, 40, 277–283. [Google Scholar] [CrossRef]
- Agui, N.; Mihara, M.; Kurahashi, H. Effect of juvenile hormone analogue on ovarian development of the reproductive-diapausing parasitic fly, Melinda pusilla. Jpn. J. Sanit. Zool. 1991, 42, 311–317. [Google Scholar] [CrossRef]
- Hiroyoshi, S.; Reddy, G.V.P.; Mitsuhashi, J. Effects of juvenile hormone analogue (methoprene) and 20-hydroxyecdysone on reproduction in Polygonia c-aureum (Lepidoptera: Nymphalidae) in relation to adult diapause. J. Comp. Physiol. A 2017, 203, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Hahn, D.A.; Denlinger, D.L. Energetics of insect diapause. Annu. Rev. Entomol. 2011, 56, 103–121. [Google Scholar] [CrossRef]
- Visser, B.; Ellers, J. Lack of lipogenesis in parasitoids: A review of physiological mechanisms and evolutionary implications. J. Insect Physiol. 2008, 54, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.Q.; Liu, W.; Zhu, F.; Lei, C.L.; Wang, X.P. Fatty acid synthase 2 contributes to diapause preparation in a beetle by regulating lipid accumulation and stress tolerance genes expression. Sci. Rep. 2017, 7, 40509. [Google Scholar] [CrossRef]
- Gao, Q.; Feng, S.; Liu, W.; Wang, X.P.; Wang, J.L. Research progress in juvenile hormone regulation mechanism in insect reproductive diapause. J. Environ. Entomol. 2018, 40, 988–994. [Google Scholar]
- Tan, Y.; Zhou, X.R.; Pang, B.P. Reference gene selection and evaluation for expression analysis using qRT-PCR in Galeruca daurica (Joannis). Bull. Entomol. Res. 2017, 107, 359–368. [Google Scholar] [CrossRef]
- Chen, L.; Zhou, X.R.; Gao, L.J.; Tan, Y.; Pang, B.P. Change of carbohydrate, protein and lipid contents in Galeruca daurica (Coleoptera: Chrysomelidae) adults during over summering. Acta Entomol. Sin. 2018, 61, 808–814. [Google Scholar]
- Ma, H.Y.; Zhou, X.R.; Tan, Y.; Pang, B.P. Proteomic analysis of adult Galeruca daurica (Coleoptera: Chrysomelidae) at different stages during summer diapause. Comp. Biochem. Physiol. 2019, 29, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhou, X.R.; Tan, Y.; Pang, B.P.; Xinbayin. Cloning and expression profiling of the Galeruca daurica juvenile hormone binding protein gene, GdJHBP. Chin. J. Appl. Entomol. 2020, 57, 623–631. [Google Scholar]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Xian, A.; Fan, L.; Raychowdhury, R.; Zeng, Q. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Epig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−∆∆Ct) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Colinet, H.; Vernon, P.; Hance, T. Does thermal-related plasticity in size and fat reserves influence supercooling abilities and cold-tolerance in Aphidius colemani (Hymenoptera: Aphidiinae) mummies? J. Therm. Biol. 2007, 32, 374–382. [Google Scholar] [CrossRef]
- Wang, Y.; Xue, R.D. Effect of juvenile hormone and light in the initiation and termination of diapause in Culex tritaeniorhynchus. Chin. J. Vector Biol. Control 1994, 5, 410–412. [Google Scholar]
- Liu, W.; Tan, Q.Q.; Zhu, L.; Li, Y.; Zhu, F.; Lei, C.L.; Wang, X.P. Absence of juvenile hormone signalling regulates the dynamic expression profiles of nutritional metabolism genes during diapause preparation in the cabbage beetle Colaphellus bowringi. Insect Mol. Biol. 2017, 26, 530–542. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Busche, J.M.; Zhang, X. Identification of juvenile hormone target genes in the adult female mosquitoes. Insect Biochem. Mol. Biol. 2010, 40, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Vermunt, A.W.M.; Koopmanschap, A.B.; Vlak, J.M.; de Kort, C.A. Expression of the juvenile hormone esterase gene in the Colorado potato beetle, Leptinotarsa decemlineata: Photoperiodic and juvenile hormone analog response. J. Insect Physiol. 1999, 45, 135–142. [Google Scholar] [CrossRef]
- Wroblewski, V.J.; Harshman, L.G.; Hanzlik, T.N.; Hammock, B.D. Regulation of juvenile hormone esterase gene expression in the tobacco budworm (Heliothis virescens). Arch. Biochem. Biophys. 1990, 278, 461–466. [Google Scholar] [CrossRef]
- Gu, X.; Kumar, S.; Kim, E.; Kim, Y. A whole genome screening and RNA interference identify a juvenile hormone esterase-like gene of the diamondback moth, Plutella xylostella. J. Insect Physiol. 2015, 80, 81–87. [Google Scholar] [CrossRef]
- Duan, D.; Zheng, R.; Lin, S.; Chen, Y.; Tian, H.; Zhao, J.; Tian, S.; Wei, H.; Gu, X. Modulation of juvenile hormone esterase gene expression against development of Plutella xylostella (Lepidoptera: Plutellidae). J. Econ. Entomol. 2016, 109, 865–872. [Google Scholar] [CrossRef]
- Zhu, L.; Yin, T.Y.; Sun, D.; Liu, W.; Zhu, F.; Lei, C.L.; Wang, X.P. Juvenile hormone regulates the differential expression of putative juvenile hormone esterases via methoprene-tolerant in non-diapause-destined and diapause-destined adult female beetle. Gene 2017, 627, 373–378. [Google Scholar] [CrossRef]
- Hu, X.L.; Niu, J.J.; Meng, Q.; Chai, Y.H.; Chan, K.M. Effects of two juvenile hormone analogue insecticides, fenoxycarb and methoprene, on Neocaridina davidi. Environ. Pollut. 2019, 253, 89–99. [Google Scholar] [CrossRef]
- Kim, I.H.; Pham, V.; Jablonka, W.; Goodman, W.G.; Ribeiro, J.M.C.; Andersen, J.F. A mosquito hemolymph odorant-binding protein family member specifically binds juvenile hormone. J. Biol. Chem. 2017, 292, 15329–15339. [Google Scholar] [CrossRef] [PubMed]
- Nouzova, M.; Michalkova, V.; Ramirez, C.E.; Fernandez-Lima, F.; Noriega, F.G. Inhibition of juvenile hormone synthesis in mosquitoes by the methylation inhibitor 3-deazaneplanocin A (DZNep). Insect Biochem. Mol. Biol. 2019, 113, 103183. [Google Scholar] [CrossRef] [PubMed]
- Ritdachyeng, E.; Manaboon, M.; Tobe, S.S.; Singtripop, T. Molecular characterization and gene expression of juvenile hormone binding protein in the bamboo borer, Omphisa fuscidentalis. J. Insect Physiol. 2012, 58, 1493–1501. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.N.; Liang, G.M.; Ma, L.; Jiang, T.; Xiao, H.J. Dissecting the role of juvenile hormone binding protein in response to hormone and starvation in the cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). J. Econ. Entomol. 2019, 112, 1411–1417. [Google Scholar] [CrossRef]
- Liu, W.; Li, Y.; Zhu, L.; Zhu, F.; Lei, C.L.; Wang, X.P. Juvenile hormone facilitates the antagonism between adult reproduction and diapause through the methoprene-tolerant gene in the female Colaphellus bowringi. Insect Biochem. Mol. Biol. 2016, 74, 50–60. [Google Scholar] [CrossRef]
- Belles, X.; Santos, C.G. The MEKRE93 (Methoprene tolerant-Krüppel homolog 1-E93) pathway in the regulation of insect metamorphosis, and the homology of the pupal stage. Insect Biochem. Mol. Biol. 2014, 52, 60–68. [Google Scholar] [CrossRef]
- Song, J.; Wu, Z.; Wang, Z.; Deng, Z.; Zhou, S. Krüppel-homolog 1 mediates juvenile hormone action to promote vitellogenesis and oocyte maturation in the migratory locust. Insect Biochem. Mol. Biol. 2014, 52, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Xu, Y.; Lin, X. Role of Broad-Complex (Br) and Krüppel homolog 1 (Kr-h1) in the ovary development of Nilaparvata lugens. Front. Physiol. 2017, 8, 1013. [Google Scholar] [CrossRef] [PubMed]
- Li, K.L.; Yuan, S.Y.; Nanda, S.; Wang, W.X.; Lai, F.X.; Fu, Q.; Wan, P.J. The roles of E93 and Kr-h1 in metamorphosis of Nilaparvata lugens. Front. Physiol. 2018, 9, 1677. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Yang, R.L.; Wang, W.P.; Zhou, Q.H.; Chen, E.H.; Yuan, G.R.; Wang, J.J.; Dou, W. Involvement of Met and Kr-h1 in JH-mediated reproduction of female Bactrocera dorsalis (Hendel). Front. Physiol. 2018, 9, 482. [Google Scholar] [CrossRef]
- Mao, Y.; Li, Y.; Gao, H.; Lin, X. The direct interaction between E93 and Kr-h1 mediated their antagonistic effect on ovary development of the brown planthopper. Int. J. Mol. Sci. 2019, 20, 2431. [Google Scholar] [CrossRef]
- Saha, T.T.; Roy, S.; Pei, G.; Dou, W.; Zou, Z.; Raikhel, A.S. Synergistic action of the transcription factors Krüppel homolog 1 and Hairy in juvenile hormone/Methoprene-tolerant-mediated gene-repression in the mosquito Aedes aegypti. PLoS Genet. 2019, 15, e1008443. [Google Scholar] [CrossRef]
- Hu, K.; Tian, P.; Yang, L.; Tang, Y.; Qiu, L.; He, H.; Ding, W.; Li, Y. Molecular characterization of the Krüppel-homolog 1 and its role in ovarian development in Sogatella furcifera (Hemiptera: Delphacidae). Mol. Biol. Rep. 2020, 47, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.N.; Li, X.J.; Zhao, J.J.; Zhu-Salzman, K. Cloning and characterization of Methoprene-tolerant (Met) and Krüppel homolog 1 (Kr-h1) genes in the wheat blossom midge, Sitodiplosis mosellana. Insect Sci. 2020, 27, 292–303. [Google Scholar] [CrossRef] [PubMed]
- Comas, D.; Piulachs, M.D.; Bellés, X. Induction of vitellogenin gene transcription in vitro by juvenile hormone in Blattella germanica. Mol. Cell. Endocrinol. 2001, 183, 93–100. [Google Scholar] [CrossRef]
- Parthasarathy, R.; Sun, Z.; Bai, H.; Palli, S.R. Juvenile hormone regulation of vitellogenin synthesis in the red flour beetle, Tribolium castaneum. Insect Biochem. Mol. Biol. 2010, 40, 405–414. [Google Scholar] [CrossRef]
- Smykal, V.; Bajgar, A.; Provaznik, J.; Fexova, S.; Buricova, M.; Takaki, K.; Hodkova, M.; Jindra, M.; Dolezel, D. Juvenile hormone signaling during reproduction and development of the linden bug, Pyrrhocoris apterus. Insect Biochem. Mol. Biol. 2014, 45, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Junger, M.A.; Rintelen, F.; Stocker, H.; Wasserman, J.D.; Vegh, M.; Radimerski, T.; Greenbery, M.E.; Hafen, E. The Drosophila forkhead transcription factor FOXO mediates the reduction in cell number associated with reduced insulin signaling. J. Biol. 2003, 2, 20. [Google Scholar] [CrossRef] [PubMed]
- Sim, C.; Denlinger, D.L. Insulin signaling and FOXO regulate the overwintering diapause of the mosquito Culex pipiens. Proc. Natl. Acad. Sci. USA 2008, 105, 6777–6781. [Google Scholar] [CrossRef]
- Yin, Z.J.; Dong, X.L.; Kang, K.; Chen, H.; Dai, X.Y.; Wu, G.A.; Zheng, L.; Zhai, Y.F. FoxO transcription factor regulate hormone mediated signaling on nymphal diapause. Front. Physiol. 2018, 9, 1654. [Google Scholar] [CrossRef]
- Hansen, I.A.; Sieglaff, D.H.; Shiao, S.H.; Munro, J.B.; Knox, J.; Raikhel, A.S. The forkhead box gene family of transcription factors of the yellow fever mosquito Aedes aegypti and its role in mosquito reproduction. Am. J. Trop. Med. Hyg. 2006, 75, 262. [Google Scholar]
- Peng, Z.Q.; Hao, Y.J. Role of DaFOXO1 in the regulation of superoxide dismutase gene expression and developmental duration summer diapause pupae of Delia antiqua (Diptera: Anthomyiidae). Acta Entomol. Sin. 2019, 62, 1359–1368. [Google Scholar]
- Sim, C.; Denlinger, D.L. Juvenile hormone III suppresses forkhead of transcription factor in the fat body and reduces fat accumulation in the diapausing mosquito, Culex pipiens. Insect Mol. Biol. 2013, 22, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kikuta, S.; Hagiwara-Komoda, Y.; Noda, H.; Kikawada, T. A novel member of the trehalose transporter family functions as an H(+)-dependent trehalose transporter in the reabsorption of trehalose in malpighian tubules. Front. Physiol. 2012, 3, 290. [Google Scholar] [CrossRef] [PubMed]
- Li, J.X.; Cao, Z.; Guo, S.; Tian, Z.; Liu, W.; Zhu, F.; Wang, X.P. Molecular characterization and functional analysis of two trehalose transporter genes in the cabbage beetle, Colaphellus bowringi. J. Asia Pac. Entomol. 2020, 23, 627–633. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DEG_ID | Cka vs. Ta | CKb vs. Tb | Nr_Annotation | ||||
|---|---|---|---|---|---|---|---|
| FDR | Log2FC | Regulated | FDR | Log2FC | Regulated | ||
| c85279.graph_c0 | 2.37 × 10−13 | 2.575 | up | 2.92 × 10−2 | 1.303 | up | vitellogenin-like |
| c82613.graph_c0 | 6.25 × 10−6 | 1.369 | up | 1.19 × 10−3 | 1.380 | up | fatty acid synthase-like |
| c74827.graph_c0 | 3.39 × 10−3 | 1.336 | up | 5.83 × 10−3 | 1.558 | up | uncharacterized protein LOC111513065 |
| c71191.graph_c0 | 1.61 × 10−5 | 1.272 | up | 4.84 × 10−3 | 1.228 | up | probable pseudouridine-5'-phosphatase |
| c69643.graph_c0 | 8.60 × 10−3 | −1.007 | down | 7.39 × 10−4 | −1.295 | down | retinoid-inducible serine carboxypeptidase-like |
| c37561.graph_c0 | 2.11 × 10−6 | −1.034 | down | 1.46 × 10−9 | −1.617 | down | PREDICTED: crustapain-like |
| c64926.graph_c0 | 5.25 × 10−3 | −1.065 | down | 7.01 × 10−11 | −1.777 | down | -- |
| c84589.graph_c0 | 9.74 × 10−5 | −1.160 | down | 1.64 × 10−5 | −1.526 | down | protein msta-like |
| c72112.graph_c0 | 1.80 × 10−5 | −1.360 | down | 6.99 × 10−30 | −2.327 | down | facilitated trehalose transporter Tret1-like |
| c73343.graph_c0 | 2.44 × 10−3 | −1.365 | down | 4.20 × 10−2 | −1.330 | down | -- |
| c82886.graph_c0 | 3.61 × 10−6 | −1.401 | down | 4.31 × 10−2 | −1.235 | down | nose resistant to fluoxetine protein 6-like isoform X1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, H.-Y.; Li, Y.-Y.; Li, L.; Tan, Y.; Pang, B.-P. Regulation of Juvenile Hormone on Summer Diapause of Geleruca daurica and Its Pathway Analysis. Insects 2021, 12, 237. https://doi.org/10.3390/insects12030237
Ma H-Y, Li Y-Y, Li L, Tan Y, Pang B-P. Regulation of Juvenile Hormone on Summer Diapause of Geleruca daurica and Its Pathway Analysis. Insects. 2021; 12(3):237. https://doi.org/10.3390/insects12030237
Chicago/Turabian StyleMa, Hong-Yue, Yan-Yan Li, Ling Li, Yao Tan, and Bao-Ping Pang. 2021. "Regulation of Juvenile Hormone on Summer Diapause of Geleruca daurica and Its Pathway Analysis" Insects 12, no. 3: 237. https://doi.org/10.3390/insects12030237
APA StyleMa, H.-Y., Li, Y.-Y., Li, L., Tan, Y., & Pang, B.-P. (2021). Regulation of Juvenile Hormone on Summer Diapause of Geleruca daurica and Its Pathway Analysis. Insects, 12(3), 237. https://doi.org/10.3390/insects12030237