
The Control of the American Leafhopper Erasmoneura vulnerata (Fitch) in European Vineyards: Impact of Synthetic and Natural Insecticides


 and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
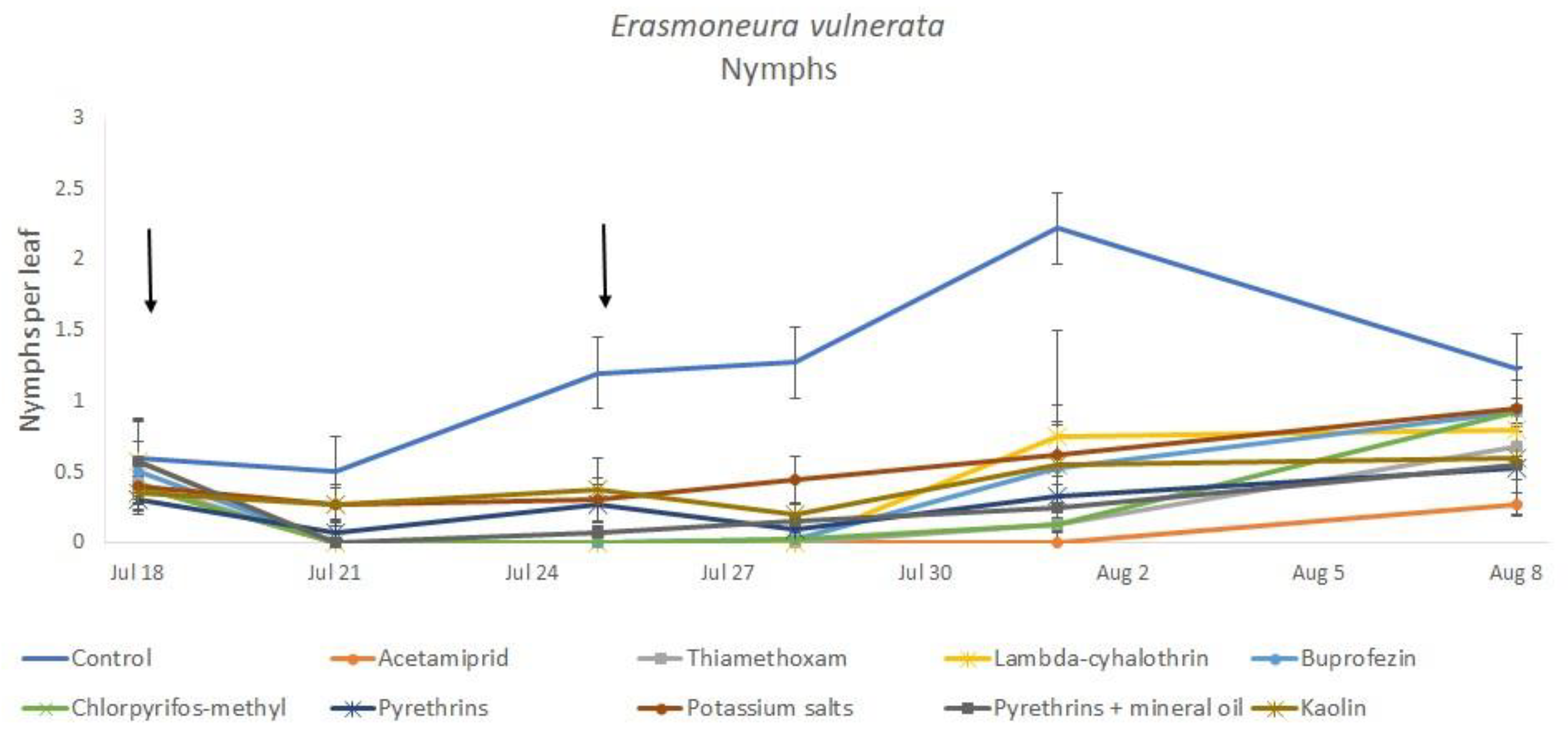
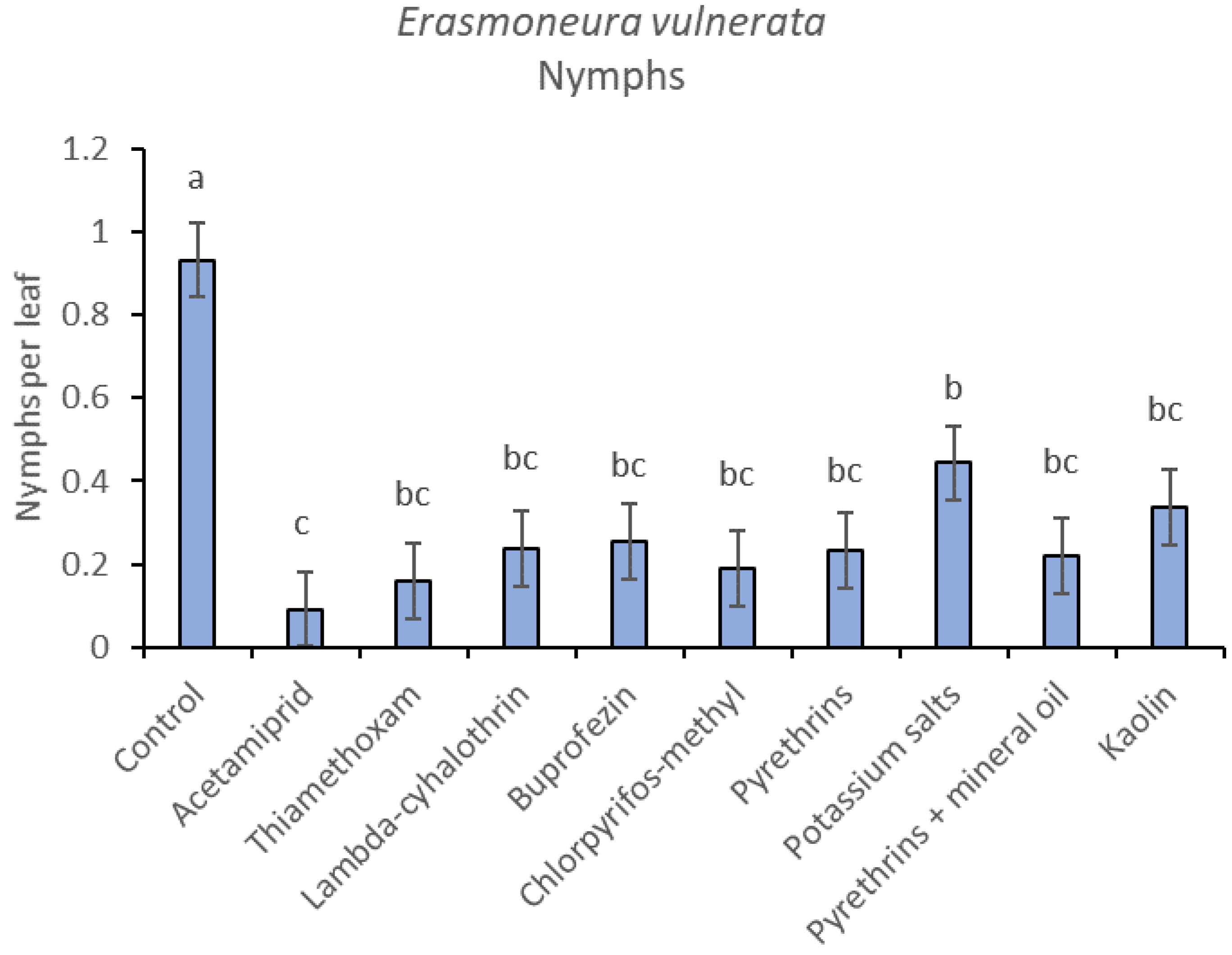
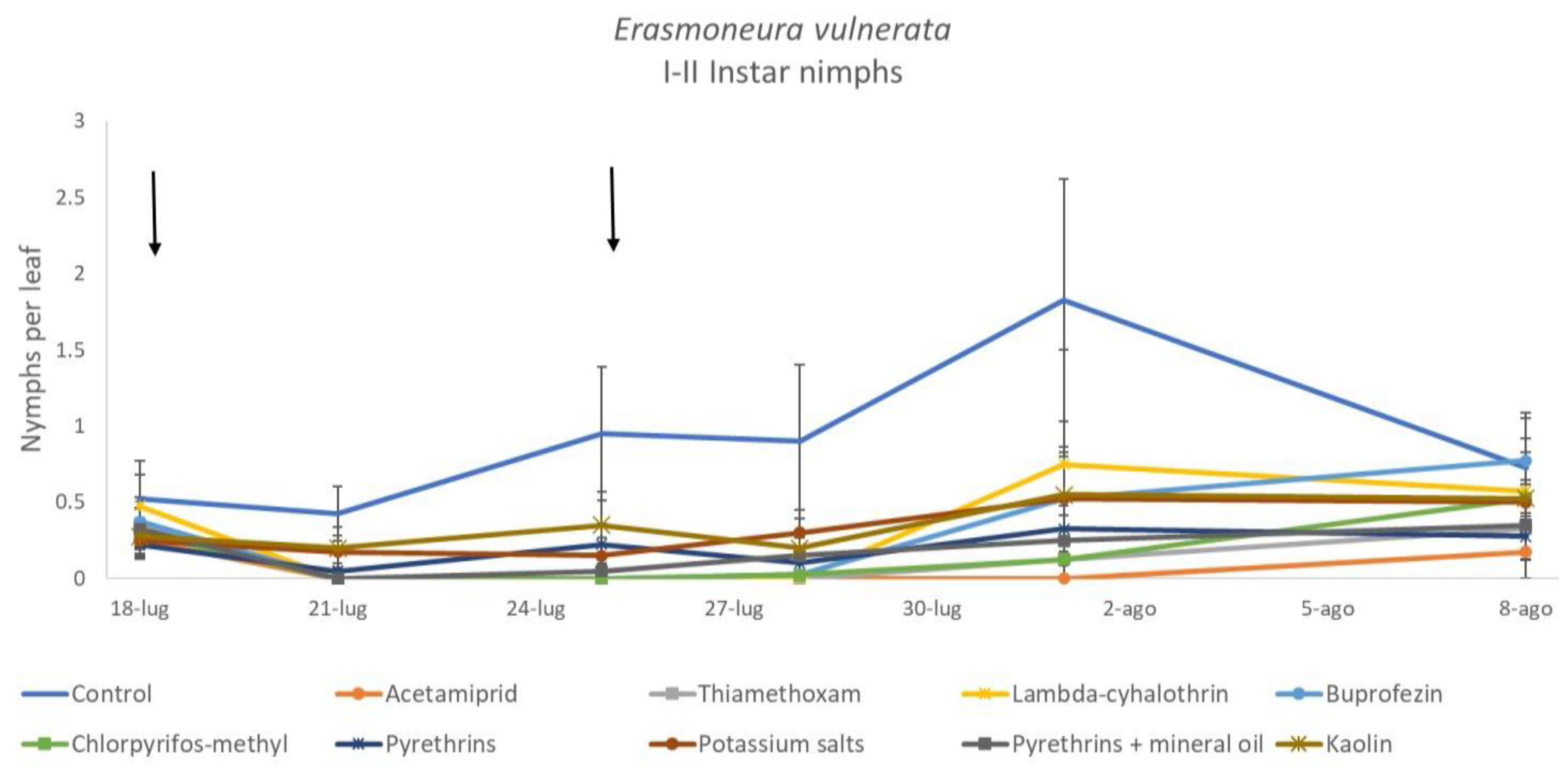
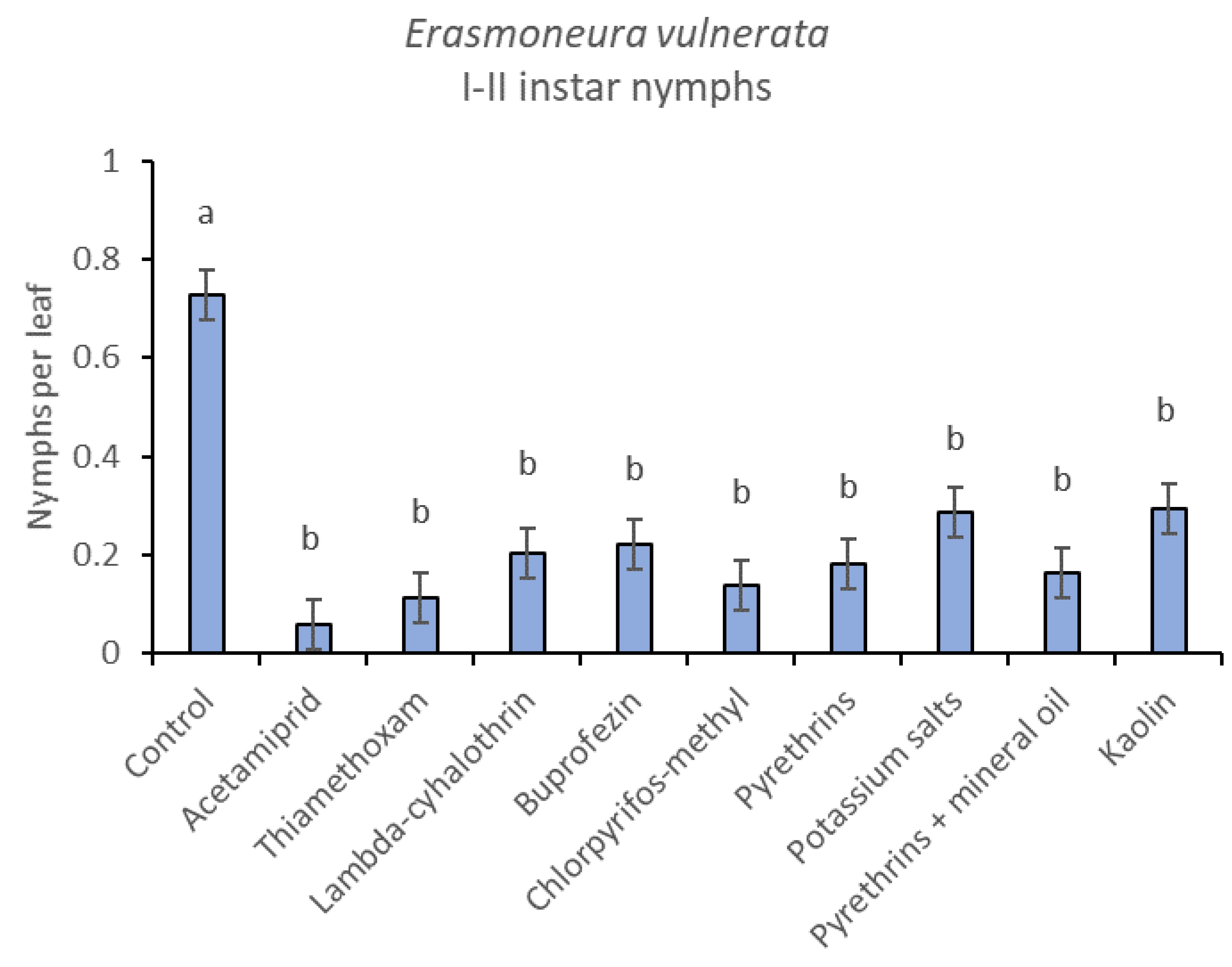
3. Results
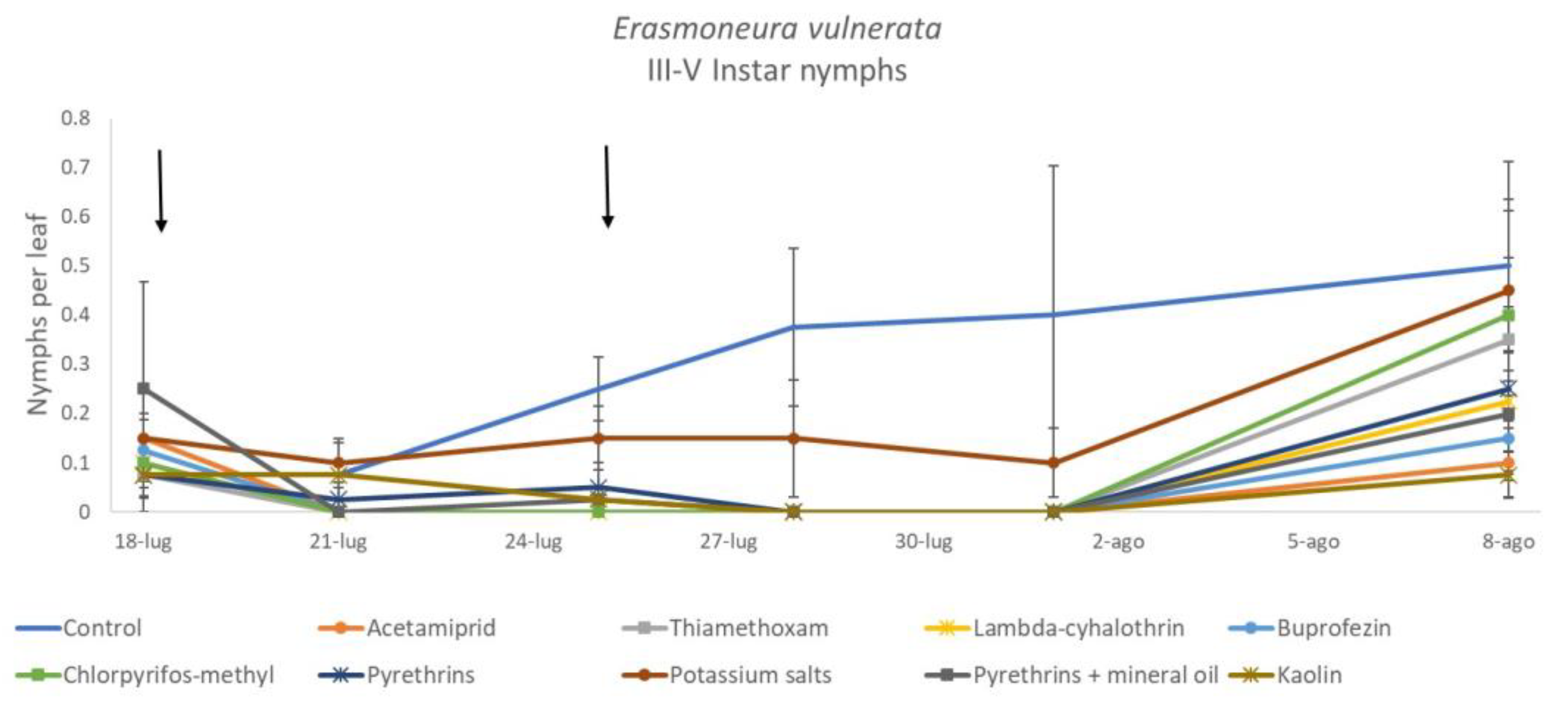
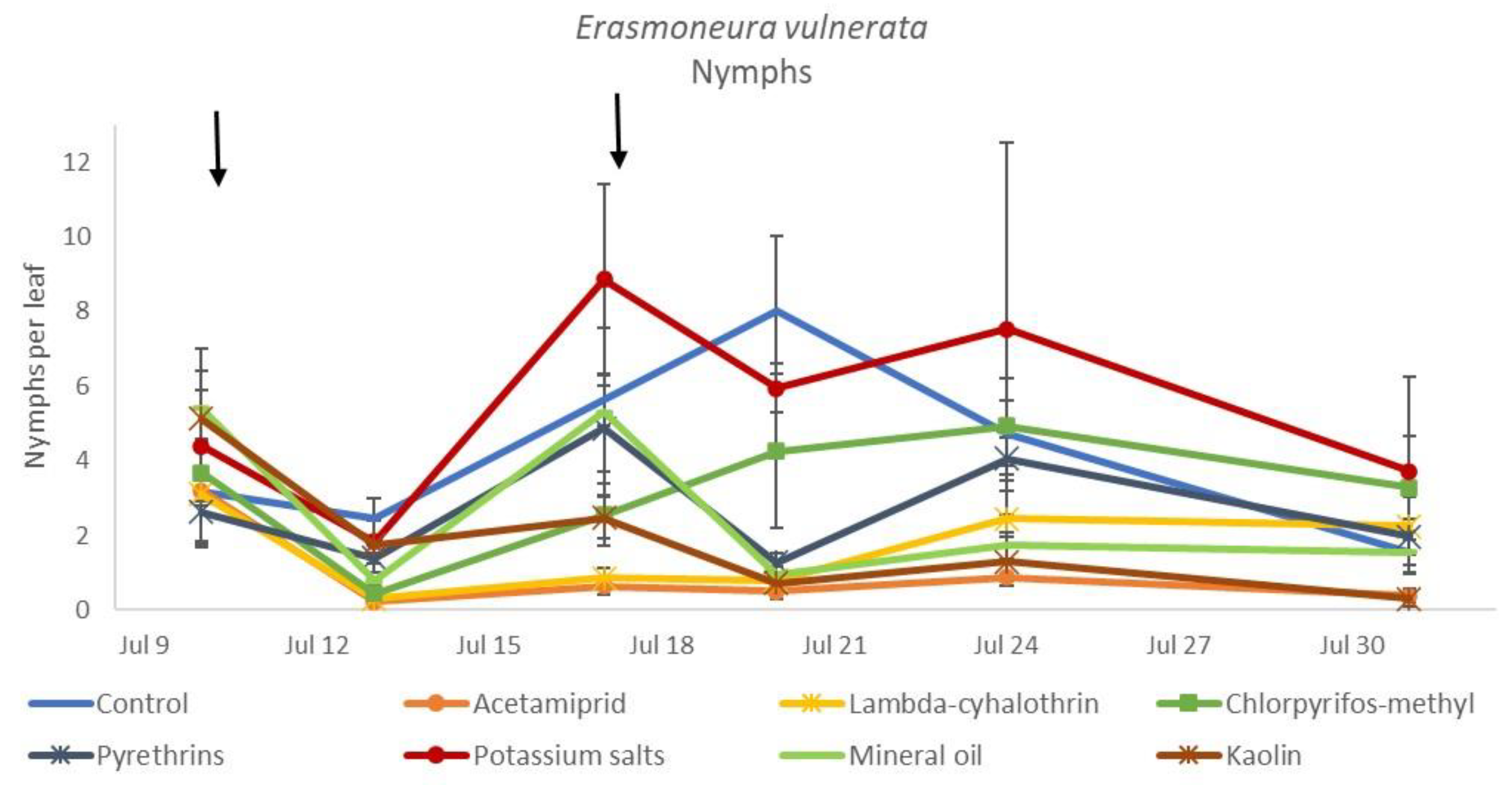
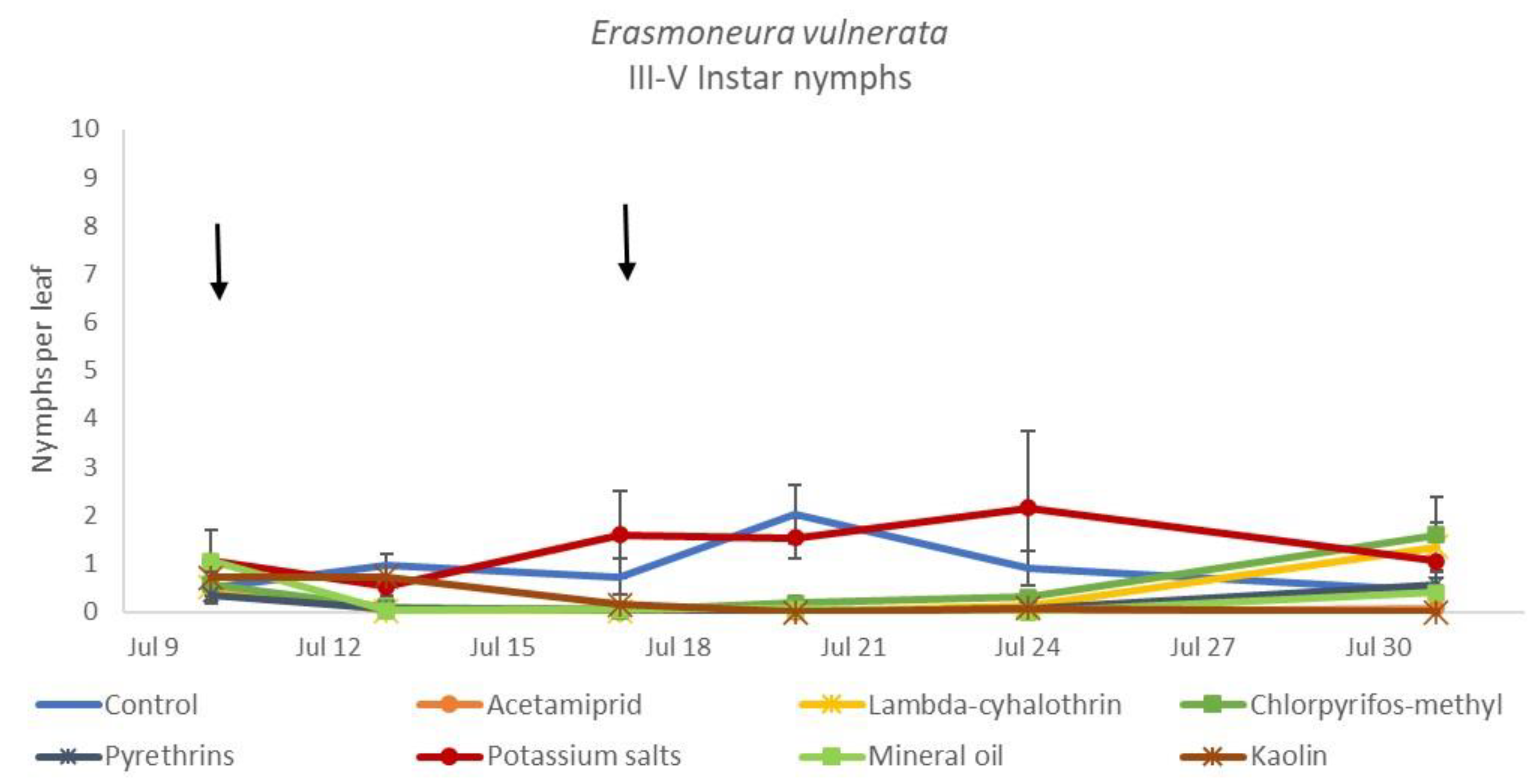
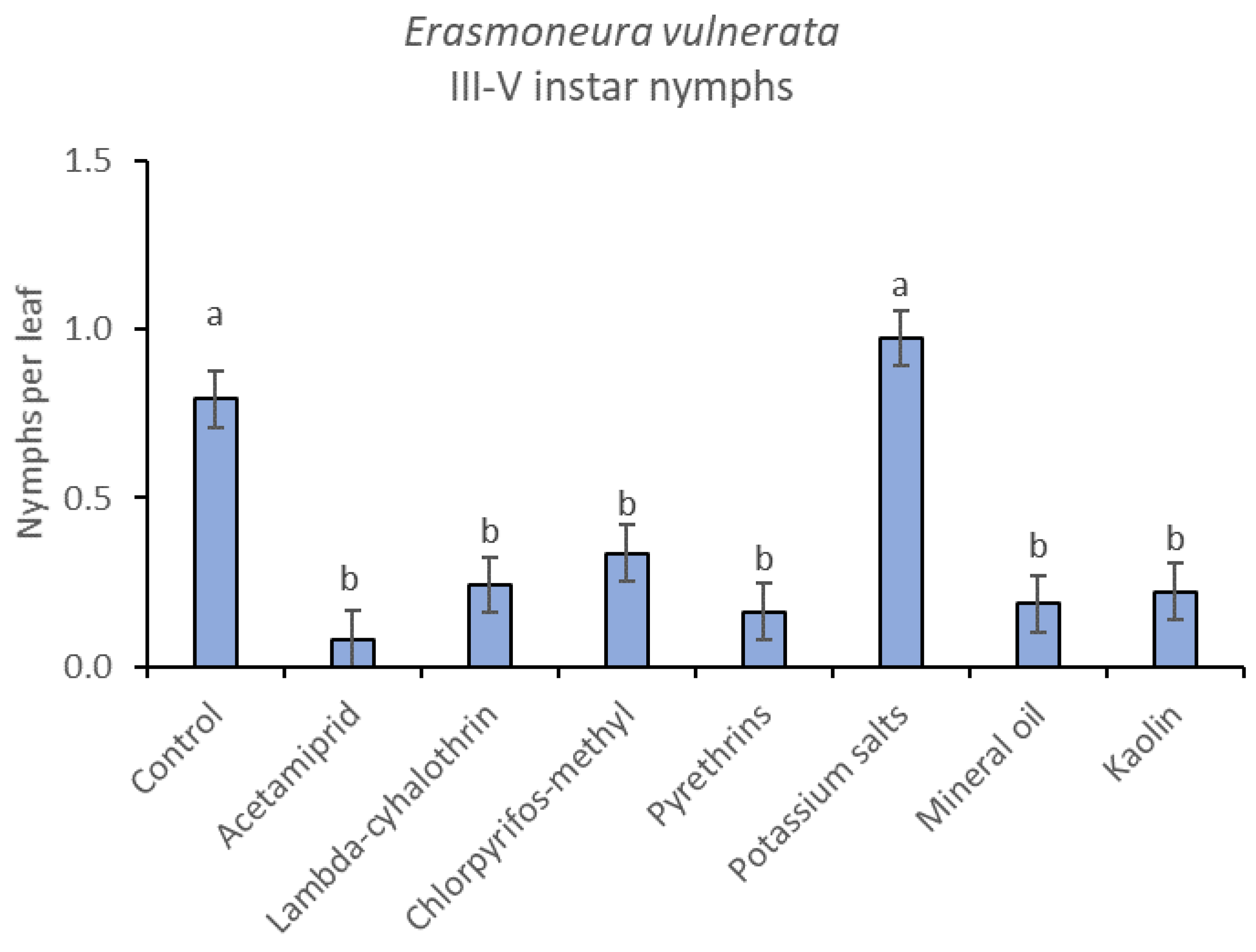
3.1. 2017
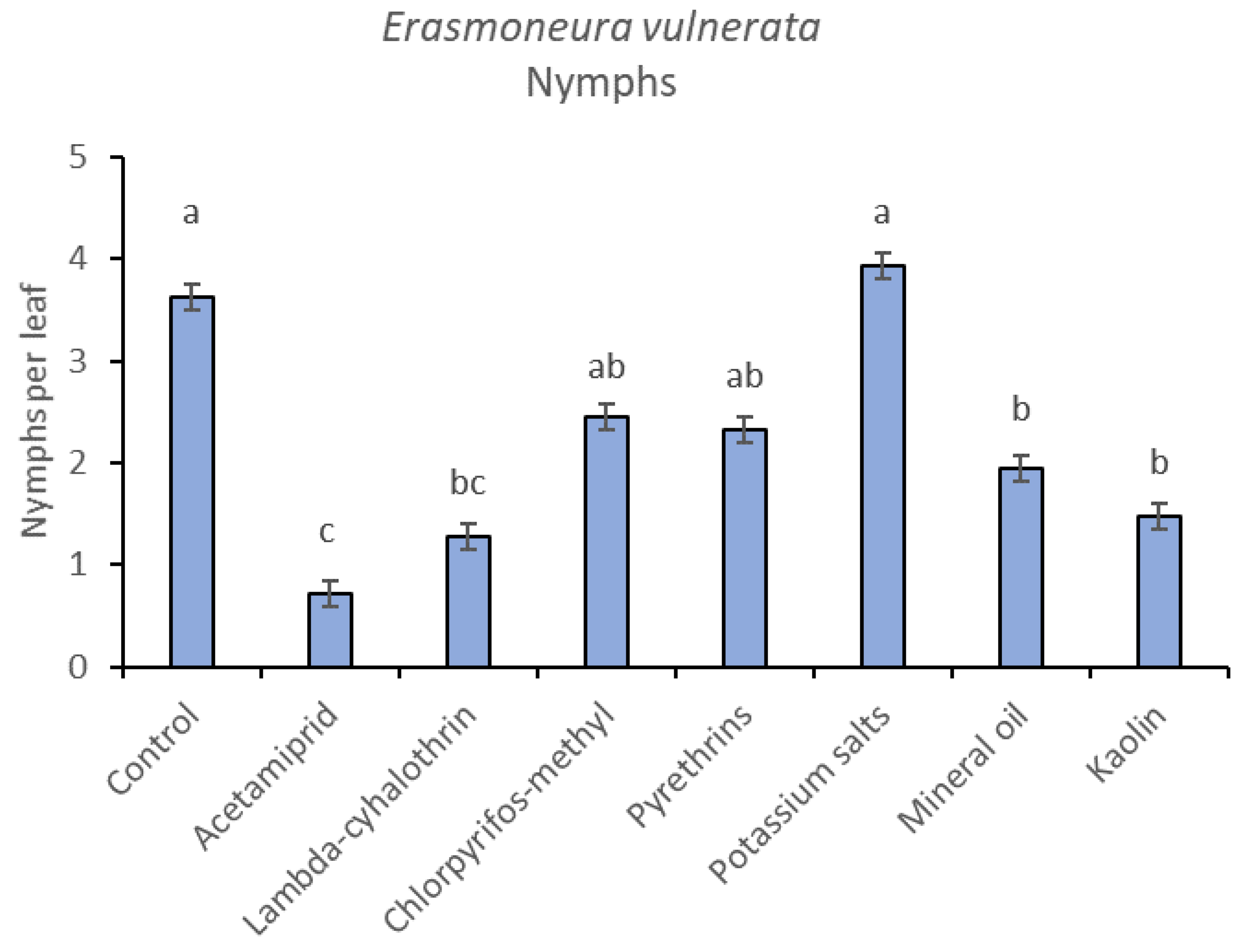
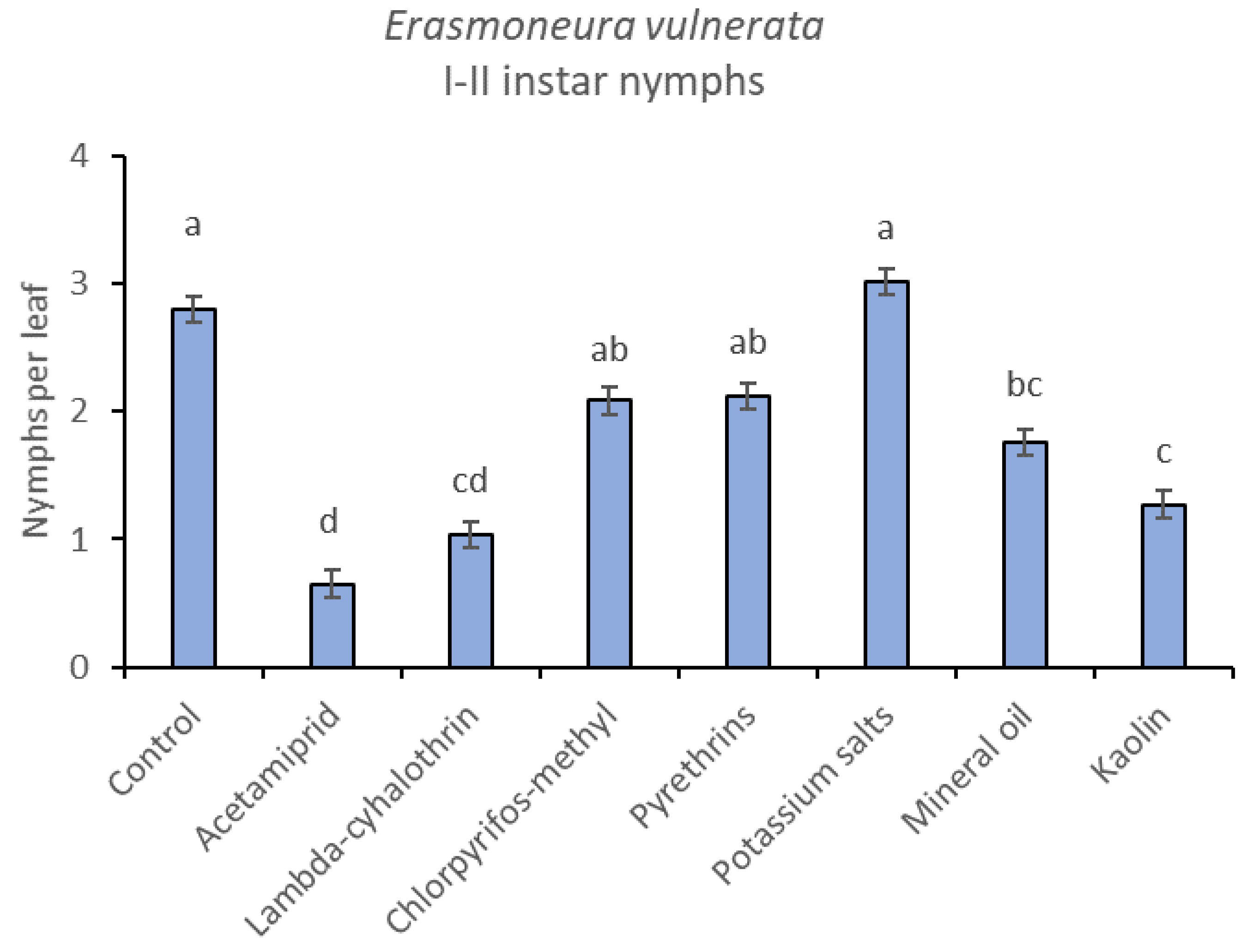
3.2. 2018
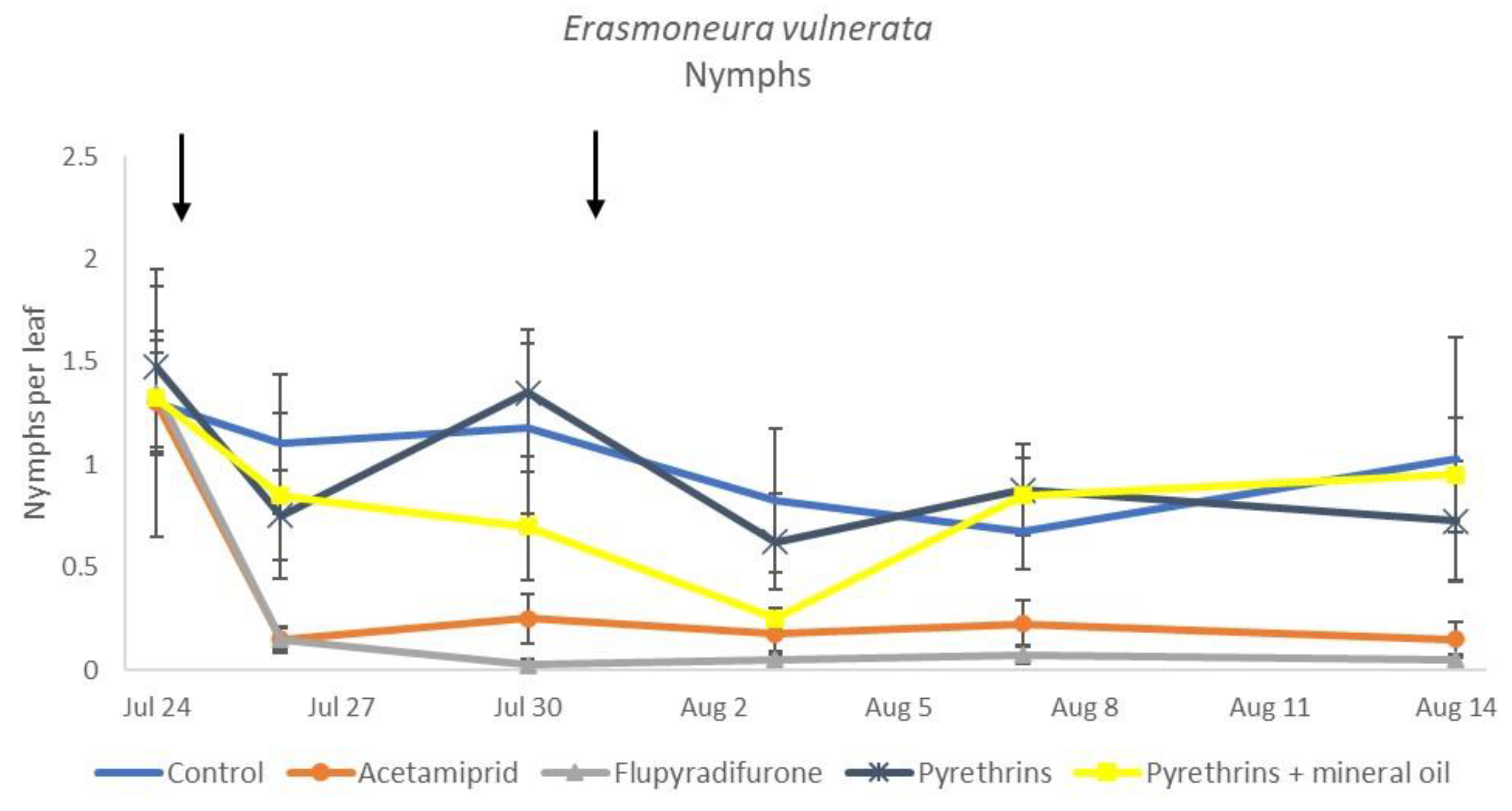
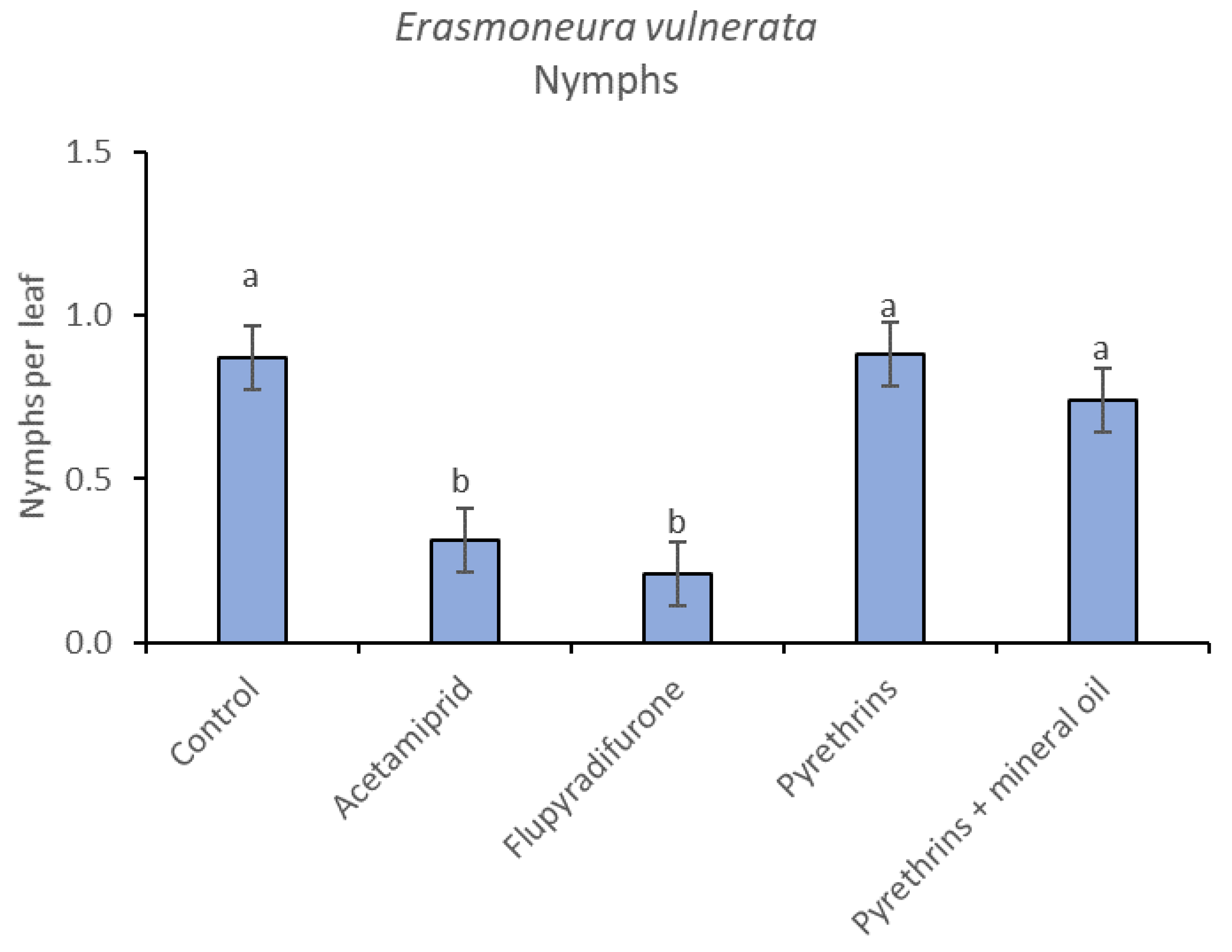
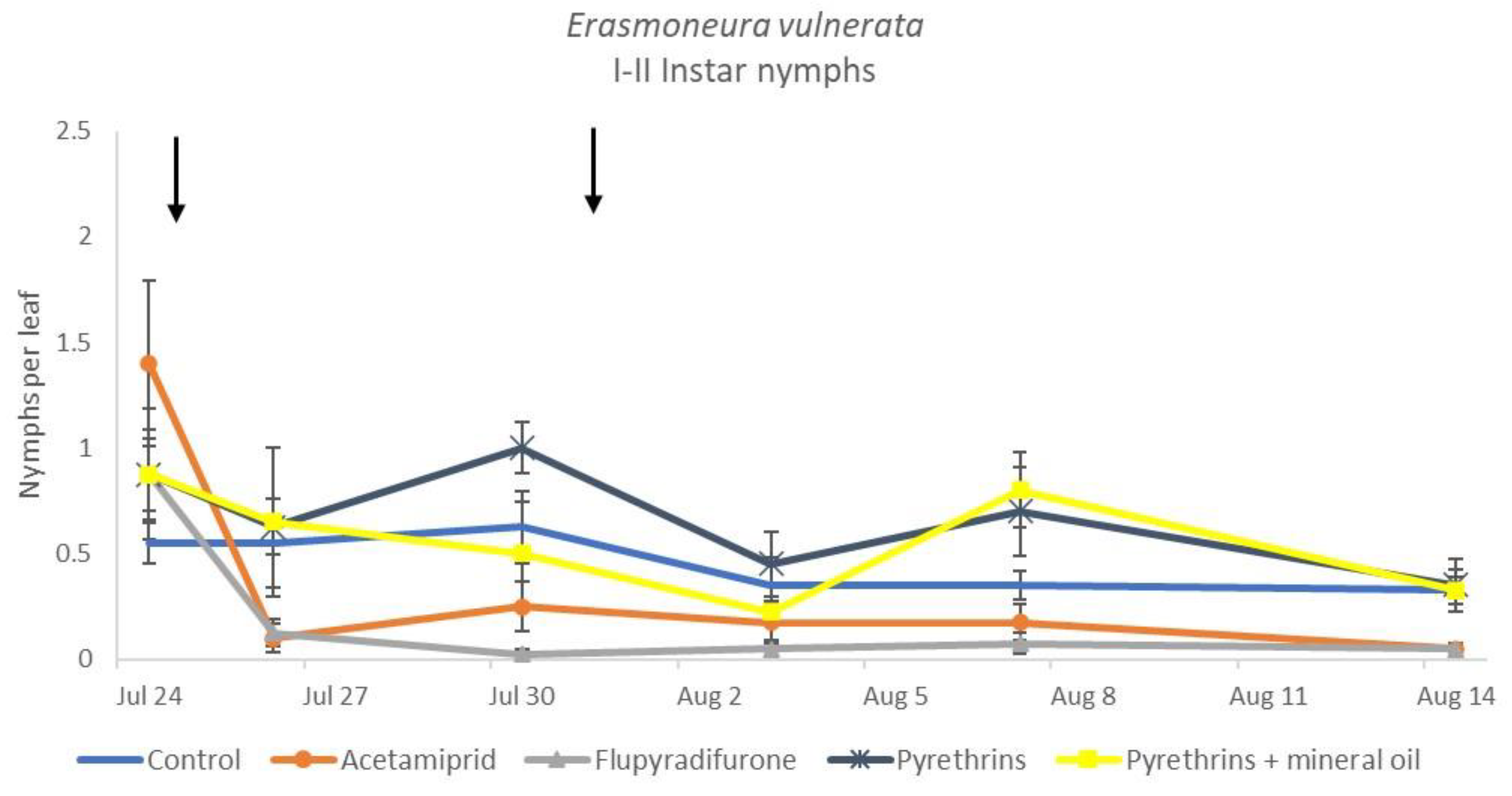
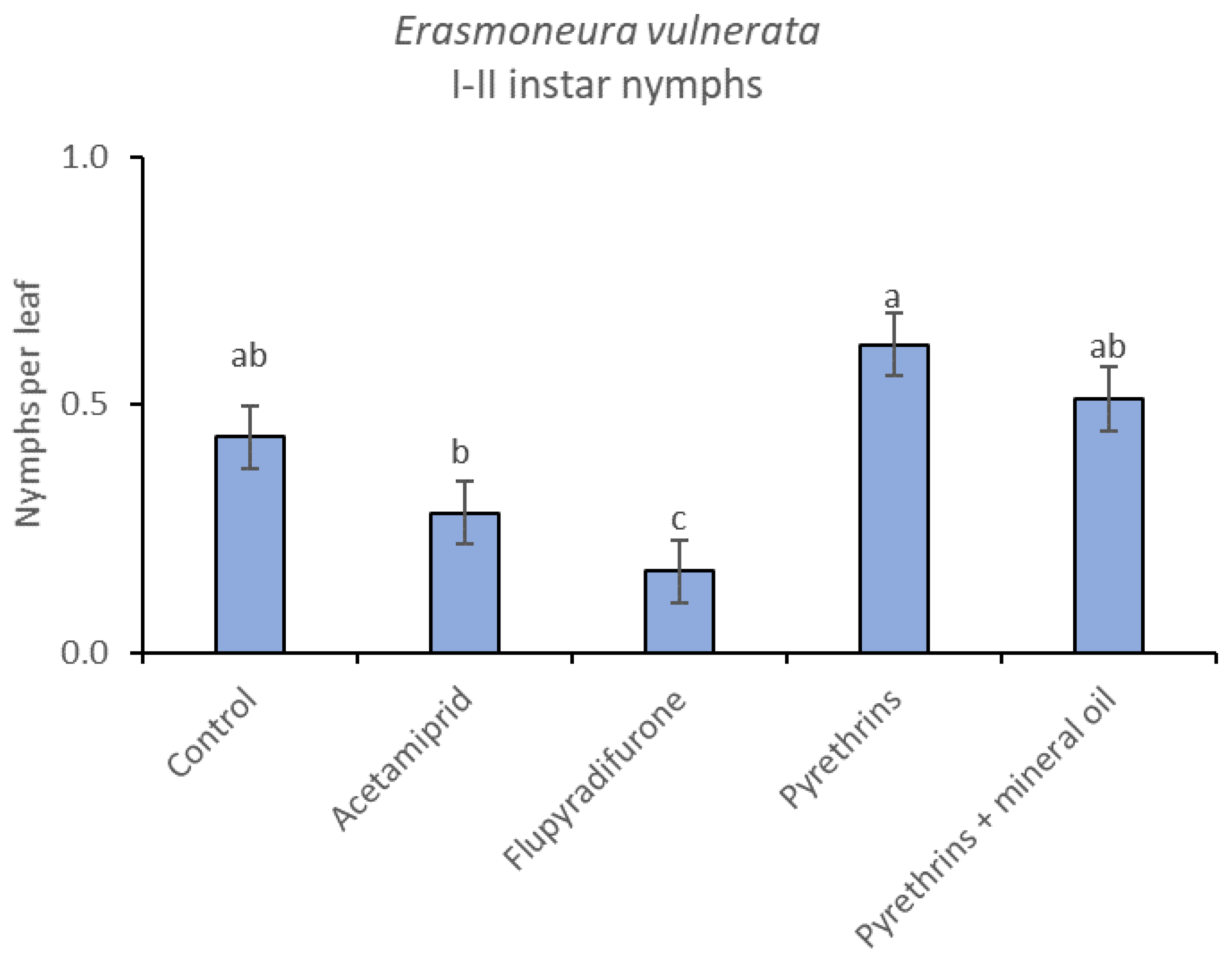
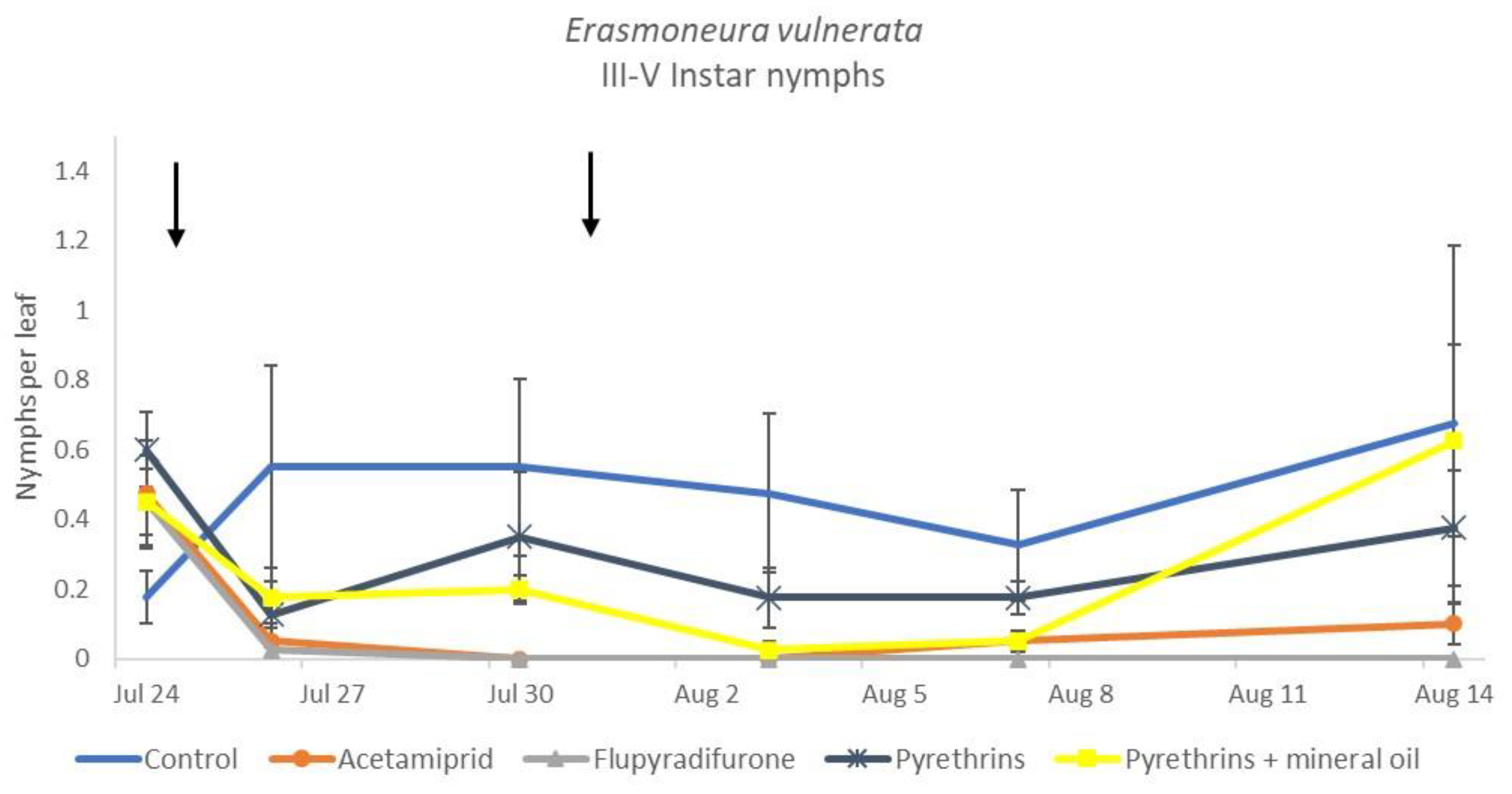
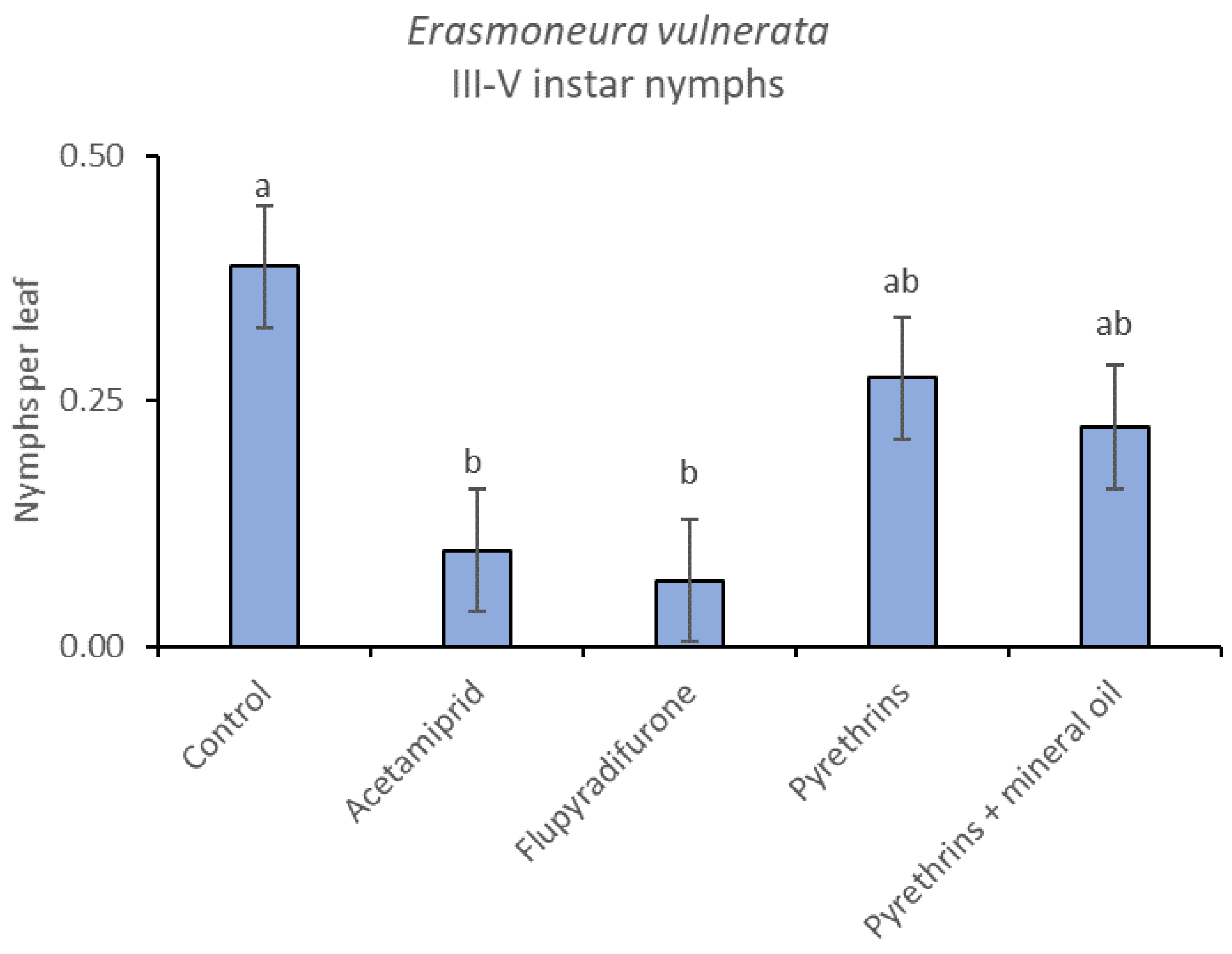
3.3. 2019
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vidano, C. Le cicaline italiane della vite. Hemiptera Typhlocibinae. Boll. Zool. Agr. Bachic. 1958, 1, 61–115. [Google Scholar]
- Vidano, C. Alterazioni provocate da insetti in Vitis osservate, sperimentate e comparate. Ann. Fac. Sci. Agric. Univ. Torino 1963, 1, 513–644. [Google Scholar]
- Schvester, D.; Moutous, G.; Bonfils, J.; Carle, P. Ètude biologique des cicadelles de la vigne dans le sud-ouest de la France. Ann. Epiphyt. 1962, 13, 205–237. [Google Scholar]
- Moutous, G.; Fos, A. Influence des niveaux de populations de cicadelles de la vigne (Empoasca flavescens Fab.) sur le symptôme de la grillure des feuilles. Ann. Zool. Ecol. Anim. 1973, 5, 173–185. [Google Scholar]
- Cerutti, F.; Baumgärtner, J.; Delucchi, V. Ricerche sull’ecosistema ‘vigneto’ in Ticino: I. Campionamento delle popolazioni di Empoasca vitis Goethe (Hom., Cicadellidae, Typhlocybinae). Mitt. Schweiz. Entomol. Ges. 1988, 61, 29–41. [Google Scholar]
- Pavan, F.; Pavanetto, E.; Duso, C.; Girolami, V. Population dynamics of Empoasca vitis (Goethe) and Zygina rhamni (Ferr.) on vines in northern Italy. In Proceedings of the 6th Auchenorrhyncha Meeting, Turin, Italy, 7–11 September 1987; Vidano, C., Arzone, A., Eds.; Consiglio Nazionale delle Ricerche: Turin, Italy, 1988; pp. 517–524. [Google Scholar]
- Decante, D.; van Helden, M. Population ecology of Empoasca vitis (Goethe) and Scaphoideus titanus (Ball) in Bordeaux vineyards: Influence of migration and landscape. Crop Prot. 2006, 25, 696–704. [Google Scholar] [CrossRef]
- Pavan, F.; Picotti, P.; Girolami, V. Strategie per il controllo di Empoasca vitis Goethe su vite. L’Informatore Agrar. 1992, 48, 65–72. [Google Scholar]
- Baillod, M.; Jermini, M.; Antonin, P.; Linder, C.; Mittaz, C.; Carrera, E.; Udry, V.; Schmid, A. Stratégies de lutte contre la cicadelle verte de la vigne, Empoasca vitis (Goethe). Efficacité des insecticides et problématique liée à la nuisibilité. Rev. Suisse Vitic. Arboric. Hortic. 1993, 25, 133–141. [Google Scholar]
- Bonafos, R.; Serrano, E.; Auger, P.; Kreiter, S. Resistance to deltamethrin, lambda-cyhalothrin and chlorpyriphos-ethyl in some populations of Typhlodromus pyri Scheuten and Amblyseius andersoni (Chant) (Acari:Phytoseiidae) from vineyards in the south-west of France. Crop Prot. 2007, 26, 169–172. [Google Scholar] [CrossRef]
- Girolami, V.; Mori, N.; Marchesini, E.; Duso, C. Organophosphate resistance in grape leafhoppers and IPM strategies. Redia 2001, 84, 1–17. [Google Scholar]
- Vidano, C.; Arzone, A.; Arnò, C. Researches on natural enemies of viticolous Auchenorrhyncha. In Integrated pest control in viticolture. In Proceedings of the Meet of EC Experts’ Group, Portoferraio, Italy, 26–28 September 1985; pp. 97–101. [Google Scholar]
- Cerutti, F.; Delucchi, V.; Baumgärtner, J.; Rubli, D. Ricerche sull’ecosistema ‘vigneto’ in Ticino: II. La colonizzazione dei vigneti da parte della cicalina Empoasca vitis Goethe (Hom., Cicadellide, Typhlocybinae) e del suo parassitoide Anagrus atomus Haliday (Hym. Mymaridae), e importanza della flora circostante. Mitt. Schweiz. Entomol. Ges. 1989, 62, 253–267. [Google Scholar]
- Van Helden, M.; Decante, D. The possibilities for conservation biological control as a management strategy against Empoasca vitis. Integrated control in viticulture. IOBC/WPRS Bull. 2001, 24, 291–297. [Google Scholar]
- Ponti, R.; Ricci, C.; Torricelli, R. The ecological role of hedges on population dynamics of Anagrus spp. (Hymenoptera: Mymaridae) in vineyards of Central Italy. IOBC/WPRS Bull. 2003, 26, 117–122. [Google Scholar]
- Zanolli, P.; Martini, M.; Mazzon, L.; Pavan, F. Morphological and Molecular Identification of Anagrus ‘atomus’ Group (Hymenoptera: Mymaridae) Individuals from Different Geographic Areas and Plant Hosts in Europe. J. Insect Sci. 2016, 38, 1–14. [Google Scholar]
- Schvester, D.; Carle, P.; Moutous, G. Transmission de la flavescence dorée de la vigne par Scaphoideus littoralis Ball, (Homopt. Jassidae). Ann. Epiphyt. 1961, 14, 175–198. [Google Scholar]
- Angelini, E.; Negrisolo, E.; Clair, D.; Borgo, M.; Boudon-Padieu, E. Phylogenetic relationships among Flavescence dorée strains and related phytoplasmas determined by heteroduplex mobility assay and sequence of ribosomal and nonribosomal DNA. Plant Pathol. 2003, 52, 663–672. [Google Scholar] [CrossRef]
- Malembic-Maher, S.; Desqué, D.; Khalil, D.; Salar, P.; Bergey, B.; Danet, J.-L.; Duret, S.; Dubrana-Ourabah, M.-P.; Beven, L.; Ember, I.; et al. When a Palearctic bacterium meets a Nearctic insect vector: Genetic and ecological insights into the emergence of the grapevine Flavescence dorée epidemics in Europe. PLoS Pathog. 2020, 16, e1007967. [Google Scholar] [CrossRef]
- Chuche, J.; Thiery, D. Biology and ecology of the flavescence dorée vector Scaphoideus titanus: A review. Agron. Sust. Develop. 2014, 34, 381–403. [Google Scholar] [CrossRef]
- Alma, A.; Lessio, F.; Gonella, E.; Picciau, L.; Mandrioli, M.; Tota, F. New insights in phytoplasma-vector interaction: Acquisition and inoculation of flavescence dorée phytoplasma by Scaphoideus titanus adults in a short window of time. Ann. App. Biol. 2018, 173, 55–62. [Google Scholar] [CrossRef]
- Mori, N.; Pavan, F.; Lessio, F.; Alma, A. Vettori dei giallumi della vite, fra vecchie e nuove conoscenze. L’Informatore Agrar. 2020, 10, 56–59. [Google Scholar]
- Gusberti, M.; Jermini, M.; Wyss, E.; Linder, C. Efficacité d’insecticides contre Scaphoideus titanus en vignobles biologiques et effets secondaires. Rev. Suisse Vitic. Arboric. Hortic. 2008, 40, 173–177. [Google Scholar]
- Tacoli, F.; Mori, N.; Pozzebon, A.; Cargnus, E.; Da Vià, S.; Zandigiacomo, P.; Duso, C.; Pavan, F. Control of Scaphoideus titanus with natural products in organic vineyards. Insects 2017, 8, 129. [Google Scholar] [CrossRef] [PubMed]
- Delaiti, M.; Angeli, G.; Sandri, O.; Tomasi, C.; Ioriatti, C. Nuovi insetticidi per il contenimento della cicalina verde della vite. L’Informatore Agrar. 2005, 25, 73–76. [Google Scholar]
- Posenato, G.; Marchesini, E.; Mori, N. Efficacia di thiamethoxam su Empoasca vitis a confronto con lambda-cyhalothrin, abamectina, indoxacarb e chlorpyrifos. Atti Delle Giornate Fitopatol. 2006, 1, 111–116. [Google Scholar]
- Pozzebon, A.; Pederiva, M.; Moret, R.; Duso, C. The impact of a number of insecticides on Empoasca vitis populations in north-eastern Italy. IOBC/WPRS Bull. 2011, 67, 285–288. [Google Scholar]
- Pavan, F.; Mori, N.; Bressan, S.; Mutton, P. Control strategies for grapevine phytoplasma diseases: Factors influencing the profitability of replacing symptomatic plants. Phytopathol. Mediterr. 2012, 51, 11–22. [Google Scholar]
- Duso, C.; Bressan, A.; Mazzon, L.; Girolami, V. First record of the grape leafhopper Erythroneura vulnerata Fitch (Hom., Cicadellidae) in Europe. J. Appl. Entomol. 2005, 129, 170–172. [Google Scholar] [CrossRef]
- Duso, C.; Borgo, M.; Pozzebon, A.; Mazzon, L.; Mori, N.; Pavan, F.; Fornasiero, D.; Marchesini, E.; Martinez-Sañudo, I.; Zanettin, G. Vite: Erasmoneura vulnerata, una minaccia da valutare. L’Informatore Agrar. 2017, 44, 31–35. [Google Scholar]
- Olivier, C.; Vincent, C.; Saguez, J.; Galka, B.; Weintraub, P.G.; Maixner, M. Leafhoppers and planthoppers: Their bionomics, pathogen transmission and management in vineyards. In Arthropod Management in Vineyards; Springer: Dordrecht, The Netherlands, 2012; pp. 253–270. [Google Scholar]
- Zimmerman, R.; Kondratieff, B.; Nelson, E.; Sclar, C. The life history of two species of grape leafhoppers on wine grapes in western Colorado. J. Kans. Entomol. Soc. 1996, 69, 337–345. [Google Scholar]
- Duso, C.; Moret, R.; Manera, A.; Berto, D.; Fornasiero, D.; Marchegiani, G.; Pozzebon, A. Investigations on the grape leafhopper Erasmoneura vulnerata in North-eastern Italy. Insects 2019, 10, 44. [Google Scholar] [CrossRef]
- Seljak, G. First record of the Nearctic leafhopper Erasmoneura vulnerata (Fitch, 1851) (Hemiptera, Cicadomorpha: Cicadellidae) in Slovenia. Acta Entomol. Slov. 2011, 19, 37–42. [Google Scholar]
- Rizzoli, A.; Battelli, R.; Conedera, M.; Jermini, M. First record of Erasmoneura vulnerata Fitch, 1851 (Hemiptera, Cicadellidae, Typhlocybinae) in Switzerland. Alp. Entomol. 2020, 4, 151. [Google Scholar] [CrossRef]
- Duso, C.; Zanettin, G.; Gherardo, P.; Pasqualotto, G.; Raniero, D.; Rossetto, F.; Tirello, P.; Pozzebon, A. Colonization patterns, phenology and seasonal abundance of the Nearctic leafhopper Erasmoneura vulnerata (Fitch), a new pest in European vineyards. Insects 2020, 11, 731. [Google Scholar] [CrossRef] [PubMed]
- O’Hearn, J.S.; Walsh, D.B. Evaluating the Toxicity of Candidate Organic and Conventional Insecticides on Western Grape Leafhopper and Virginia Creeper Leafhopper (Hemiptera: Cicadellidae) under Vineyard and Laboratory Conditions. J. Entomol. Sci. 2020, 52, 95–103. [Google Scholar] [CrossRef]
- Lavezzaro, S.; Morando, A.; Gallesio, G. Un quadriennio di prove di lotta contro la cicalina verde della vite in Piemonte. Atti Delle Giornate Fitopatol. 2006, 1, 117–122. [Google Scholar]
- Medina-Pastor, P.; Triacchini, G. The 2018 European Union report on pesticide residues in food. EFSA J. 2020, 18, 6057. [Google Scholar] [CrossRef]
- Jactel, H.; Verheggen, F.; Thiéry, D.; Escobar-Gutiérrez, A.J.; Gachet, E.; Desneux, N. Alternatives to neonicotinoids. Environ. Int. 2019, 129, 423–429. [Google Scholar] [CrossRef]
- Mori, N.; Posenato, G.; Sancassani, G.; Tosi, L.; Girolami, V. Insetticidi per il controllo delle cicaline nei vigneti. L’Informatore Agrar. 1999, 15, 93–97. [Google Scholar]
- Puterka, G.J.; Reinke, M.; Luvisi, D.; Ciomperik, M.A.; Bartels, D.; Wendel, L.; Glenn, D.M. Particle film, Surround WP, effects on glassy-winged sharpshooter behavior and its utility as a barrier to sharpshooter infestations in grape. Plant Health Prog. 2003. [Google Scholar] [CrossRef]
- Tacoli, F.; Pavan, F.; Cargnus, E.; Tilatti, E.; Pozzebon, A.; Zandigiacomo, P. Efficacy and mode of action of kaolin in the control of Empoasca vitis and Zygina rhamni (Hemiptera: Cicadellidae) in vineyards. J. Econ. Entomol. 2017, 110, 1164–1178. [Google Scholar] [CrossRef]
- Rossetto, F. Effetti di insetticidi su Erasmoneura vulnerata e valutazioni sui danni causati dall’insetto. Second-cycle degree. In Agricultural Science and Technology; Padova University: Padova, Italy, 2019. [Google Scholar]
- Duso, C.; Van Leeuwen, T.; Pozzebon, A. Improving the compatibility of pesticides and predatory mites: Recent findings on physiological and ecological selectivity. Curr. Opin. Insect Sci. 2020, 39, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Pozzebon, A.; Tirello, P.; Moret, R.; Pederiva, M.; Duso, C. A fundamental step in IPM on grapevine: Evaluating the side effects of pesticides on predatory mites. Insects 2015, 6, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Duso, C.; Ahmad, S.; Tirello, P.; Pozzebon, A.; Klaric, V.; Baldessari, M.; Malagnini, V.; Angeli, G. The impact of insecticides applied in apple orchards on the predatory mite Kampimodromus aberrans (Acari Phytoseiidae). Exp. Appl. Acarol. 2014, 62, 391–414. [Google Scholar] [CrossRef] [PubMed]
- Tacoli, F.; Cargnus, E.; Duso, C.; Pozzebon, A.; Tirello, P.; Pavan, F. Side effects of kaolin and bunch-zone leaf removal on predatory mite populations (Acari: Phytoseiidae) occurring in vineyards. J. Econ. Entomol. 2019, 112, 1292–1298. [Google Scholar] [CrossRef] [PubMed]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Active | Trade | Concentration | Dose | Group | Number of |
|---|---|---|---|---|---|---|
| Ingredients | Mark | Applications | ||||
| 2017 | Untreated control | - | - | - | - | - |
| Acetamiprid | Epik SL | 50 g/L | 150 mL/hL | Neonicotinoids | 1 | |
| Thiamethoxam | Actara 25 WG | 25% | 20 g/hL | Neonicotinoids | 1 | |
| Lambda-cyhalothrin | Karate Zeon | 9.40% | 25 mL/hL | Pyrethroids | 1 | |
| Buprofezin | Applaud Plus | 25.00% | 200 g/hL | Thiadiazines | 1 | |
| Chlorpyrifos-methyl | Reldan LO | 21.40% | 150 mL/hL | Organophosphates | 1 | |
| Pyrethrins | Biopiren Plus | 18.6 g/L | 160 mL/hL | Pyrethrins | 2 | |
| Pyrethrins + | Biopiren Plus + | 18.6 g/L | 160 mL/hL | Pyrethrins + | 2 | |
| mineral oil | Chemol Plus | 80% | 500 mL/hL | Mineral oils | ||
| Potassium salts | Ciopper | 455 g/L | 150 mL/hL | Salts | 2 | |
| Mineral oil | Chemol Plus | 80% | 500 mL/hL | Mineral oils | 2 | |
| Kaolin | Surround | 95% | 4 Kg/hL | Kaolin | 2 | |
| 2018 | Untreated control | - | - | - | - | - |
| Acetamiprid | Epik SL | 50 g/L | 150 mL/hL | Neonicotinoids | 1 | |
| Lambda-cyhalothrin | Karate Zeon | 9.40% | 25 mL/hL | Pyrethroids | 1 | |
| Chlorpyrifos-methyl | Reldan LO | 21.40% | 150 mL/hL | Organophosphates | 1 | |
| Pyrethrins | Biopiren Plus | 18.6 g/L | 160 mL/hL | Pyrethrins | 2 | |
| Potassium salts | Ciopper | 455 g/L | 150 mL/hL | Organic salts | 2 | |
| Mineral oil | Chemol Plus | 80% | 500 mL/hL | Mineral oils | 2 | |
| Kaolin | Surround | 95% | 4 Kg/hL | Kaolin | 2 | |
| 2019 | Untreated | - | - | - | - | - |
| Acetamiprid | Epik SL | 50 g/L | 150 mL/hL | Neonicotinoids | 1 | |
| Flupyradifurone | Sivanto Prime | 200 g/L | 60 mL/hL | Butenolides | 1 | |
| Pyrethrins | Biopiren Plus | 18.6 g/L | 160 mL/hL | Pyrethrins | 2 | |
| Pyrethrins + | Biopiren Plus + | 18.6 g/L | 160 mL/hL | Pyrethrins + | 2 | |
| mineral oil | Oliocin | 80% | 500 mL/hL | Mineral oils |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tirello, P.; Marchesini, E.; Gherardo, P.; Raniero, D.; Rossetto, F.; Pozzebon, A.; Duso, C. The Control of the American Leafhopper Erasmoneura vulnerata (Fitch) in European Vineyards: Impact of Synthetic and Natural Insecticides. Insects 2021, 12, 85. https://doi.org/10.3390/insects12020085
Tirello P, Marchesini E, Gherardo P, Raniero D, Rossetto F, Pozzebon A, Duso C. The Control of the American Leafhopper Erasmoneura vulnerata (Fitch) in European Vineyards: Impact of Synthetic and Natural Insecticides. Insects. 2021; 12(2):85. https://doi.org/10.3390/insects12020085
Chicago/Turabian StyleTirello, Paola, Enrico Marchesini, Pamela Gherardo, Damiano Raniero, Filippo Rossetto, Alberto Pozzebon, and Carlo Duso. 2021. "The Control of the American Leafhopper Erasmoneura vulnerata (Fitch) in European Vineyards: Impact of Synthetic and Natural Insecticides" Insects 12, no. 2: 85. https://doi.org/10.3390/insects12020085
APA StyleTirello, P., Marchesini, E., Gherardo, P., Raniero, D., Rossetto, F., Pozzebon, A., & Duso, C. (2021). The Control of the American Leafhopper Erasmoneura vulnerata (Fitch) in European Vineyards: Impact of Synthetic and Natural Insecticides. Insects, 12(2), 85. https://doi.org/10.3390/insects12020085