Seasonal Captures of Trissolcus japonicus (Ashmead) (Hymenoptera: Scelionidae) and the Effects of Habitat Type and Tree Species on Detection Frequency


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Seasonal Phenology and Habitat Type
2.2. Host Plant Comparisons
2.3. Parasitoid Identification
2.4. Statistical Analysis
3. Results
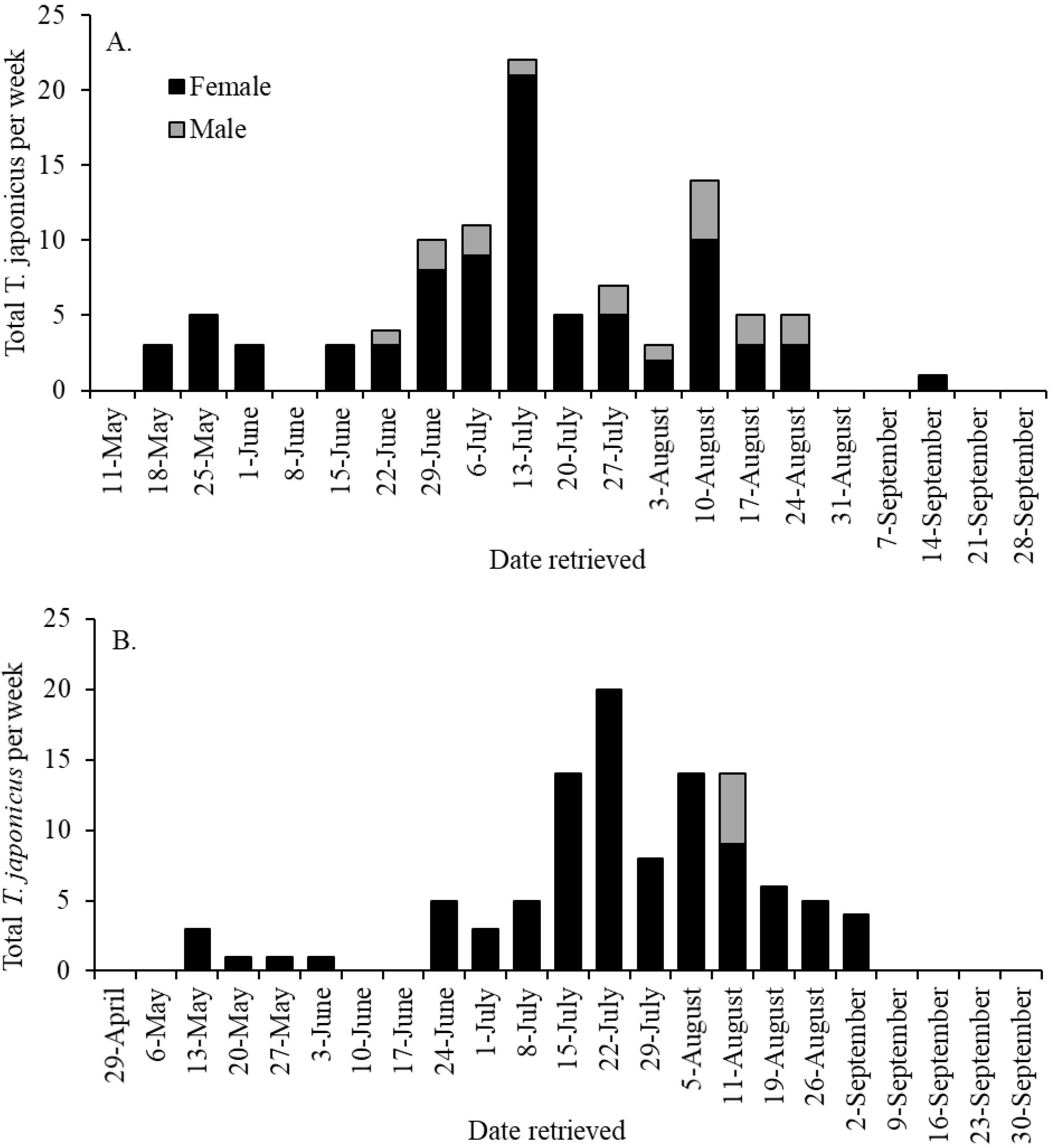
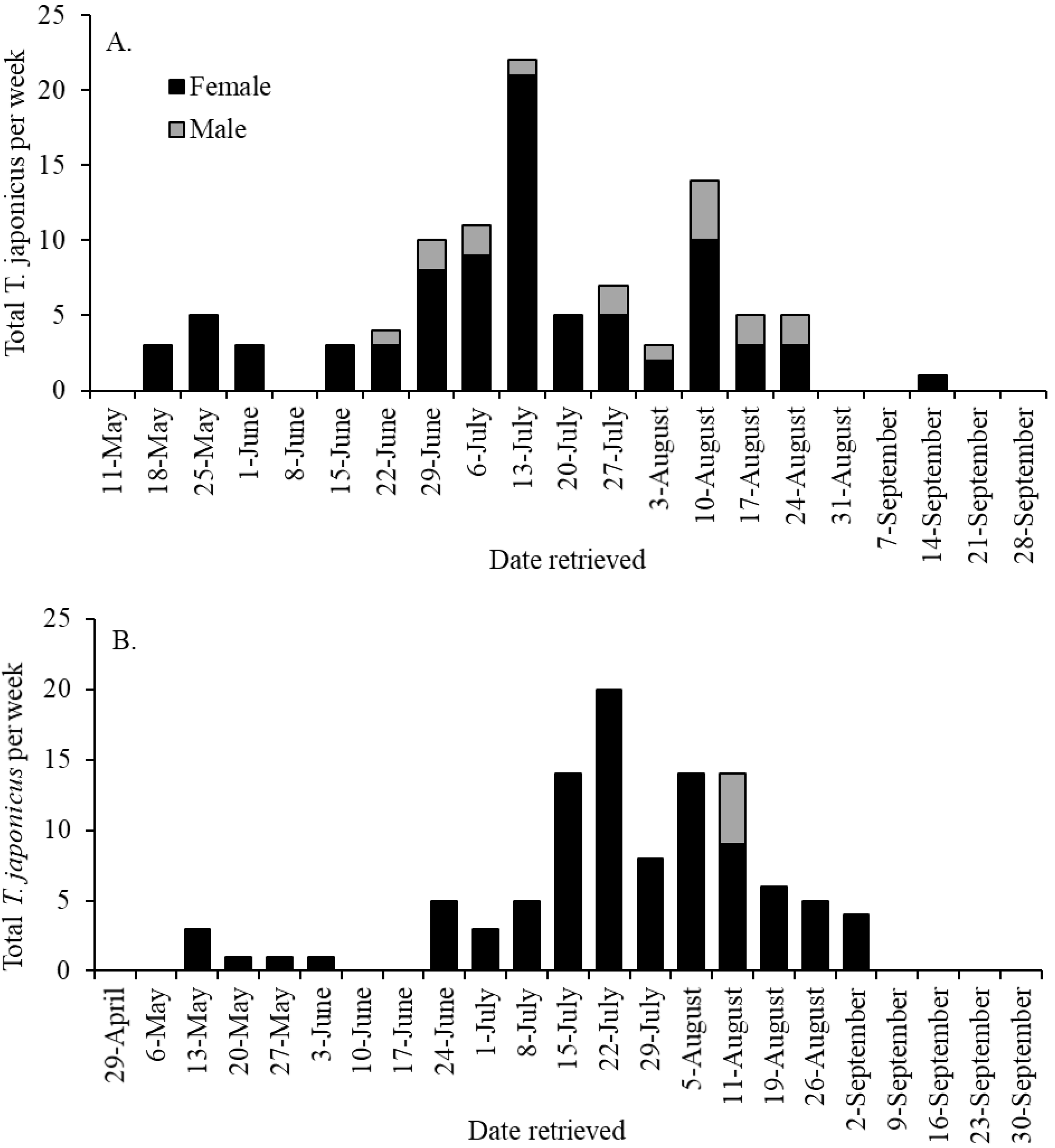
3.1. Seasonal Captures of Trissolcus japonicus
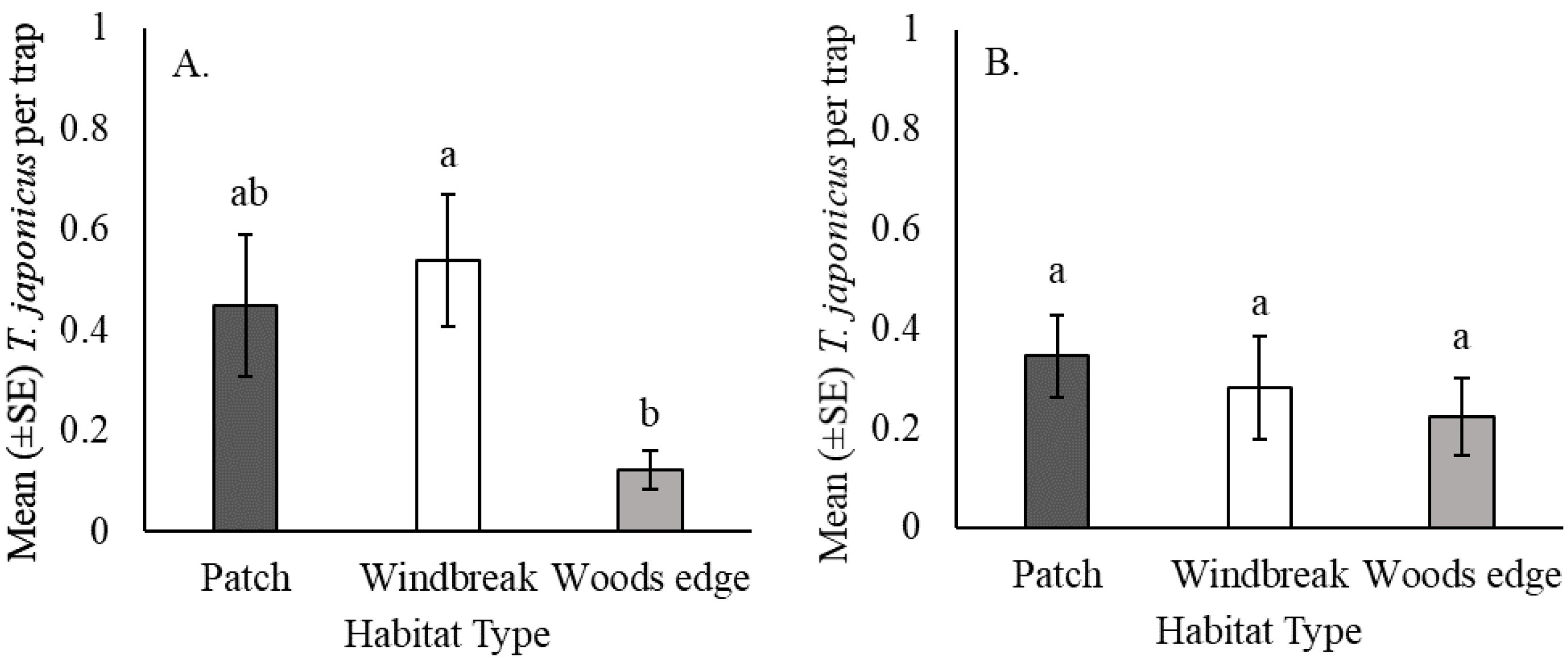
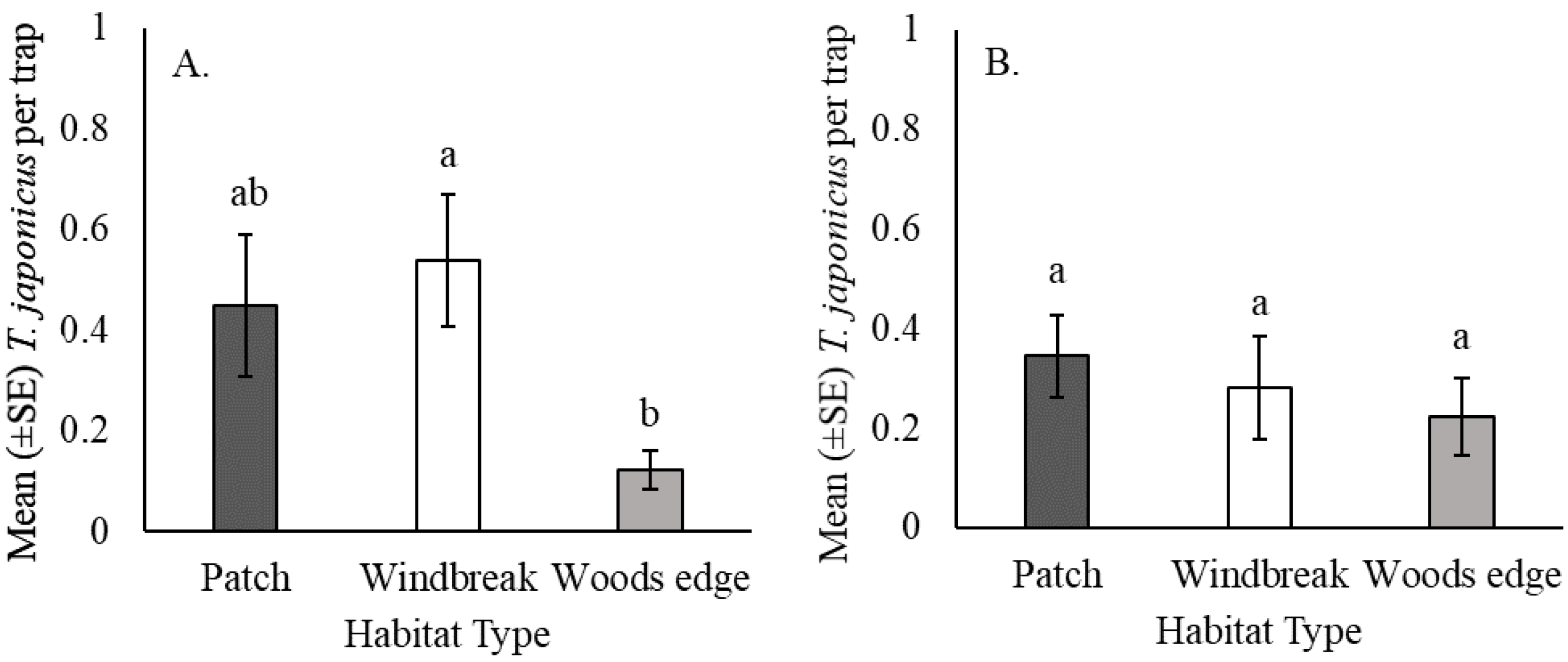
3.2. Effect of Habitat Type on T. japonicus Captures
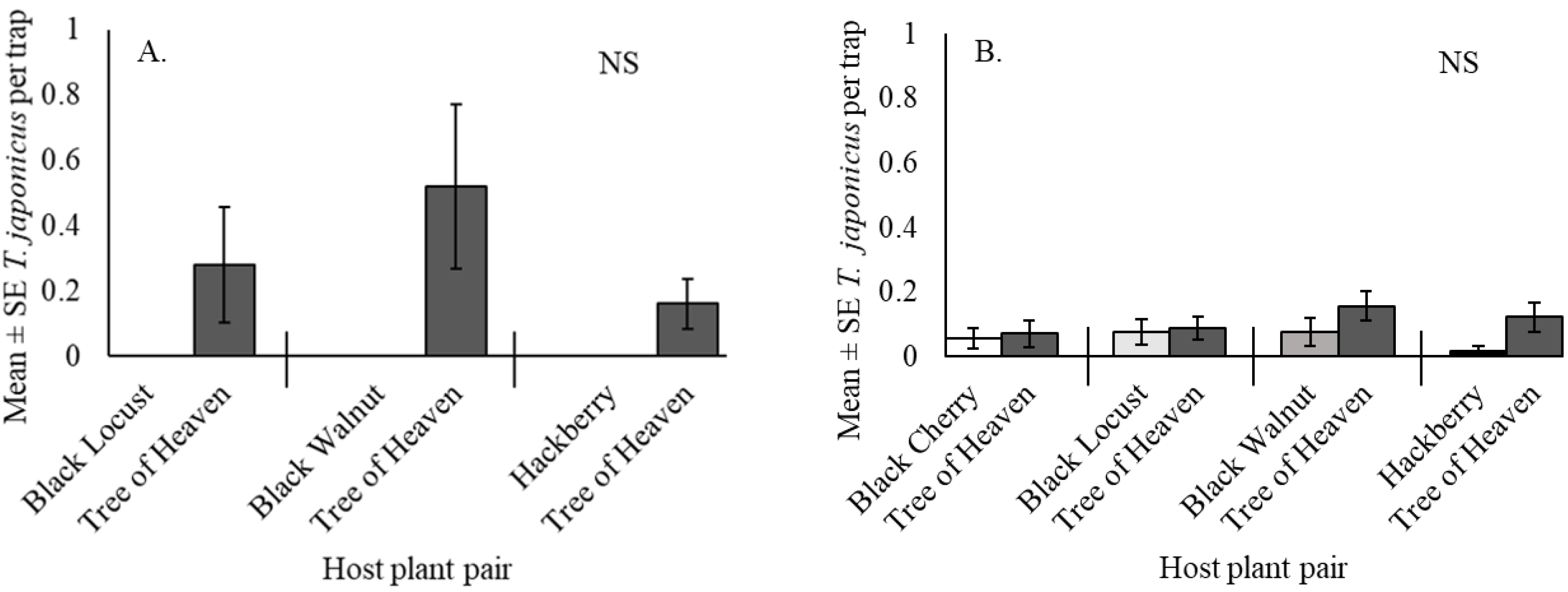
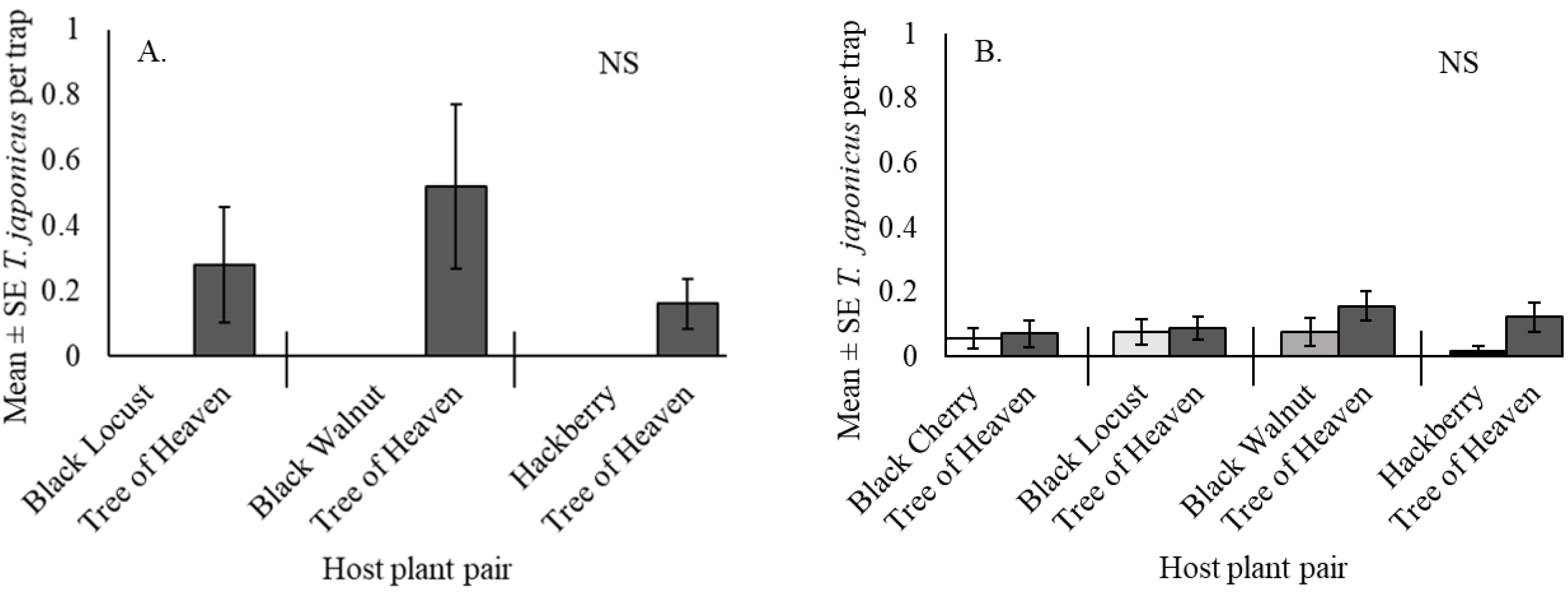
3.3. Trissolcus japonicus Captures in Paired Host Trees at the Woods Edge
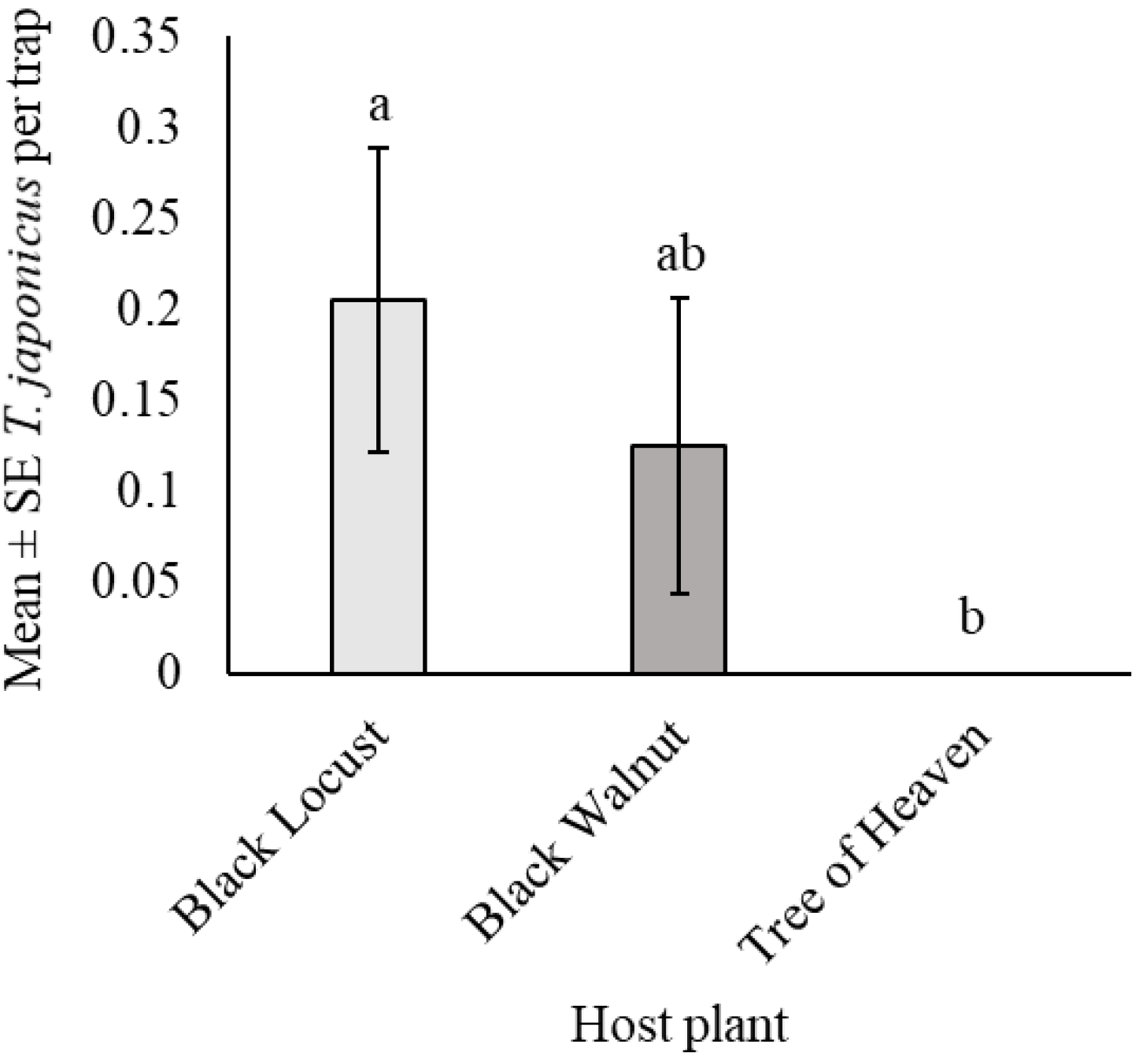
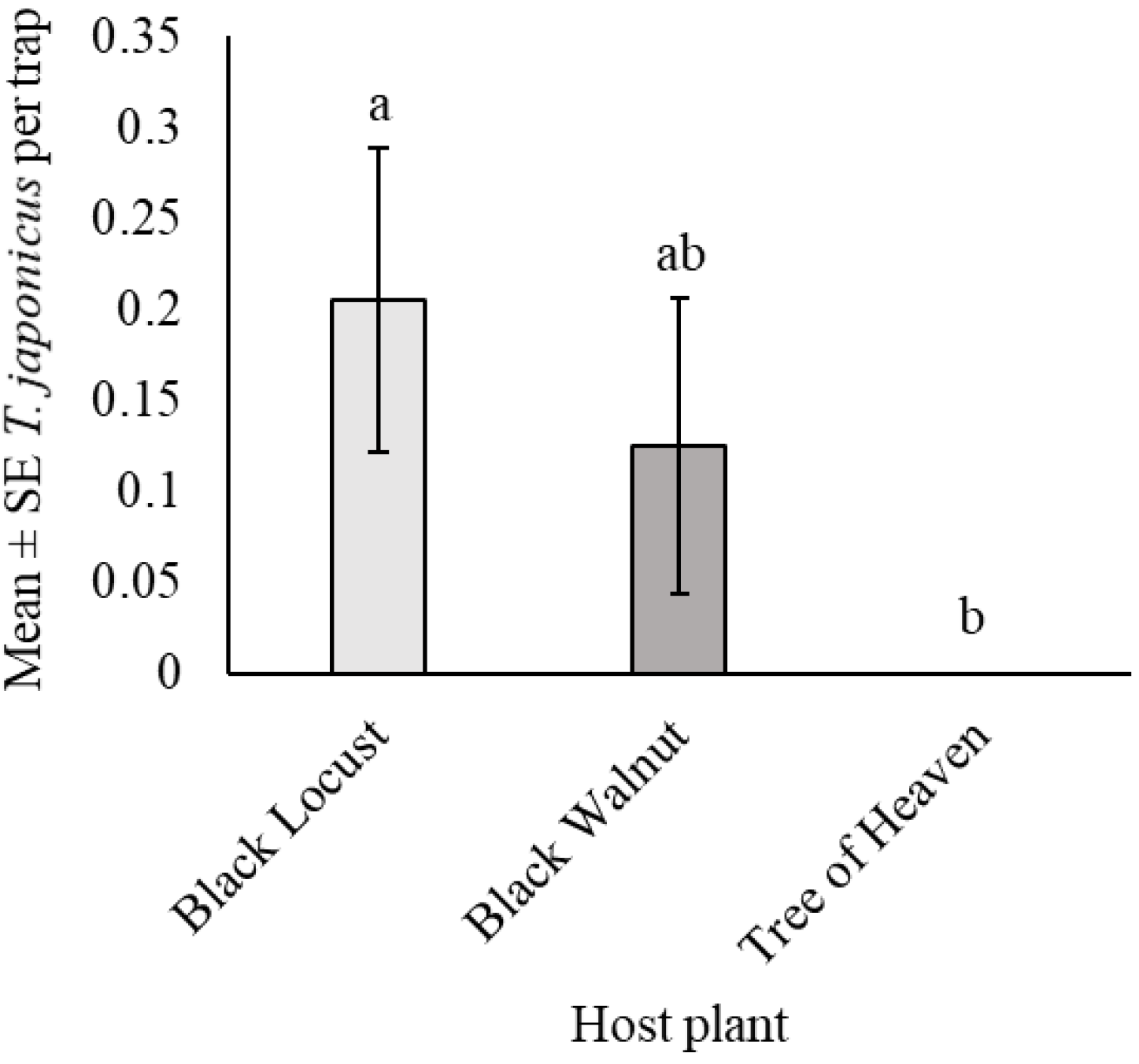
3.4. Trissolcus japonicus Captures Among Three Tree Species in Windbreaks
3.5. Captures of Native Parasitoids of Halyomorpha halys
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leskey, T.C.; Nielsen, A.L. Impact of the invasive brown marmorated stink bug in North America and Europe: History, biology, ecology, and management. Annu. Rev. Entomol. 2018, 63, 599–618. [Google Scholar] [CrossRef] [Green Version]
- Herrick, C. Brown Marmorated Stink Bug Causes $37 Million in Losses to Mid-Atlantic Apple Growers. Growing Produce. 2011. Available online: https://www.growingproduce.com/fruits/apples-pears/brown-marmorated-stink-bug-causes-37-million-in-losses-to-mid-atlantic-apple-growers/ (accessed on 15 May 2018).
- Leskey, T.C.; Lee, D.-H.; Short, B.D.; Wright, S.E. Impact of insecticides on the invasive Halyomorpha halys (Hemiptera: Pentatomidae): Analysis of insecticide lethality. J. Econ. Entomol. 2012, 105, 1726–1735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abram, P.K.; Hoelmer, K.A.; Acebes-Doria, A.; Andrews, H.; Beers, E.H.; Bergh, J.C.; Bessin, R.; Biddinger, D.; Botch, P.; Buffington, M.L.; et al. Indigenous arthropod natural enemies of the invasive brown marmorated stink bug in North America and Europe. J. Pest Sci. 2017, 90, 1009–1020. [Google Scholar] [CrossRef]
- Dieckhoff, C.; Tatman, K.M.; Hoelmer, K.A. Natural biological control of Halyomorpha halys by native egg parasitoids: A multi-year survey in northern Delaware. J. Pest Sci. 2017, 90, 1–16. [Google Scholar] [CrossRef]
- Herlihy, M.V.; Talamas, E.J.; Weber, D.C. Attack and success of native and exotic parasitoids on eggs of Halyomorpha halys in three Maryland habitats. PLoS ONE 2016, 11, e0150275. [Google Scholar]
- Cornelius, M.L.; Dieckhoff, C.; Hoelmer, K.A.; Olsen, R.T.; Weber, D.C.; Herlihy, M.V.; Talamas, E.J.; Vinyard, B.T.; Greenstone, M.H. Biological control of sentinel egg masses of the exotic invasive stink bug Halyomorpha halys (Stål) in Mid-Atlantic USA ornamental landscapes. Biol. Control 2016, 103, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.L.; Jennings, D.E.; Hooks, C.R.R.; Shrewsbury, P.M. Field surveys of egg mortality and indigenous egg parasitoids of the brown marmorated stink bug, Halyomorpha halys, in ornamental nurseries in the mid-Atlantic region of the USA. J. Pest Sci. 2017, 90, 1159–1168. [Google Scholar] [CrossRef]
- Yang, Z.Z.-Q.; Yao, Y.Y.-X.; Qiu, L.-F.L.; Li, Z.Z.-X. A new species of Trissolcus (Hymenoptera: Scelionidae) parasitizing eggs of Halyomorpha halys (Heteroptera: Pentatomidae) in China with comments on its biology. Ann. Entomol. Soc. Am. 2009, 102, 39–47. [Google Scholar]
- Zhang, J.; Zhang, F.; Gariepy, T.; Mason, P.; Gillespie, D.; Talamas, E.; Haye, T. Seasonal parasitism and host specificity of Trissolcus japonicus in northern China. J. Pest Sci. 2017, 90, 1127–1141. [Google Scholar] [CrossRef] [Green Version]
- Talamas, E.J.; Herlihy, M.V.; Dieckhoff, C.; Hoelmer, K.A.; Buffington, M.; Bon, M.-C.; Weber, D.C. Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae) emerges in North America. J. Hymenopt. Res. 2015, 43, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Hedstrom, C.; Lowenstein, D.; Andrews, H.; Bai, B.; Wiman, N. Pentatomid host suitability and the discovery of introduced populations of Trissolcus japonicus in Oregon. J. Pest Sci. 2017, 90, 1169–1179. [Google Scholar] [CrossRef]
- Milnes, J.M.; Wiman, N.G.; Talamas, E.J.; Brunner, J.F.; Hoelmer, K.A.; Buffington, M.L.; Beers, E.H. Discovery of an exotic egg parasitoid of the brown marmorated stink bug, Halyomorpha halys (Stål) in the Pacific Northwest. Proc. Entomol. Soc. Wash. 2016, 118, 466–470. [Google Scholar] [CrossRef]
- Morrison, W.R.; Blaauw, B.R.; Nielsen, A.L.; Talamas, E.; Leskey, T.C. Predation and parasitism by native and exotic natural enemies of Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) eggs augmented with semiochemicals and differing host stimuli. Biol. Control 2018, 121, 140–150. [Google Scholar] [CrossRef]
- Costi, E.; Haye, T.; Maistrello, L. Surveying native egg parasitoids and predators of the invasive Halyomorpha halys in Northern Italy. J. Appl. Entomol. 2019, 143, 299–307. [Google Scholar] [CrossRef]
- Peverieri, G.S.; Talamas, E.; Bon, M.C.; Marianelli, L.; Bernardinelli, I.; Malossini, G.; Benvenuto, L.; Roversi, P.F.; Hoelmer, K. Two Asian egg parasitoids of Halyomorpha halys (Stål) (Hemiptera, Pentatomidae) emerge in northern Italy: Trissolcus mitsukurii (Ashmead) and Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae). J. Hymenopt. Res. 2018, 67, 37–53. [Google Scholar] [CrossRef]
- Stahl, J.; Tortorici, F.; Pontini, M.; Bon, M.C.; Hoelmer, K.; Marazzi, C.; Tavella, L.; Haye, T. First discovery of adventive populations of Trissolcus japonicus in Europe. J. Pest Sci. 2019, 92, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Abram, P.K.; Talamas, E.J.; Acheampong, S.; Mason, P.G.; Gariepy, T.D. First detection of the samurai wasp, Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae), in Canada. J. Hymenopt. Res. 2019, 68, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Abram, P.K.; Mills, N.J.; Beers, E.H. Review: Classical biological control of invasive stink bugs with egg parasitoids—what does success look like? Pest Manag. Sci. 2020, 76, 1980–1992. [Google Scholar] [CrossRef]
- Kaser, J.M.; Akotsen-Mensah, C.; Talamas, E.J.; Nielsen, A.L. First report of Trissolcus japonicus parasitizing Halyomorpha halys in North American agriculture. Fla. Entomol. 2018, 101, 680–683. [Google Scholar]
- Quinn, N.F.; Talamas, E.J.; Acebes-Doria, A.L.; Leskey, T.C.; Bergh, J.C. Vertical sampling in tree canopies for Halyomorpha halys (Hemiptera: Pentatomidae) life stages and its egg parasitoid, Trissolcus japonicus (Hymenoptera: Scelionidae). Environ. Entomol. 2019, 48, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Quinn, N.F.; Talamas, E.J.; Leskey, T.C.; Bergh, J.C. Sampling methods for adventive Trissolcus japonicus (Hymenoptera: Scelionidae) in a wild tree host of Halyomorpha halys (Hemiptera: Pentatomidae). J. Econ. Entomol. 2019, 112, 1997–2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milnes, J.M.; Beers, E.H. Trissolcus japonicus (Hymenoptera: Scelionidae) causes low levels of parasitism in three North American pentatomids under field conditions. J. Insect Sci. 2019, 19, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowarik, I.; Säumel, I. Biological flora of central Europe: Ailanthus altissima (Mill.) Swingle. Perspectives in Plant Ecology. Perspect. Plant Ecol. Evol. Syst. 2007, 8, 207–237. [Google Scholar] [CrossRef]
- Call, L.; Nilsen, E. Analysis of spatial patterns and spatial association between the invasive tree-of heaven (Ailanthus altissima) and native black locust (Robinia pseudoacacia). Am. Midl. Nat. 2003, 150, 1–14. [Google Scholar] [CrossRef]
- Acebes-Doria, A.L.; Leskey, T.C.; Bergh, J.C. Temporal and directional patterns of nymphal Halyomorpha halys (Hemiptera: Pentatomidae) movement on the trunk of selected wild and fruit tree hosts in the Mid-Atlantic region. Environ. Entomol. 2017, 46, 258–267. [Google Scholar]
- Bakken, A.J.; Schoof, S.C.; Bickerton, M.; Kamminga, K.L.; Jenrette, J.C.; Malone, S.; Abney, M.A.; Herbert, D.A.; Reisig, D.; Kuhar, T.P.; et al. Occurrence of brown marmorated stink bug (Hemiptera: Pentatomidae) on wild hosts in nonmanaged woodlands and soybean fields in North Carolina and Virginia. Environ. Entomol. 2015, 44, 1011–1021. [Google Scholar]
- Talamas, E.J.; Johnson, N.F.; Buffington, M. Key to Nearctic species of Trissolcus Ashmead (Hymenoptera, Scelionidae), natural enemies of native and invasive stink bugs (Hemiptera, Pentatomidae). J. Hymenopt. Res. 2015, 43, 45–110. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.Y.; Zhong, Y.Z.; Zhang, J.P.; Wang, X.P.; Zhang, F. A comparative scanning electron microscopy study on antennal sensilla of Trissolcus japonicus and Trissolcus plautiae, egg parasitoids of stink bugs (Pentatomidae). Ann. Entomol. Soc. Am. 2016, 109, 112–120. [Google Scholar]
- SAS Institute Inc. SAS Software, version 9.4; The SAS Institute: Cary, NC, USA, 2018. [Google Scholar]
- Van Nouhuys, S.; Lei, G. Parasitoid-host metapopulation dynamics: The causes and consequences of phenological asynchrony. J. Anim. Ecol. 2004, 73, 526–535. [Google Scholar] [CrossRef]
- Bergh, J.C.; Morrison, W.R., III; Joseph, S.V.; Leskey, T.C. Characterizing spring emergence of adult Halyomorpha halys using experimental overwintering shelters and commercial pheromone traps. Entomol. Exp. Appl. 2017, 162, 336–345. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, A.L.; Chen, S.; Fleischer, S.J. Coupling developmental physiology, photoperiod, and temperature to model phenology and dynamics of an invasive heteropteran, Halyomorpha halys. Front. Physiol. 2016, 7, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godfray, H.C.J.; Hassel, M.P.; Holt, R.D. The population dynamic consequences of phenological asynchrony between parasitoids and their hosts. J. Anim. Ecol. 1994, 63, 1–10. [Google Scholar] [CrossRef]
- Nielsen, A.L.; Hamilton, G.C.; Matadha, D. Developmental rate estimation and life table analysis for Halyomorpha halys (Hemiptera: Pentatomidae). Environ. Entomol. 2008, 37, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Acebes-Doria, A.L.; Agnello, A.M.; Alston, D.G.; Andrews, H.; Beers, E.H.; Bergh, J.C.; Bessin, R.; Blaauw, B.R.; Buntin, G.D.; Burkness, E.C.; et al. Season-long monitoring of the brown marmorated stink bug (Hemiptera: Pentatomidae) throughout the United States using commercially available traps and lures. J. Econ. Entomol. 2020, 113, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Philips, C.R.; Kuhar, T.P.; Dively, G.P.; Hamilton, G.; Whalen, J.; Kamminga, K. Seasonal abundance and phenology of Halyomorpha halys (Hemiptera: Pentatomidae) on different pepper cultivars in the Mid-Atlantic (United States). J. Econ. Entomol. 2017, 110, 192–200. [Google Scholar]
- NEWA. 2019. Available online: http://newa.cornell.edu/ (accessed on 1 November 2019).
- Amir-Maafi, M.; Parker, B.L. Density dependence of Trissolcus spp. (Hymenoptera: Scelionidae) on eggs of Eurygaster integriceps Puton (Hemiptera: Scutelleridae). Arab J. Plant Prot. 2002, 20, 62–64. [Google Scholar]
- Elzinga, J.A.; van Nouhuys, S.; van Leeuwen, D.J.; Biere, A. Distribution and colonisation ability of three parasitoids and their herbivorous host in a fragmented landscape. Basic Appl. Ecol. 2007, 8, 75–88. [Google Scholar] [CrossRef]
- Valladares, G.; Salvo, A.; Cagnolo, L. Habitat fragmentation effects on trophic processes of insect-plant food webs. Conserv. Biol. 2006, 20, 212–217. [Google Scholar] [CrossRef]
- Meats, A.; Pando, M.S.C. Ratio-dependent parasitism with Trissolcus basalis (Wollaston) (Hymenoptera: Scelionidae) on egg rafts of Nezara viridula (Linnaeus) (Hemiptera: Pentatomidae): Effect of experimental variables and compatibility of “ratio” and “Holling” models. Aust. J. Entomol. 2002, 41, 243–252. [Google Scholar]
- Formella, A.; Dorman, S.J.; Taylor, S.V.; Kuhar, T.P. Effects of aggregation lure and tree species on Halyomorpha halys (Hemiptera: Pentatomidae) seasonal oviposition. J. Econ. Entomol. 2019, 113, 203–210. [Google Scholar] [CrossRef]
- Bergmann, E.J.; Venugopal, P.D.; Martinson, H.M.; Raupp, M.J.; Shrewsbury, P.M. Host plant use by the invasive Halyomorpha halys (Stål) on woody ornamental trees and shrubs. PLoS ONE 2016, 11, e1049975. [Google Scholar] [CrossRef] [PubMed]
- Boyle, S.M.; Weber, D.C.; Hough-Goldstein, J.; Hoelmer, K.A. Host kairomones influence searching behavior of Trissolcus japonicus (Hymenoptera: Scelionidae), a parasitoid of Halyomorpha halys (Heteroptera: Pentatomidae). Environ. Entomol. 2020, 49, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, G. Stochastic aggregative responses and spatial patterns of parasitism in patchy host-parasitoid interactions. Oecologia 1986, 70, 402–410. [Google Scholar] [PubMed]
- Tillman, P.G. Diversity of stink bug (Hemiptera: Pentatomidae) egg parasitoids in woodland and crop habitats in southwest Georgia, USA. Fla. Entomol. 2016, 99, 286–291. [Google Scholar] [CrossRef] [Green Version]
- Konopka, J.K.; Haye, T.; Gariepy, T.D.; McNeil, J.N. Possible coexistence of native and exotic parasitoids and their impact on control of Halyomorpha halys. J. Pest Sci. 2017, 90, 1119–1125. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat | Trissolcus | Telenomus | Anastatus | Encyrtidae | |||||
|---|---|---|---|---|---|---|---|---|---|
| japonicus | brochymenae | thyantae | euschisti | edessae | podisi | spp. | spp. | ||
| Patch | 40 | 2 | 0 | 4 | 0 | 2 | 2 | 2 | 0 |
| Windbreak | 50 | 6 | 0 | 14 | 1 | 3 | 2 | 1 | 0 |
| Woods edge | 11 | 2 | 1 | 11 | 0 | 4 | 5 | 4 | 2 |
| Total | 101 | 10 | 1 | 29 | 1 | 9 | 9 | 7 | 2 |
| Habitat | Trissolcus | Telenomus | Gryon | Anastatus | Encyrtidae | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| japonicus | brochymenae | thyantae | euschisti | podisi | persimilis | spp. | spp. | spp. | ||
| Patch | 43 | 2 | 5 | 18 | 20 | 0 | 9 | 1 | 5 | 2 |
| Windbreak | 35 | 4 | 7 | 25 | 17 | 1 | 2 | 1 | 5 | 0 |
| Woods edge | 26 | 4 | 7 | 15 | 6 | 2 | 10 | 0 | 4 | 1 |
| Total | 104 | 10 | 19 | 58 | 43 | 3 | 21 | 2 | 14 | 3 |
| Host Plant | Trissolcus | Telenomus | Gryon | ||
|---|---|---|---|---|---|
| japonicus | brochymenae | euschisti | spp. | spp. | |
| Black Locust | 0 | 1 | 1 | 3 | 1 |
| Tree of Heaven | 7 | 1 | 1 | 0 | 0 |
| Black Walnut | 0 | 0 | 0 | 0 | 0 |
| Tree of Heaven | 13 | 1 | 1 | 0 | 0 |
| Hackberry | 0 | 0 | 0 | 0 | 0 |
| Tree of Heaven | 4 | 0 | 2 | 0 | 0 |
| Total | 24 | 3 | 5 | 3 | 1 |
| Host Plant Pair | Trissolcus | Telenomus | Gryon | |||
|---|---|---|---|---|---|---|
| japonicus | thyantae | euschisti | podisi | spp. | spp. | |
| Black Cherry | 3 | 1 | 6 | 1 | 1 | 0 |
| Tree of Heaven | 4 | 0 | 1 | 0 | 0 | 0 |
| Black Locust | 5 | 0 | 4 | 1 | 1 | 0 |
| Tree of Heaven | 6 | 0 | 3 | 6 | 0 | 0 |
| Black Walnut | 5 | 0 | 3 | 0 | 0 | 0 |
| Tree of Heaven | 10 | 0 | 5 | 0 | 0 | 0 |
| Hackberry | 1 | 1 | 5 | 2 | 0 | 1 |
| Tree of Heaven | 8 | 0 | 8 | 1 | 1 | 0 |
| Total | 42 | 2 | 35 | 11 | 3 | 1 |
| Host Plant | Trissolcus | Telenomus | |||||
|---|---|---|---|---|---|---|---|
| japonicus | brochymenae | thyantae | euschisti | edessae | podisi | spp. | |
| Tree of Heaven | 0 | 1 | 2 | 6 | 1 | 9 | 2 |
| Black Locust | 8 | 1 | 0 | 9 | 0 | 2 | 0 |
| Black Walnut | 5 | 1 | 0 | 6 | 0 | 6 | 2 |
| Total | 13 | 3 | 2 | 21 | 1 | 17 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quinn, N.F.; Talamas, E.J.; Leskey, T.C.; Bergh, J.C. Seasonal Captures of Trissolcus japonicus (Ashmead) (Hymenoptera: Scelionidae) and the Effects of Habitat Type and Tree Species on Detection Frequency. Insects 2021, 12, 118. https://doi.org/10.3390/insects12020118
Quinn NF, Talamas EJ, Leskey TC, Bergh JC. Seasonal Captures of Trissolcus japonicus (Ashmead) (Hymenoptera: Scelionidae) and the Effects of Habitat Type and Tree Species on Detection Frequency. Insects. 2021; 12(2):118. https://doi.org/10.3390/insects12020118
Chicago/Turabian StyleQuinn, Nicole F., Elijah J. Talamas, Tracy C. Leskey, and J. Christopher Bergh. 2021. "Seasonal Captures of Trissolcus japonicus (Ashmead) (Hymenoptera: Scelionidae) and the Effects of Habitat Type and Tree Species on Detection Frequency" Insects 12, no. 2: 118. https://doi.org/10.3390/insects12020118
APA StyleQuinn, N. F., Talamas, E. J., Leskey, T. C., & Bergh, J. C. (2021). Seasonal Captures of Trissolcus japonicus (Ashmead) (Hymenoptera: Scelionidae) and the Effects of Habitat Type and Tree Species on Detection Frequency. Insects, 12(2), 118. https://doi.org/10.3390/insects12020118